半滑舌鳎酪氨酸酶基因(TYR)和多巴色素异构酶基因(DCT)的克隆表达与分析*
2022-01-04吴垚磊李仰真赵玉柱陈松林
吴垚磊 李仰真 王 娜 马 腾 赵玉柱 陈松林
半滑舌鳎酪氨酸酶基因()和多巴色素异构酶基因()的克隆表达与分析*
吴垚磊1,2李仰真1,2王 娜1马 腾1,2赵玉柱1,2陈松林1①
(1. 中国水产科学研究院黄海水产研究所 农业农村部海洋渔业可持续发展重点实验室 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室 山东 青岛 266071; 2. 上海海洋大学 水产科学国家级实验教学示范中心 上海 201306)
作为我国鲆鳎鱼类中的一种主要养殖品种,半滑舌鳎()有时会发生无眼侧黑化(Melanism)、有眼侧白化(Albinism)的体色异常现象。本研究克隆了半滑舌鳎体色相关的酪氨酸酶基因()和多巴色素异构酶基因()的cDNA序列,并对这2个基因进行系统发育分析和时空表达分析。实验获得了基因编码区cDNA序列长度为1620 bp,编码539个氨基酸;基因编码区cDNA序列长度为1551 bp,编码516个氨基酸。结果显示,和在20日龄前的鱼苗体内表达量较高,尤其是在变态关键时期(15~20日龄)表达量最高,30日龄时的表达量锐减到很低水平;在其他时期的皮肤组织中,这2个基因在有眼侧正常皮肤和无眼侧黑化皮肤的表达量最高,在有眼侧白化皮肤和无眼侧正常皮肤中表达量极低;在其他时期的其他组织中,在眼睛中的表达量最高,其次是肝脏,在脾脏和肌肉中表达量极低。研究表明,和基因是半滑舌鳎无眼侧黑化发生和有眼侧体色维持的关键基因。本研究为查明半滑舌鳎体色异常机制提供了重要依据和参考。
半滑舌鳎;体色;酪氨酸酶基因();多巴色素异构酶基因()
半滑舌鳎()是我国特有的珍贵经济鱼类,年产量达10 000 t以上(农业农村部渔业局, 2019)。近年来,在人工育苗及养殖条件下,苗种经常出现体色异常现象,主要表现为有眼侧白化(即有眼侧原本的黑色素细胞无法正常合成黑色素,正常情况下有眼侧为黄或金黄色)和无眼侧黑化(即无眼侧黑色素细胞大量异常聚集,正常情况下无眼侧为白色) (李仰真等, 2014),对其经济效益造成严重影响。
鲆鳎类具有与众不同的体色分布,这与其独特的变态发育过程有关。鲆鳎类的体色发生过程分为2个阶段:第1阶段,从出膜到其变态发育之前,黑色素细胞呈对称性分布,主要位于身体的两侧;第2阶段,眼睛向一侧移动,体型呈侧扁型,到变态发育完成,由体色对称转变成黑色素在有眼侧集中沉着,而无眼侧几乎无黑色素沉着(Burton, 1988)。
研究表明,酪氨酸酶(Tyrosinase, TYR)基因与人类疾病以及动物的体色具有密切关系。在黑色素的合成中,TYR是其中的关键环节,其C端有2个金属离子结合位点(与催化活性相关),是一种双核铜结合蛋白;其N端具有负责将合成的序列转运至胞内内质网进一步加工的信号肽(徐伟等, 2017)。
多巴色素异构酶(dopachrome tautomerase, DCT)是黑色素合成中的限速酶,具有包括TYR、DCT、TYRP1在内的酪氨酸酶家族相似的蛋白质一级结构(王亚琪等, 2019)。DCT在黑色素细胞内发挥调节功能,使酪氨酸酶的催化作用处于较优状态。当其表达水平发生变化时,体内的黑色素含量也会发生变化(孙学亮等, 2018)。王亚琪等(2019)研究发现,基因还调节细胞凋亡,决定着细胞的存活状态。
目前,鲜见海水鱼体色发育不同时期、的表达差异分析研究。近年来,研究人员对半滑舌鳎无眼侧黑化的分子机制进行了研究,发现视黄酸受体(宋雪松, 2018)、阿黑皮素原(POMC) (朱学武, 2016)、黑色素富集激素(MCH)及其受体等(徐永江等, 2017; 史学营等, 2017)在半滑舌鳎无眼侧黑化过程中发挥重要作用,且在此过程中,半滑舌鳎鳞片的组织学形态及相关调控过程也发生显著变化。本研究通过克隆半滑舌鳎、基因,并通过qRT-PCR进行定量表达分析,探讨这2个基因表达量与半滑舌鳎体色变化的关系,旨在了解体色变化的分子基础,为鱼类体色相关研究提供资料。
1 材料与方法
1.1 实验材料
1.1.1 样品采集 半滑舌鳎取自河北省唐山市维卓水产养殖有限公司。对亲鱼暂养、诱导性成熟,并对其人工授精。待受精卵充分发育、仔鱼顺利出膜,分别取出膜1、5、10、15、20和30 d的样品若干。待其生长至3月龄时,取半滑舌鳎有眼侧黑色皮肤(正常)、有眼侧白化皮肤、无眼侧白色皮肤(正常)和无眼侧黑化皮肤样本;取6月龄和12月龄半滑舌鳎有眼侧黑色皮肤(正常)、有眼侧白化皮肤、无眼侧白色皮肤(正常)、无眼侧黑化皮肤、肝、脾、眼、肌肉样本。以上样品置于RNA保存液(RNAwait,北京索莱宝科技有限公司),过夜后放入–20℃冰箱中保存,备用。
1.1.2 实验试剂 RNA提取使用TRIZol reagent (TaKaRa),RNA反转录试剂盒为PrimeScript RT reagent kit with a gDNA eraser (TaKaRa)。PCR扩增试剂为2×PCR MasterMix、DNA Marker DL 2000,胶回收试剂盒为TIANgelTM胶回收试剂盒(以上均为天根生化),荧光定量表达使用SYBR Premix Ex™ (TaKaRa)。连接转化使用pMD18-T Vector、Trans1-T1感受态细胞(上海天根生物)、RNaseA-free ddH2O、100 mg/mL氨苄(Amp)、LB液体培养基、固体抗Amp LB培养基。
1.2 实验方法
1.2.1 总RNA的提取 采集上述不同发育阶段的稚鱼以及3月龄幼鱼的不同皮肤组织;取6、12月龄鱼的4种不同皮肤组织、眼、肝脏、脾脏、肌肉,使用TRIZol法提取组织总RNA,琼脂糖凝胶电泳检测RNA完整性,并通过NanoVue™ Plus分光光度计检测RNA浓度及260 nm/280 nm值,–80℃保存。
提取半滑舌鳎不同发育阶段的稚鱼样本,3月龄幼鱼的4种不同皮肤组织以及6月龄和12月龄个体的有眼侧正常皮肤、有眼侧白化皮肤、无眼侧正常皮肤、无眼侧黑化皮肤、眼、肝脏、脾脏和肌肉共8个组织的RNA,通过PrimeScript RT reagent kit with a gDNA eraser (TaKaRa)反转录试剂盒进行反转录得到相应的cDNA。
1.2.2、基因cDNA的克隆和序列测定 在NCBI的GenBank数据库中搜索半滑舌鳎和基因的预测序列。确定、基因的保守区,用Primer 5.0在保守区设计特异性引物(表1)。基因PCR扩增条件:95℃ 5 min,之后40个循环(94℃ 30 s,58℃ 30 s,72℃ 1.5 min),72℃ 7 min,最后4℃保存;基因PCR扩增条件:95℃ 5 min,之后40个循环(94℃ 30 s,55℃ 30 s,72℃ 1.5 min),72℃ 7 min,最后4℃保存。
扩增产物经1.2%琼脂糖凝胶电泳分离后,用TIANgelTM胶回收试剂盒(北京天根生化)对目的片段进行纯化回收,连接至pMD19-T1载体(TaKaRa, 日本)上,转化到T1感受态细胞中,挑取其中的阳性克隆,送至睿博兴科生物技术有限公司测序。
1.2.3 序列分析 经过BioEdit比对和NCBI在线BLAST程序,检测所得序列的准确性,并得到、基因cDNA序列,通过NCBI搜索、基因的同源序列,并使用Blastx程序检索蛋白相似性;运用DNAMAN 6.0软件推导编码的氨基酸序列及蛋白分子量,通过SMART在线数据库进行相关蛋白的信号肽及结构域预测,并将半滑舌鳎和GenBank中其他物种的TYR、DCT蛋白序列进行比对分析;通过MEGA-X软件,以邻接法Neighbor-Joining (NJ)构建系统进化树(Saitou, 1987)。
1.2.4 实时荧光定量PCR分析 以测得的半滑舌鳎、基因序列为模板,设计定量引物(表1),同时,将半滑舌鳎的β-Actin基因作为、基因的内参基因,合成其特异性引物(表1)。使用SYBR Premix Ex™(TaKaRa)试剂盒,以得到的cDNA为模板,浓度为100 ng/μL,在ABI 7500 SDS仪(美国应用生物公司)上进行qRT-PCR反应。采用两步法,反应程序:95℃预变性30 s,95℃ 5 s,58℃ 30 s,共40个循环;60℃ 30 s,72℃ 30 s,熔解曲线温度为55℃~95℃。每个样品设3个生物学重复,3个技术重复,使用2−ΔΔCt法计算、基因的相对表达量。用Origin 7.5软件作图,用SPSS 17.0软件进行单因素方差分析(one-way ANOVA)。通过检验进行显著性比较,<0.05表示差异显著。

表1 半滑舌鳎TYR基因克隆使用的PCR扩增引物
2 结果与分析
2.1 TYR、DCT基因cDNA和编码蛋白序列分析
利用RT-PCR技术成功获得基因完整编码序列。半滑舌鳎基因编码区序列长为1620 bp,共编码539个氨基酸,其编码的TYR蛋白预测分子量为61.008 kDa,分析见图1。
利用RT-PCR成功获得基因完整编码序列。基因的开放阅读框为1551 bp,编码516个氨基酸,DCT蛋白预测分子量为57.948 kDa (图2)。
2.2 同源性比较与系统发育树
通过NCBI的Blastp程序,搜索TYR同源蛋白氨基酸序列,选取了大刺鳅(: XP_026189265.1)、斑马鱼(: AMO00812.1)、青鳉(: BAA06155.1)、尼罗罗非鱼(: XP_003441635.1)、黄金鲈(: XP_028429419.1)、绿头鸭(: XP_005029640.2)、小鼠(: BAX25613.1)、人(: AAA61242.1)等16种有代表性的物种并确定相似性(表2),通过MEGA-X进行系统进化分析(图3)。结果发现,半滑舌鳎TYR与鲈形目、颌针鱼目等鱼类为同一分支,而爬行类、哺乳类等构成另外独立的分支。
NCBI网站搜索DCT同源蛋白氨基酸序列,选取斑马鱼(: AAJ64222.1)、剑尾鱼(: XP_032423860.1)、鲫鱼(: XP_026126897.1)、红鳍东方鲀(: XP_003961932.1)、褐牙鲆(: XP_019945763.1)、雉鸡(: XP_031466474.1)、欧亚野猪(: BAF48040.1)等19种有代表性的物种,确定相似性(表3),并通过MEGA-X进行系统进化分析(图4)。
结果表明,半滑舌鳎DCT与鲀形目、鳉形目、鲱形目、鲤形目等其他鱼类形成独立的分支,而两栖、爬行和哺乳类形成另外的分支(图4)。
2.3 TYR、DCT基因表达差异分析
2.3.1、基因在半滑舌鳎早期发育阶段以及皮肤组织中的转录水平 在稚鱼发育不同时期的qRT-PCR结果显示(图3),从出膜后1~20 d的仔鱼组织中,的表达总体上呈现递增趋势,尤其是在变态关键时期(15~20日龄)表达量最高,30日龄时表达量锐减到很低水平。3个时期基因的实时荧光定量结果显示(图5),在半滑舌鳎各皮肤组织中,有眼侧黑色(正常)皮肤基因的表达量随时间变化具有显著差异(<0.05),从3月龄、6月龄到12月龄,该基因表达量呈递增趋势,而在有眼侧白化、无眼侧白色(正常)和无眼侧黑化皮肤中随时间变化差异不显著(>0.05);同一时期,4种皮肤组织基因的表达均存在显著差异(<0.05),黑色皮肤中该基因的表达显著高于有眼侧白化、无眼侧白色(正常)皮肤,而黑色皮肤之间和白色皮肤之间基因的表达均无显著差异(>0.05) (图6)。

图1 半滑舌鳎TYR基因ORF序列及推导的氨基酸序列

图2 半滑舌鳎DCT基因ORF序列及推导的氨基酸序列

表2 半滑舌鳎TYR氨基酸序列与其他脊椎动物的同源性比较

图3 TYR氨基酸序列的系统进化树分析
通过qRT-PCR技术,检测基因在不同时期仔鱼及成鱼皮肤组织中的表达情况(图6)。结果显示,从出膜后1~20 d的仔鱼组织中,的表达总体上呈递增趋势,尤其在变态关键时期(15~20日龄)表达量最高,30日龄时表达量锐减到很低水平。基因在不同发育时期的仔鱼体内的qRT-PCR结果显示(图7),在半滑舌鳎各皮肤组织中,有眼侧黑色(正常)皮肤、无眼侧黑化皮肤和无眼侧白色(正常)皮肤基因的表达随时间变化显著递增(<0.05),而在有眼侧白化皮肤中随时间变化差异不显著(>0.05);同一时期,4种皮肤中基因表达均存在显著差异(<0.05),黑色皮肤中该基因的表达显著高于白色皮肤(<0.05),而黑色皮肤之间和白色皮肤之间的基因表达均无显著差异(>0.05) (图8)。

表3 半滑舌鳎DCT氨基酸序列与其他脊椎动物的同源性比较

图4 DCT氨基酸序列的系统进化树分析

图5 半滑舌鳎TYR早期生长发育阶段相对定量表达

图6 半滑舌鳎TYR皮肤组织相对定量表达

图7 半滑舌鳎DCT在仔鱼生长发育阶段相对定量表达

图8 半滑舌鳎DCT皮肤组织相对定量表达
2.3.2 不同组织中、基因的表达差异 半滑舌鳎的6月龄和12月龄,基因在不同组织中的表达情况如图9、图10所示。在6月龄成鱼中,表达量在眼睛中最高,表达最高为眼部组织;另外,有眼侧黑色皮肤和无眼侧黑化皮肤表达量也相对较高;同时,肝脏组织也具有较高的表达量,在脾脏中极低表达,在有眼侧白化皮肤、无眼侧正常(白色)皮肤和肌肉中几乎不表达。在12月龄半滑舌鳎中,在眼睛中表达最高,但显著低于6月龄,其有眼侧正常皮肤(黑色)、无眼侧黑化皮肤的表达较高,脾脏、肝脏中的表达极低,无眼侧正常皮肤(白色)和肌肉中仅微量表达或几乎不表达。
在6月龄半滑舌鳎中,表达在眼睛中最高,肝脏中也有较高的表达量;另外,有眼侧黑色皮肤和无眼侧黑化皮肤表达量相对较少,有眼侧白化和无眼侧正常皮肤、脾脏、肌肉几乎不表达。在12月龄半滑舌鳎中,在眼睛中表达最高,但稍低于6月龄的表达量,肝脏、有眼侧正常皮肤(黑色)、无眼侧黑化皮肤中有较高表达,在无眼侧正常皮肤(白色)、脾脏和肌肉中几乎不表达。
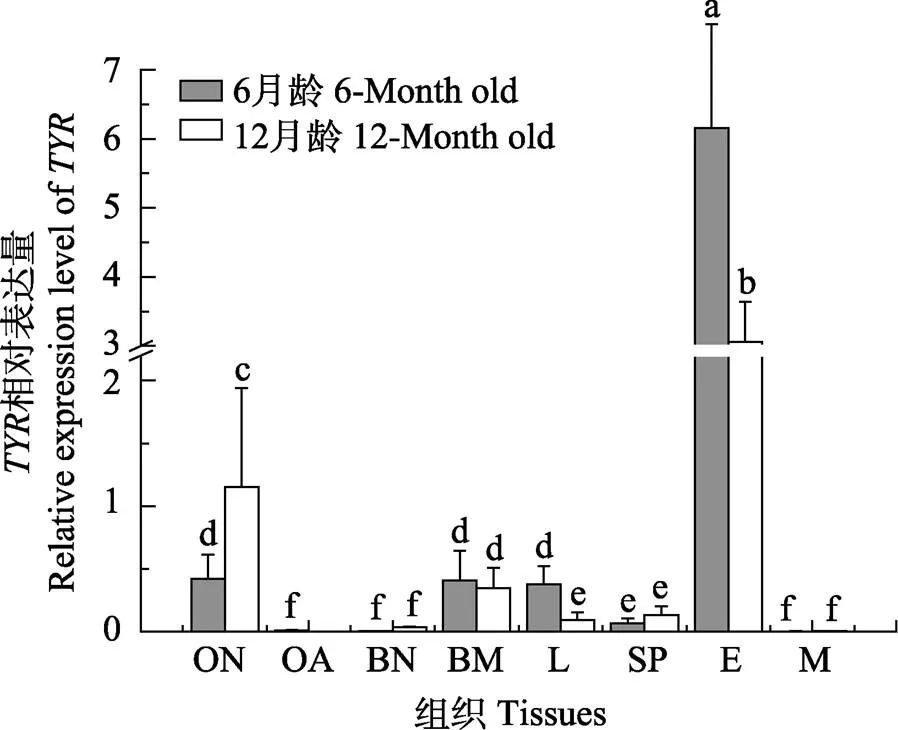
图9 半滑舌鳎TYR在不同组织中的相对表达量

图10 半滑舌鳎DCT在不同组织中的相对表达量
3 讨论
3.1 序列分析
本研究利用qRT-PCR技术,获得了半滑舌鳎的基因的编码区序列,其CDS序列由539个氨基酸构成,编码的TYR蛋白预测分子量为61.008 kDa,其蛋白N端具有由19个氨基酸组成的信号肽,具有指导蛋白质跨膜转移的作用;中间有由236个氨基酸组成的功能性结构域,与该蛋白执行黑色素合成的生理功能密切相关。氨基酸同源性分析显示,半滑舌鳎TYR与其他鱼类的同源性高达80%~85%,与爬行动物、鸟类和哺乳动物存在50%以上的同源性。
基因作为一种酪氨酸酶基因,其表达能够影响酪氨酸酶的活性以及黑色素合成的效率。本研究通过基因克隆技术获得了半滑舌鳎基因的编码区序列,该基因蛋白序列由516个氨基酸组成,DCT蛋白预测分子量为57.948 kDa,蛋白N端有由21个氨基酸组成的信号肽,具有指导蛋白质跨膜转移的作用;中间有由231个氨基酸组成的功能性结构域,与该蛋白执行黑色素合成与沉着的生理功能密切相关。氨基酸同源分析显示,半滑舌鳎DCT与其他硬骨鱼类有高达77.33%~88.05%的同源性,与爬行类、鸟类以及哺乳类存在50%以上的同源性。
3.2 时空表达差异分析
通过实时荧光定量PCR技术,检测了仔鱼、基因表达情况。结果显示,从出膜后1~20 d的仔鱼组织中,、的表达总体上呈递增趋势,表明、基因在半滑舌鳎变态发育过程中显著上调。
对半滑舌鳎不同生长时期的皮肤组织进行检测发现,基因在有眼侧白化皮肤和无眼侧白色皮肤(正常)中表达极低,且随时期变化无显著差异;该基因在有眼侧黑色(正常)皮肤和无眼侧黑化皮肤的表达量差异不大,但随着时期变化存在递增趋势,表达量为白色皮肤组织的20倍以上。Pomerantz等(1975)和Iwata等(1990)先后发现基因在人体黑色皮肤组织中活性显著高于非黑色皮肤组织,这与本研究的结果基本一致。
基因在同时期半滑舌鳎有眼侧白化和无眼侧白色(正常)皮肤中表达较低,无眼侧白色(正常)皮肤随时间变化存在递增趋势,有眼侧白化皮肤随时间变化无显著差异;该基因在有眼侧黑色(正常)皮肤和无眼侧黑化皮肤的表达差异不大,均随时间变化有递增趋势,表达量为白色皮肤的4倍以上。通过转录组技术对斑马鱼色素细胞分析显示,黑色素细胞中基因的表达显著高于其他色素细胞(Higdon, 2013);基因在鲫() (Zhu, 2016)和莫桑比克罗非鱼() (Zhang, 2017)黑色皮肤中的表达显著高于非黑色皮肤,与本研究的结果相一致。
对半滑舌鳎其他组织检测发现,基因在半滑舌鳎眼睛中的表达量极高,为肝脏表达量的16倍以上;其次为肝脏和脾脏,肌肉中的表达量极低,且均随时间变化无显著差异。基因在同一时期半滑舌鳎的眼睛中表达最高,约为肝脏表达量的10倍,且随时间变化有递减趋势;该基因在脾脏和肌肉中的表达量较低,尤其是肌肉中的表达可忽略不计。以上差异表明,、都与半滑舌鳎黑色素发生过程密切相关,但生理功能存在一定的差异。本研究还发现,该过程在眼睛中较黑色皮肤更为活跃。由此可以推测,黑色素对于鲆鳎鱼类眼色和视觉具有更重要的作用,这有待后续实验进行充分验证。
4 结论
本研究结合PCR技术和GenBank序列比对,获得半滑舌鳎酪氨酸酶相关基因和的编码区cDNA序列,分析了其分子结构、同源性及系统进化特性,通过定量方式研究了在组织中的表达特性。和基因在脊椎动物中具有进化保守性,均与爬行动物、鸟类和哺乳动物存在50%以上的同源性,与其他鱼类存在高达70%以上的同源性。通过实时荧光定量PCR实验阐明了半滑舌鳎出膜后不同时期和不同组织中的、基因表达特性,并分析了其与半滑舌鳎体色异常的关系,本研究为揭示半滑舌鳎体色异常机制提供了重要依据和参考。
BURTON D. Melanophore comparisons in different forms of ambicoloration in the flatfishand. Journal of Zoology, 1988, 214(2): 353–360
Fisheries and Fisheries Administration, Ministry of Agriculture and Rural Affairs. Chinese Fishery Statistics Yearbook. Beijing: Agricultural Press, 2019, 5–22 [农业农村部渔业渔政管理局. 中国渔业统计年鉴. 北京: 农业出版社, 2019, 5–22]
HIGDON C W, MITRA R D, JOHNSON S L. Gene expression analysis of zebrafish melanocytes, iridophores, and retinal pigmented epithelium reveals indicators of biological function and developmental origin. PLoS One, 2013, 8(7): e67801
IWATA M, CORN T, IWATA S,. The relationship between tyrosinase activity and skin color in human foreskins. Society for Investigative Dermatology, 1990, 95(1): 9–15
LI Y Z, CHEN S L, XING H F,. The preliminary study of albinism in. Journal of Fisheries of China, 2014, 38(1): 76–83 [李仰真, 陈松林, 邢贺飞, 等. 半滑舌鳎白化现象的初步研究. 水产学报, 2014, 38(1): 76–83]
POMERANTZ S H, ANCES I G. Tyrosinase activity in human skin influence of race and age in newborns. Journal of Clinical Investigation, 1975, 55(5): 1127–1131
SAITOU N, NEI M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 1987, 4(4): 406–425
SHI X Y, LIU X Z, SHI Y,. Molecular characterization ofand its corelation with blind-side hypermelanosis in. Progress in Fishery Sciences, 2017, 38(1): 91–102 [史学营, 柳学周, 石莹, 等. 半滑舌鳎()黑色素聚集素受体()表达特性及其与无眼侧黑化的关系. 渔业科学进展, 2017, 38(1): 91–102]
SONG X S. Cloning, expression patterns and physiological roles of two retinoic acid receptors in blind side hypermelanosis in. Master´s Thesis of Shanghai Ocean University, 2018 [宋雪松. 半滑舌鳎视黄酸受体克隆及其对无眼侧黑化的调控作用研究. 上海海洋大学硕士研究生学位论文, 2018]
SUN X L, GAO W W, SHI H Y,. Design and evaluation of primers for analysis ofgene expression in blood parrot ()by using real-time fluorescence quantitative PCR technique. Tianjin Agricultural Sciences, 2018, 24(12): 46–50 [孙学亮, 高微微, 石洪玥, 等. 血鹦鹉基因表达qPCR分析的引物设计与评估. 天津农业科学, 2018, 24(12): 46–50]
WANG Y Q, HU L L, WANG R N,. Advances in research on animal coat color candidate gene dopachrome- tautomerase (). Journal of Hebei Normal University of Science and Technology, 2019, 33(1): 54–57 [王亚琪, 胡露露, 王瑞宁, 等. 动物毛色候选基因的研究进展. 河北科技师范学院学报, 2019, 33(1): 54–57]
XU W, FENG J Q, HUANG L,. Research progress ongene. China Animal Husbandry Journal, 2017, 53(4): 23–26 [徐伟, 封竣淇, 黄兰, 等.基因研究进展. 中国畜牧杂志, 2017, 53(4): 23–26]
XU Y J, ZHU X W, LIU X Z,. Cloning and expression of melanin-concentrating hormone in half-smooth tongue sole (). Progress in Fishery Sciences, 2017, 38(1): 81–90 [徐永江, 朱学武, 柳学周, 等. 半滑舌鳎()黑色素富集激素基因的克隆和表达. 渔业科学进展, 2017, 38(1): 81–90]
ZHANG Y, LIU J, FU W,. Comparative transcriptome and DNA methylation analyses of the molecular mechanisms underlying skin color variations in Crucian carp (L.). BMC Genetics, 2017, 18(1): 95
ZHU W, WANG L, DONG Z,. Comparative transcriptome analysis identifies candidate genes related to skin color differentiation in red tilapia. Scientific Reports, 2016, 6: 31347
ZHU X W. Studies on regulation mechanisms underlie blind-side hypermelanosis of farmed flatfish. Master´s Thesis of Shanghai Ocean University, 2016 [朱学武. 养殖鲆鲽类无眼侧黑化调控机制研究. 上海海洋大学硕士研究生学位论文, 2016]
Expression Analysis ofandGenes Related to Body Color inat Different Periods and in Different Tissues
WU Yaolei1,2, LI Yangzhen1,2, WANG Na1, MA Teng1,2, ZHAO Yuzhu1,2, CHEN Songlin1①
(1. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences; Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs; Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao, Shandong 266071, China;2. National Experimental Teaching Demonstration Center of Aquatic Science, Shanghai Ocean University, Shanghai 201306, China)
Melanin commonly exists in animal tissues. The animal body surface has different colors and abundant pigment patterns due to differences in the distribution and development of melanin and related cells. Different pigment patterns have important functions such as biological camouflage, courtship, etc., especially in fish., one of the dominant aquaculture flatfish species in China, often suffers from two body color anomalies, i.e. melanism and albinism, which have hindered the development of high-quality seeds and aquaculture. In this study, cDNA sequences of two color-related genes (.and) were cloned and phylogenetically analyzed. Further, expression levels of both genes were analyzed in different stages and in different tissues. The cDNA sequence length ofgene coding region is 1620 bp, encoding 539 amino acids. The length of the cDNA sequence in the coding region ofgene is 1551 bp, encoding 516 amino acids. In this study, it was found that the expression levels ofandwere high in fry that were less than 20-day-old, especially during the critical period of metamorphosis (15 to 20-day-old), and decreased to a very low level at 30-day-old. In other skin tissues, the expression levels of these two genes were highest in normal skin on the ocular side and in dark skin on the blind side, and extremely low in albino skin on the ocular side and in normal skin on the blind side. Among other tissues at other times, the highest expression was in the eye, followed by the liver, with very low expression in the spleen and muscles. Results show thatandgenes are the key genes for blind-side melanogenesis and ocular-side color maintenance in. This study provides an important basis and reference for identifying the mechanism of color anomaly in.
; Body color; Tyrosinase gene (); Dopa isomerase gene ()
CHEN Songlin, E-mail: chensl@ysfri.ac.cn
S917.4
A
2095-9869(2021)06-0042-11
10.19663/j.issn2095-9869.20200516001
http://www.yykxjz.cn/
吴垚磊, 李仰真, 王娜, 马腾, 赵玉柱, 陈松林. 半滑舌鳎酪氨酸酶基因()和多巴色素异构酶基因()的克隆表达与分析. 渔业科学进展, 2021, 42(6): 42–52
WU Y L, LI Y Z, WANG N, MA T, ZHAO Y Z, CHEN S L. Expression analysis ofandgenes related to body color inat different periods and in different tissues. Progress in Fishery Sciences, 2021, 42(6): 42–52
陈松林,研究员,E-mail: chensl@ysfri.ac.cn
2020-05-16,
2020-05-29
*国家自然科学基金项目(31702333)、现代农业产业技术体系(CARS-47-G03)和山东省泰山学者攀登计划项目共同资助 [This work was supported by National Natural Science Foundation of China (31702333), China Agriculture Research System (CARS-47-G03), and Taishan Scholar Climbing Program of Shandong Province]. 吴垚磊,E-mail: 1305495981@qq.com
(编辑 冯小花)
