橘色双冠丽鱼黑色素细胞和黄色素细胞的分离、培养与鉴定*
2022-01-04宋红梅周康奇汪学杰牟希东胡隐昌
宋红梅 周康奇 田 雪 汪学杰 刘 超 刘 奕 牟希东 胡隐昌
橘色双冠丽鱼黑色素细胞和黄色素细胞的分离、培养与鉴定*
宋红梅1周康奇2田 雪3汪学杰1刘 超1刘 奕1牟希东1胡隐昌1①
(1. 中国水产科学研究院珠江水产研究所 农业农村部休闲渔业重点实验室 广东省现代休闲渔业工程技术研究中心 广东 广州 510380;2. 淡水鱼类资源与生殖发育教育部重点实验室 重庆 402460; 3. 河南师范大学水产学院 河南省水产动物养殖工程技术研究中心 河南 新乡 453007)
为探讨橘色双冠丽鱼()黑色素细胞和黄色素细胞的分离、培养与鉴定方法,观察鱼类色素细胞生长特征,本研究以橘色双冠丽鱼的尾鳍、鳞片、皮肤组织为材料,在25℃~28℃条件下,分别在胰蛋白酶溶液和胶原酶溶液中消化6h和12h,可有效分离出色素细胞团,细胞悬液经气孔尼龙网过滤后,于35%-45%-55% Percoll试剂中梯度分离和收集黄色素细胞和黑色素细胞。采用K-SFM培养基进行细胞原代培养和细胞纯化,抑制成纤维细胞和角朊细胞的生长,用十四烷酰佛波醇乙酸酯(TPA)、双抗、bFGF的DMEM培养基进行色素细胞传代培养;运用巴染色和透射电镜观察细胞形态,并用分子标记技术进行细胞鉴定。结果显示,细胞分离时,黑色素细胞位于Percoll试剂45%和35%浓度层之间,黄色素细胞位于Percoll试剂35%浓度层上,2种色素细胞均凝聚成絮状。透射电子显微镜观察发现,黑色素细胞内部含大量黑色素小体,而黄色素细胞内部含喋呤体和类胡萝卜素囊泡;黑色素细胞左旋多巴(L-DOPA)染色呈阳性;取第10代色素细胞进行体色相关基因黑皮素1受体(melanocortin receptor 1,)、酪氨酸酶(tyrosinase,)和()PCR检测,显示基因扩增条带特异性强,表明获得的2种色素细胞具有良好的生物学活性。本研究建立了橘色双冠丽鱼黄色素细胞和黑色素细胞的分离培养与鉴定方法,为进一步开展鱼类体色细胞分化和体色形成的分子机理研究提供了细胞模型。
橘色双冠丽鱼;色素细胞;分离;培养;鉴定
体色是鱼类独特的表型和经济性状(Sköld, 2013),尤其对于观赏鱼类,体色和斑纹直接决定其观赏价值和市场价值。鱼类体色繁杂多样、五彩斑斓,深入认识鱼类体色形成的调控机制,寻求人工改良鱼类体色的有效途径,是一个非常值得探索的课题。鱼类体色的形成和维持受到一系列细胞、基因和生理因素的复杂调控(Colihueque, 2010),其体色构成不仅与色素合成有关,更取决于皮肤、鳞片和鳍条所含色素细胞的种类、数量、分布及色素细胞之间的相互作用(Inaba, 2012)。色素细胞是鱼类体色形成的基本单位和重要载体,目前,已报道鱼类至少有黄色素细胞(xanthophores)、红色素细胞(erythrophores)、虹彩细胞(iridophores)、白色素细胞(leucophores)、黑色素细胞(melanophores)、蓝色素细胞(cyanophores)和紫红色素细胞(erythro-iridophores) 7种不同的色素细胞(徐伟等, 2007; Goda, 2011; Inaba, 2012; Singh, 2015)。
成功分离和体外培养色素细胞是探究动物体色相关基因生物学功能的基础,也是了解鱼类体色形成和体色变异分子机理的前提(Gola, 2012; Shigeki, 2011)。Eisinger等(1982)通过0.25%胰酶消化处理人包皮组织后,通过添加十四烷酰佛波醇乙酸酯(TPA)和霍乱毒素(CT)首次成功分离和体外培养了人的黑色素细胞;Tamura等(1987)通过人胚胎提取物和TPA,成功从小鼠皮肤获得黑色素细胞。此后,研究者也相继在其他动物皮肤中成功分离黑色素细胞,如羊驼()(白瑞等, 2013)、乌骨鸡()(田颖刚等, 2014)、狐() (鲍加荣, 2015)。但鲜见鱼类色素细胞分离培养的相关报道。本研究在参考之前哺乳动物等色素细胞提取研究的基础上,以具有典型体色褪色过程(黑色–灰色–亮黄色)的橘色双冠丽鱼()为实验对象,构建鱼类体表黑色素细胞和黄色素细胞分离、培养和鉴定方法,旨在为鱼类体色基因功能研究提供细胞模型,为利用体外培养色素细胞进行鱼类色素细胞分化和体色形成机理的相关研究奠定基础。
1 材料与方法
1.1 实验材料
实验用橘色双冠丽鱼采自中国水产科学研究院珠江水产研究所观赏渔业研究基地,挑选健硕无伤的鱼暂养,黑色时期平均体长为(5.0±0.5) cm,黄色时期平均体长为(7.0±0.5) cm,保证日常投喂和正常光照,控制适宜的养殖水温(23℃~28℃)和充足的溶氧。
1.2 主要试剂
K-SFM培养基、DMEM basic (1×)培养基、bFGF (碱性成纤维细胞生长因子)、大豆胰蛋白抑制剂均购自Gibco (美国);胰蛋白酶(Typsin 1∶250 for biochemistry)、EDTA(乙二胺四乙酸)、TPA、DNaseⅠ(脱氧核糖核酸酶Ⅰ)、99%胎牛血清(FBS)、胶原酶Ⅰ(CollageⅠ)、DMSO(二甲基亚砜)均购自Sigma公司(美国);左旋多巴(L-DOPA)均购自MCE(美国);牛血清蛋白、Percoll溶液购自Invitrogen公司(美国);新洁尔灭(利尔康,山东);PBS缓冲液、SSC杂交缓冲液(海利克思,上海);RNA提取试剂盒Total RNA kitⅡ购自OMEGA公司(美国);反转录试剂盒PrimeScriptTMⅡ1st Strand cDNA synthesis kit购自TaKaRa (日本);双抗原液(青霉素100 U/mL、链霉素100 μg/mL)购自中国医学科学院生物医学工程研究所;其他常规药品购自广州康龙生物科技有限公司。
1.2 实验方法
1.2.1 组织的消化处理与色素细胞分离 取所需鱼鳃放血致死,用0.1%新洁尔灭洗鱼体3次,再用75%酒精消毒鱼全身。常温下取所需鱼尾鳍、皮肤、鳞片组织,将采集的组织剪碎至1 mm3大小的块状,转入50 mL离心管中,加入胰蛋白酶溶液40~45 mL,混匀,25℃~28℃下消化6~8 h。弃胰蛋白酶溶液,用1×PBS在1000 r/min离心冲洗5 min,重复5次,去除表皮组织碎块。加入胶原酶溶液混匀,25℃~28℃下消化12~15 h。转入25 μm气孔尼龙网过滤分裂组织,放置5 min,800 r/min离心8 min,过滤得液体。吸取2 mL细胞溶液样品,置于Percoll分层试剂(浓度梯度为55%-45%-35%),1000 r/min离心20~25 min,使色素细胞分离于不同浓度层。
1.2.2 色素细胞的培养、纯化与传代 将分离的色素细胞悬液转移至KCl细胞培养基中,吹打均匀,取1滴溶液接种于5mL KCl培养基的25mL不透气培养瓶中,置于28℃培养箱中原代培养,并放入一杯水,保证培养箱内的湿度。
每3 h观察原代细胞生长情况,原代细胞长至约90%时,去培养液,胰蛋白酶消化,消化后去酶液,加入KC培养基纯化细胞,置于28℃培养箱培养,获得纯化的色素细胞。
纯化过的基础原代细胞铺满培养瓶底80%~90%时,加入MC2培养基,将细胞重悬,进行细胞传代。即在新25 mL不透气培养瓶中加入MC2培养基5mL,加入1滴纯化过的细胞,置于28℃培养箱中培养。对2~3代细胞进行形态学观察比较。
1.2.3 色素细胞的显微观察鉴定 参考常用黑色素细胞的DOPA染色法(Wang, 2013),分别取第3代生长旺盛的2种色素细胞各15mL,加入4%多聚甲醛固定10 min,用1×PBS洗涤3次,加入0.1% L-DOPA溶液,孵育4 h,2 h换液1次,制作细胞爬片,鉴定色素细胞的活性和生长情况。
再各取第3代生长旺盛的2种色素细胞15 mL,1000 r/min离心3 min,弃上清液,加入1.5 mL 4%戊二醛固定液固定,保存于4℃。在JEM-1400PLUS型透射电子显微镜(日本HITACHI)下观察拍照,分析色素细胞的形态结构。
1.2.4 色素细胞体色相关基因PCR鉴定 分别取第10代生长旺盛的2种色素细胞,用Total RNA KitⅡ(OMEGA)提取RNA,用PrimeScriptTMⅡ1st strand cDNA synthesis kit (TaKaRa)合成得到cDNA。
根据从NCBI的GenBank数据库中搜索与橘色双冠丽鱼序列及相关同源物种的基因,采用Primer Premier 6.0进行PCR引物设计(表1),引物由广州艾基生物技术有限公司合成。以2种色素细胞提取RNA反转录的cDNA为模板,利用3对引物分别进行PCR扩增,反应体系(50 μL):ddH2O 32.5μL,10×buffer 5 μL,dNTPs 8μL,模板cDNA (200 ng/μL) 2 μL,酶0.5 μL,正反向引物(10μmol/L)各1 μL。PCR参数:94℃预变性4 min;94℃变性30 s,退火60 s (退火温度见表1),72℃延伸30 s,共30个循环;72℃延伸7 min。电泳检测扩增结果。

表1 实验所用引物
2 结果
2.1 橘色双冠丽鱼色素细胞分离、培养和观察
2.1.1 色素细胞提取 以2个典型褪色时期“黑色期和黄色期”橘色双冠丽鱼为材料,提取获得的2种色素细胞均保持原有颜色(图1A)。通过Percoll试剂55%-45%-35%浓度梯度离心分层,黑色素细胞位于梯度层的45%~35%之间(图1B箭头b所指),而黄色素细胞位于35%浓度层上(图1B箭头a所指)。取2种色素置于同一Percoll浓度梯度试剂离心,2种色素细胞均能明显分离和分层,黑色素细胞在下层(图1C箭头b所指),位于45%~35%之间,黄色素细胞位于35%浓度层表面(图1C箭头a所指)。

图1 橘色双冠丽鱼色素细胞分离
2.1.2 分离培养后色素细胞形态学比较观察 初步分离的2种色素细胞均凝聚呈絮状,形态观察发现,絮状物各自分布了大量黑色素细胞和黄色素细胞(图2A、2B);低倍镜下观察培养的第1代色素细胞,发现黑色素细胞呈球状,有零星色素颗粒散落出细胞外(图2C),而黄色素色素细胞全部为单个色素个体,分布在培养基表面。黄色素细胞在培养前期,细胞小,单个不聚拢(图2D、2E);经过培养,细胞迅速生长,6~12 h覆盖培养基表面约70%后细胞个体变大,慢慢聚拢;过夜培养至覆盖培养基表面约90%时,大个体色素细胞聚拢(图2F),与在鳞片上直接观察到的色素细胞形状大小基本一致。
2.1.3 橘色双冠丽鱼体表色素细胞形态学观察 为与分离培养的色素细胞形态进行对比,分别从橘色双冠丽鱼“黑色–灰色–黄色”体色过渡期鱼体上取下背部鳞片进行色素细胞观察,发现黑色鱼体上的色素细胞占绝大比例,黑色素细胞单个成簇状,有分散和聚拢2种形态(图3A)。灰色鱼体上色素细胞数量少,个体小,有黑色和黄色色素同时存在,黑色素细胞颜色较黑色鱼体鳞片上的浅,为棕色,细胞个体也较小;出现的黄色素细胞为弥散状,分布于黑色素细胞下层,色素细胞不显现完整清晰的形状(图3B)。黄色鱼体上基本无黑色素细胞,有数量较多、个体较大的黄色素细胞,有的细胞颜色较深且偏红,呈凝聚状,类似于红色素细胞(图3C)。

图2 橘色双冠丽鱼色素细胞培养后形态学观察

图3 橘色双冠丽鱼鳞片色素细胞
2.2 2种色素细胞的显微观察与生物学鉴定
2.2.1 L-DOPA染色鉴定黑色素细胞酶的活性 取第3代生长旺盛的黑色素细胞进行L-DOPA染色,在显微镜下观察黑素细胞染色情况,可见斑点状黑色或灰色黑色素细胞,L-DOPA染色呈阳性(图4B)。黄色素细胞没有多巴染色,还未见黄色素细胞酶活性的方法,只能从显微镜下观察其生长情况。
2.2.2 透射电镜分析 通过透射电镜观察黑色素细胞,可见其内部含大量的黑色素小体(图5A)。电镜下,观察到黄色素细胞内部的喋呤体(图5箭头所指a)和类胡萝卜素囊泡(图5箭头所指b),细胞内部结构完整。
2.2.3 2种色素细胞和基因的PCR检测 第10代生长旺盛的黑色素细胞和黄色素细胞和的PCR检测结果显示,扩增条带特异性良好(图6),说明获得的黑色素细胞和黄色素细胞均保持着较好的生物活性。

图4 橘色双冠丽鱼黑色素细胞的L-DOPA染色
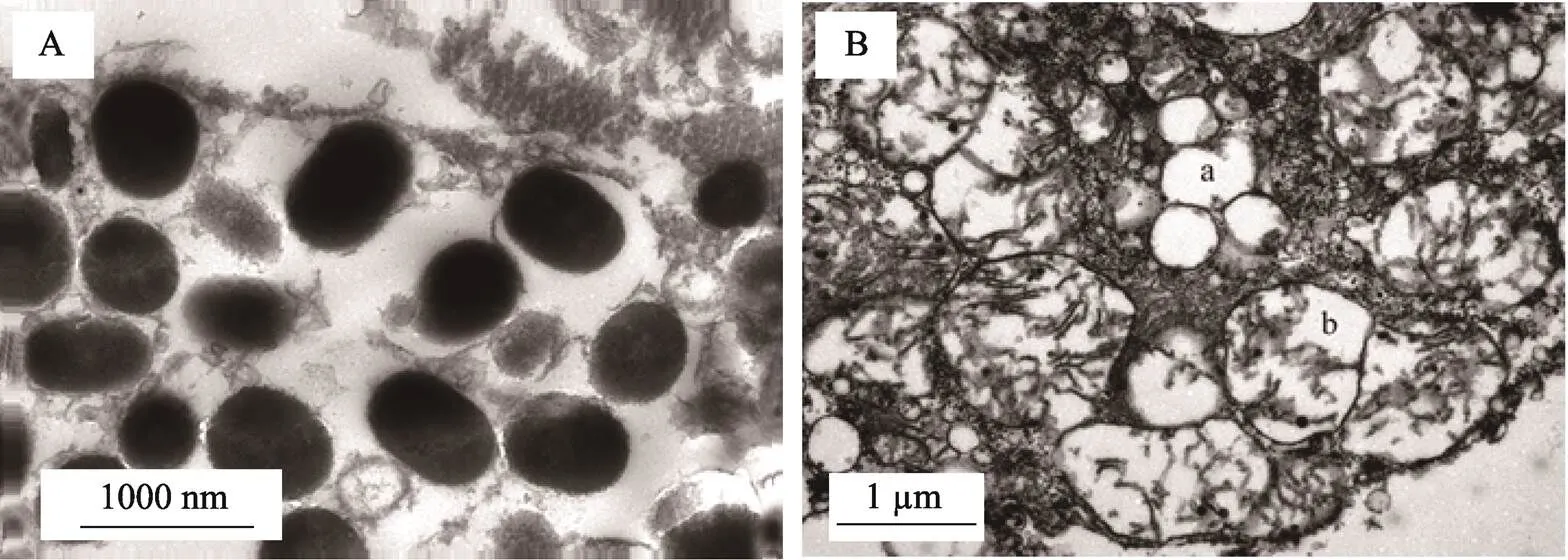
图5 电镜观察黑色素细胞和黄色素细胞的显微结构
3 分析与讨论
在细胞分离培养中,运用不同消化酶对组织进行消化处理是影响细胞提取效果的重要因素,在常用的几种组织消化酶中,DispaseⅡ酶对皮肤组织消化相对较弱,多应用于哺乳动物眼睛黑色素细胞的分离培养(鲍加荣, 2015);中性蛋白酶可选择性作用于纤维蛋白、黏连蛋白和Ⅳ型胶原,破环板桥内的结构,但对于桥粒结构作用却很少,很难将表皮完全分散;胰蛋白酶能水解细胞间质中的蛋白质,破环细胞间的半桥粒和桥粒结构,从而使细胞分散;胶原酶Ⅰ作用于结缔组织中的胶原蛋白,对细胞间质产生消化作用,且对细胞本身影响不大,能使细胞保持高活力(田颖刚等, 2014; 鲍加荣, 2015)。本研究参考Inaba等(2012)的方法并改进,在25℃~28℃下用2.5 mg/mL的胰蛋白酶消化组织6~8 h,去除表皮细胞,再用0.1 mg/mL胶原酶Ⅰ配制的溶液进行二次消化12~15 h,可分离获得活性较高的色素细胞原液。分离的2种色素细胞在Percoll梯度试剂中低速离心后分层明显,细胞层间隔35%的Percoll试剂层,推测黑色素细胞较黄色素细胞质量重,确定2种色素细胞分离时所在Percoll试剂浓度层,可为下一步实验得到培养纯化细胞奠定基础。
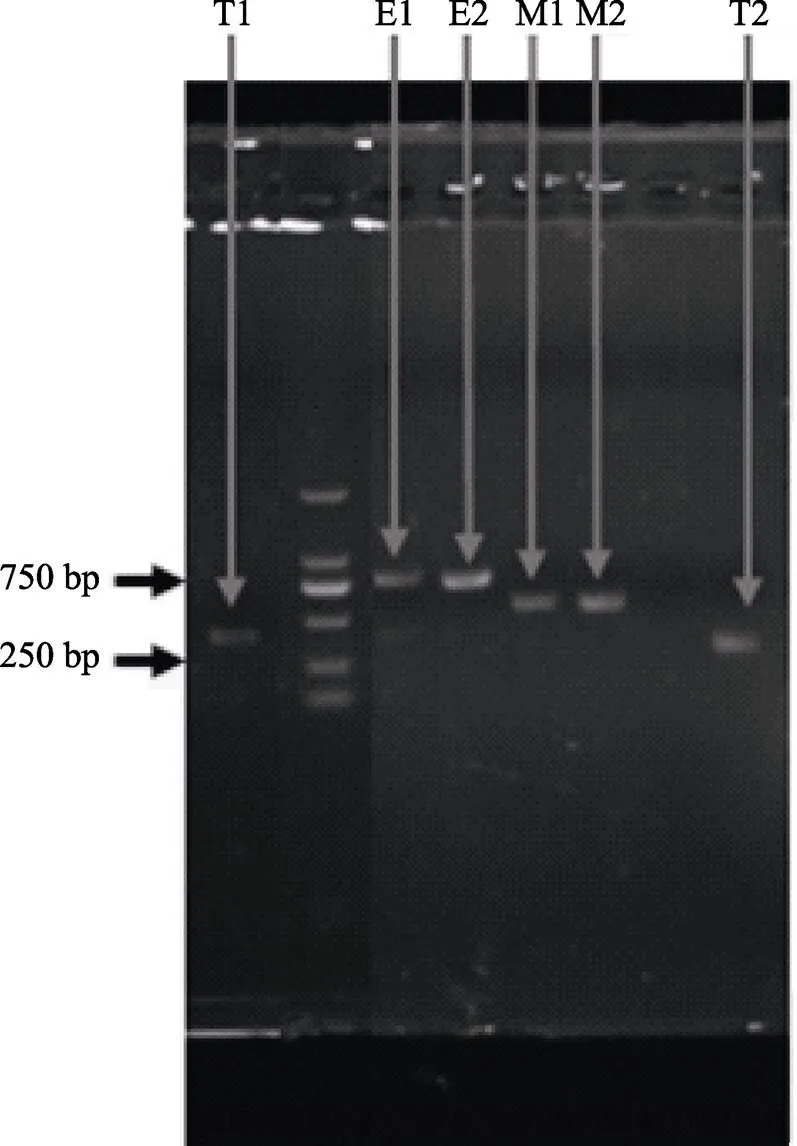
图6 色素细胞中TYR、EDNRB和MC1R基因的PCR片段扩增
色素细胞体外培养一般采用K-SFM角朊细胞培养基、DMEM、M199等,比较K-SFM和DMEM/F12 2种培养基对羊驼毛囊干细胞生长和增殖的影响,发现K-SFM培养基更有利于毛囊干细胞增殖,培养出的细胞数量多、增殖快,克隆生成能力更强(弓慧敏等, 2016)。田颖刚等(2014)以K-SFM培养基为原代培养和细胞纯化,以DMEM培养基为最终传代培养,成功建立乌骨鸡黑色素细胞系。本研究借鉴哺乳动物和鸟类(Deveci, 2001)的培养方法加以改进,采用能抑制成纤维细胞和角朊细胞生长的K-SFM培养基进行细胞原代培养和细胞纯化,以DMEM培养基为最终传代培养,获得生理状态良好的色素细胞。
鱼类的鳞片、皮肤和尾鳍有很多胶原纤维细胞和角质细胞,数量远远大于黑色素细胞和黄色素细胞,因此,色素细胞培养中,杂细胞的去除非常关键,培养基中添加促癌因子TPA可对不同细胞产生选择性毒性作用。TPA能促进色素细胞分泌特定的生长因子,促进细胞的生长、粘附和增殖,抑制成纤维细胞及角质形成,达到避免成纤维细胞和胶质污染,从而获得纯化的色素细胞(Yaar, 1991)。因此,本研究在原代培养和细胞纯化时,增大TPA用量(10 ng/mL),但传代培养时,应减少TPA用量(1 ng/mL),以减少对2种色素细胞的毒性作用。
本研究在鳞片上直接观察到的黑、黄2种色素细胞个体分明,成群地分布在鳞片上,而提取到的2种色素细胞在低倍镜下观察呈絮状,但也能看到一些单个的色素细胞,为判定所提取的色素细胞有无破裂、有无增殖能力及生物活性状况等,对色素细胞进行培养后显微观察,发现2种色素细胞均能正常生长,其中,黄色素细胞迅速生长,细胞个体变大后,慢慢聚拢成球状。通过多巴染色和透射电镜来观察黑色素小体是鉴定黑色素细胞的经典方法(Deveci, 2001; Fuller, 2001; Hu, 2007)。多巴染色培养的黑色素细胞呈现阳性反应,高倍透射电镜观察黑色素细胞内部结构,显现大量黑色素小体;黄色素细胞内部可见喋呤体和类胡萝卜素囊泡,2种色素细胞内部结构与在大麻哈鱼(Walbaum)皮肤表面通过电镜观察到的状态相似(Djurdjevič, 2015),佐证本研究所建立的提取、分离和体外培养橘色双冠丽鱼体表黑、黄2种色素细胞方法的可行性。
鱼类体色的分布和形成主要由表现为黑色、棕色表型的真黑色素和表现为黄色、红色表型的褐黑色素2种黑色素的相对数量及分布决定,二者均源于黑色素合成信号通路,都在酪氨酸基因()的调控下表达(Lin, 2007; 王成辉, 2012)。而基因和基因在黑色素合成通路中起着类似于“开关”的作用(Adalsteinsson, 1987; 蒋燕玲等, 2016)。在色素细胞发育中,基因能促进神经嵴细胞的增殖,调节神经嵴细胞分化为色素细胞(Parichy, 2000)。因此,本研究从黑、黄2种色素细胞中提取总RNA,检测色素细胞中体色关键基因和的表达,发现均能扩增出特异性条带,表明获得的黑色素细胞和黄色素细胞具有良好的生物学状态。
本研究利用Percoll梯度试剂分离橘色双冠丽鱼黑色素细胞和黄色素细胞,以K-SFM培养基(添加10 ng/mL TPA)进行原代和纯化培养,最后培养于DMEM培养基,并添加1 ng/mL TPA、20 ng/mL bFGF和1%双抗的完全培养基中,建立了2种色素细胞的培养体系,并利用形态观察、多巴染色、透射电镜观察和体色相关基因表达对培养细胞的特性进行研究,为进一步构建橘色双冠丽鱼黑色素细胞和黄色素细胞系奠定基础,为了解鱼类色素细胞发育、体色形成及相关基因功能等提供参考。
ADALSTEINSSON S, HERSTEINSSON P, GUNNARSSON E. Fox colors in relation to colors in mice and sheep. Journal of Heredity, 1987, 78(4): 235–237
BAI R, YU Z H, FAN R W,. Culture, identification of alpaca skin melanocytes in vitro. Chinese Journal of Animal and Veterinary Sciences, 2013, 44(4): 549–556 [白瑞, 于志慧, 范瑞文, 等. 羊驼皮肤黑素细胞体外培养鉴定. 畜牧兽医学报, 2013, 44(4): 549–556]
BAO J R. Analyzing coat color differences between red and silver fox (), pigment genes cloning and tentative functional investigation of TYRP1. Doctoral Dissertation of Chinese Academy of Agricultural Sciences, 2015 [鲍加荣. 赤狐和银黑狐毛色差异分析、色素基因克隆及TYRP1功能初步研究. 中国农业科学院博士研究生学位论文, 2015]
COLIHUEQUE N. Genetics of salmonid skin pigmentation: Clues and prospects for improving the external appearance of farmed salmonids. Reviews in Fish Biology and Fisheries, 2010, 20(1): 71–86
DEVECI M, GILMONT R R, TERASHI H,. Melanocyte- conditioned medium stimulates while melanocyte/keratinocyte contact inhibits keratinocyte proliferation. Journal of Burn Care and Rehabilitation, 2001, 22(1): 9–14
DJURDJEVIČ I, KREFT M E, SUŠNIK B S. Comparison of pigment cell ultrastructure and organisation in the dermis of marble trout and brown trout, and first description of erythrophore ultrastructure in salmonids., 2015, 227(5): 583–595
EISINGER M, MARKO O. Selective proliferation of normal human melanocytesin the presence of phorbol ester and cholera toxin., 1982, 79(6): 2018–2022
FULLER B B, SPAULDING D T, SMITH D R. Regulation of the catalytic activity of preexisting tyrosinase in black and Caucasian human melanocyte cell cultures., 2001, 262(2): 197–208
GODA M, OHATA M, IKOMA H,. Integumental reddish-violet coloration owing to novel dichromatic chromatophores in the teleost fish,. Pigment Cell and Melanoma Research, 2011, 24(4): 614–617
GOLA M, CZAJKOWSKI R, BAJEK A,. Melanocyte stem cells: Biology and current aspects. Medical Science Monitor: International Medical Journal of Experimental and Clinical Research, 2012, 18(10): A155–A159
GONG H M, BAI R, JIA Z F,. Effect of two kinds of culture medium on the alpaca hair follicle stem cell growth and proliferation. Journal of Shanxi Medical University, 2016, 47(5): 424–428 [弓慧敏, 白瑞, 贾宗菲, 等. 两种培养基对羊驼毛囊干细胞生长和增殖的影响. 山西医科大学学报, 2016, 47(5): 424–428]
HU D N, MCCORMICK S A, SEEDOR J A,. Isolation, purification and cultivation of conjunctival melanocytes., 2007, 84(4): 662
INABA M, YAMANAKA H, KONDO S. Pigment pattern formation by contact-dependent depolarization. Science, 2012, 335(6069): 677
JIANG Y L, SONG H M, LIU Y,. Cloning and expression analysis of the developing sequence and tissue expression of TYR gene in. Journal of Agricultural Biotechnology, 2016, 24(5): 697–707 [蒋燕玲, 宋红梅, 刘奕, 等. 橘色双冠丽鱼TYR基因的克隆及其发育时序和组织表达分析. 农业生物技术学报, 2016, 24(5): 697–707]
LIN J Y, FISHER D E. Melanocyte biology and skin pigmentation. Nature, 2007, 445(7130): 843–850
PARICHY D M, MELLGREN E M, RAWLS J F,. Mutational analysis of endothelin receptor b1 (rose) during neural crest and pigment pattern development in the zebrafish., 2000, 227(2): 294–306
SHIGEKI O, YOICHI I, YOHEI O,. Generation of human melanocytes from induced pluripotent stem cells. PLoS One, 2011, 6(1): e16182
SINGH A P, NÜSSLEIN-VOLHARD C. Zebrafish stripes as a model for vertebrate colour pattern formation., 2015, 25(2): R81–R92
SKÖLD H N, ASPENGREN S, WALLIN M. Rapid color change in fish and amphibians: Function, regulation, and emerging applications. Pigment Cell and Melanoma Research, 2013, 26(1): 29–38
TAMURA A, HALABAN R, MOELLMANN G,. Normal murine melanocytes in culture. In Vitro Cellular and Developmental Biology, 1987, 23(7): 519–522
TIAN Y G, LIAO C Y, XU D L. Primary culture and identification of skin melanocytes from black-bone silky fowl (Domesticus Brisson). China Poultry, 2014, 36(12): 6–10 [田颖刚, 廖春艳, 徐德利. 乌骨鸡皮肤黑色素细胞的原代培养及鉴定. 中国家禽, 2014, 36(12): 6–10]
WANG C H. Brief summary on genetic basis of pigmentation in fish. Journal of Shanghai Ocean University, 2012, 21(5): 103–108 [王成辉. 鱼类体色变异的遗传基础研究进展简述. 上海海洋大学学报, 2012, 21(5): 103–108]
WANG D, XU X, MA H,. Optimization of the method for the culture of melanocyte precursors from hair follicles and their activation by 1, 25-dihydroxyvitamin D3. Experimental and Therapeutic Medicine, 2013, 6(4): 967–972
XU W, LI C T, CAO D C,. Observation on scale chromatophore and body colors' genesis of carp and cruscian carp. Acta Hydrobiologica Sinica, 2007, 31(1): 67–72 [徐伟, 李池陶, 曹顶臣, 等. 几种鲤鲫鳞片色素细胞和体色发生的观察. 水生生物学报, 2007, 31(1): 67–72]
YAAR M, GILCHREST B A. Human melanocyte growth and differentiation: A decade of new data. Journal of Investigative Dermatology, 1991, 97(4): 611–617
Isolation, Culture and Identification of Polychromatic Midas cichlids () Melanophores and Xanthophores
SONG Hongmei1, ZHOU Kangqi2, TIAN Xue3, WANG Xuejie1, LIU Chao1, LIU Yi1, MU Xidong1, HU Yinchang1①
(1. Pearl River Fisheries Research Institute, Chinese Academy of Fishery Sciences; Key Laboratory of Leisure Fisheries, Ministry of Agriculture and Rural Affairs; Guangdong Modern Leisure Fisheries Engineering Technology Research Center, Guangzhou, Guangdong 510380, China; 2. China Southwest University, Key Laboratory of Freshwater Fish Resources and Reproductive Development, Ministry of the Ministry of Education, Chongqing 402460, China; 3. College of Fisheries, Engineering Technology Research Center of Henan Province for Aquatic Animal Cultivation, Henan Normal University, Xinxiang, Henan 453007, China)
This study aimed to investigate the methods of isolation, culture, and identification of melanophores and xanthophores in polychromatic midas cichlids () and to thus observe the growth characteristics of pigment cells in fish. In this study, the caudal fin, scales, and skin tissues ofwere used as the experimental material. To separate the pigment cells, the tissues were digested in trypsin solution and collagenase solution for 6 h and 12 h, respectively,at 25℃~28℃. After the cell suspension was filtered through a stoma nylon net, xanthophores and melanocytes were separated and collected by means of 35%-45%-55% Percoll multilayer density gradient centrifugation. K-SFM medium was used for primary cell culture and cell purification to inhibit the growth of fibroblasts and keratinocytes. DMEM supplemented with phorbol ester (TPA), double resistance, and bEGF,., was used for the subculture of pigment cells. The morphology of the cells was observed by L-DOPA staining and transmission electron microscopy, and the cells were identified by molecular markers. The results showed that when the cells were separated, the melanocytes were located at the boundary of the 45% and 35% Percoll reagent layers, while xanthophores were located on top of the 35% Percoll layer. Both pigment cells condensed into flocculation. Transmission electron microscopy showed that the melanocytes contained a large number of melanosomes, while the yellow cells contained the MTX and carotenoid vesicles. The melanocytes were positive under L-DOPA staining. The 10th generation of pigment cells was used for PCR detection of the body color related genes,, and, which showed strong specificity of gene amplification bands, indicating that the two pigment cells obtained had good biological activity.In this study, methods for the isolation, culture, and identification of melanophores and xanthophores of polychromatic midas cichlids were successfully established, providing a cell model for further research on the molecular mechanisms of body color cell differentiation and body color formation in fish.
; Pigment cell; Separation; Cultivation; Identification
HU Yinchang, E-mail: huyc22@163.com
S917.4
A
2095-9869(2021)06-0053-08
10.19663/j.issn2095-9869.20200617001
http://www.yykxjz.cn/
宋红梅, 周康奇, 田雪, 汪学杰, 刘超, 刘奕, 牟希东, 胡隐昌. 橘色双冠丽鱼黑色素细胞和黄色素细胞的分离、培养与鉴定. 渔业科学进展, 2021, 42(6): 53–60
SONG H M, ZHOU K Q, TIAN X, WANG X J, LIU C, LIU Y, MU X D, HU Y H. Isolation, culture and identification of polychromatic midas cichlids () melanophores and xanthophores. Progress in Fishery Sciences, 2021, 42(6): 53–60
胡隐昌,研究员,E-mail: huyc22@163.com
2020-06-17,
2020-08-20
*国家自然科学基金青年基金(802037)、广东省自然科学基金(2020A1515010304)和广州市科技计划项目(202002030047)共同资助[This work was supported by the National Natural Science Foundation of China for Young Scholars (802037),(2020A1515010304), and(202002030047)]. 宋红梅,E-mail: shm1227@126.com
(编辑 冯小花)
