基于脊髓中枢敏化内热针干预慢性软组织疼痛机制研究*
2021-12-30段轶轩张照庆骆小娟
段轶轩 张照庆 尹 晶 骆小娟 武 欢 夏 杨 胡 飞
(武汉市第三医院 武汉大学附属同仁医院疼痛康复科,武汉 430060)
慢性软组织疼痛是威胁健康最主要的原因之一,并且可导致劳动能力的丧失。来自欧美国家的流行病学调查显示,成年人慢性软组织疼痛的发生率高达30%左右。慢性疼痛还可伴随焦虑、抑郁以及饮食和睡眠障碍等,严重影响疼痛病人的生活质量,给病人及其家庭和社会带来严重负担。内热针疗法在腰椎间盘突出症、腰椎管狭窄、顽固性腰腿疼痛、各型颈椎病、粘连性肩关节囊炎等慢性软组织疼痛的治疗中疗效显著,可以有效、安全地缓解各种慢性肌肉、软组织疾病引起的痛觉过敏[1~5]。但其治疗机制值得探讨,目前已经从外周敏化,如炎性反应、抗氧化应激反应的角度进行了一系列的研究,发现其可通过降低TNF-α、IL-1β及IL-6 等炎性细胞因子的表达,改善局部炎症反应,通过降低p-Akt和NF-κB蛋白水平,抑制Akt/NF-κB信号通路的激活,促进组织损伤的修复[6]。但当机体发生慢性软组织疼痛时,躯体感觉通路被激活,除了外周炎性损伤,中枢系统也会发生一系列的复杂变化。其中中枢敏化是损伤后超敏感性疼痛的主要原因,也是慢性软组织疼痛中痛觉过敏的主要病理基础。中枢敏化是指脊髓背角伤害性突触信息传递增强导致痛觉敏感[7]。
内热针疗法是一种沿革于温针灸而优于温针灸的新式针灸,采用针芯电阻丝均匀加热,可对针尖温度进行电脑温控,通过针具的内部改良和电流的结合产生温控效应,可有效解决受热不均匀、皮肤烧烫伤的问题,同时也不再产生油烟、气味等污染环境。其又沿袭了温针灸温经散寒、活血通络的作用,又能通过对软组织的密集针刺改善肌肉痉挛和局部组织血液循环,是对温针灸疗法的沿袭和传承,又是对它的一种革新。
本研究采用内热针干预慢性软组织疼痛大鼠,通过观察L4-6脊髓节段相关递质的表达及脊髓背角神经元突触长时程增强效应的影响,来探讨内热针干预慢性软组织疼痛的脊髓中枢敏化的机制。以期为临床治疗提供实验依据,最终寻找到一种有效的治疗慢性软组织疼痛的方法,减轻病人及其给家庭和社会带来的严重负担。
方 法
1.实验材料
实验动物:清洁级雄性SD大鼠45只,3月龄,体重 300~350 g,由三峡大学动物实验中心提供,本研究涉及的所有动物操作,均严格遵循三峡大学动物伦理委员会的相关规定,动物使用许可证编号:SYXK(鄂)2017-0061。同一条件下分笼饲养,所有大鼠经适应性饲养1周后开始实验。
实验仪器:KF型内热针治疗仪(济南佳科医疗科技有限公司)。内热针总长6 cm,针柄长2 cm,针体长4 cm,针体产热长度4 cm(济南佳科医疗科技有限公司)。机械痛测痛仪(美国Model2390CE,IITC, Life science, Inc)。热痛测痛仪(美国Mode390,IITC, Life science, Inc)。Z2J-MB-NCC08 型肌电图仪(上海诺诚医疗器械有限公司)。钨丝微电极电极阻抗(美国Stoelting)。TGL-16M 台式高速冷冻离心机 (Biobase)。DR-200B 酶标仪(无锡华卫德朗)。JY600E 电泳系统 (Junyi)。110V-240V 凝胶成像仪(上海勤翔)。KZ-II型匀浆仪(康涛科技)。D3024R型台式高速冷冻型微量离心机 (DragonLab)。SLAN荧光定量PCR仪(上海宏石医疗器械有限公司)。SW-CJ-1FD超净工作台(苏净安泰)。NanoDrop2000超微量分光光度计 (Thermo)。FBZ2001-up-p标准试剂型纯水仪(青岛富勒姆科技有限公司)。
实验试剂及耗材:NC膜 (PALL),脱脂奶粉(BD),彩色预染蛋白marker, ECL, SDS-PAGE蛋白上样缓冲液 (5x),PMSF,磷酸酶抑制剂,蛋白酶抑制剂 (Biosharp),BCA蛋白浓度定量试剂盒(碧云天),TRIZOL(天根生物),三氯甲烷、异丙醇、无水乙醇(国药集团化学试剂有限公司),Hy-Pure TMMolecular Biology Grade Water (HyClone),RevertAid First Strand cDNA Synthesis Kit (Thermo),FastStart Universal SYBR Green Master (Rox)(Roche),引物(天一辉远)。
2.实验方法
分组:将45只大鼠编号,用SPSS编程随机分为对照组、模型组、内热针组,每组15只。
模型制备:组织损伤模型参考 Mc Brier等[8]与王绪明[9]机械冲击挫伤软组织的造模方法,将实验大鼠称重编号后,自然饲养,用10%的水合氯醛以3 ml/kg腹腔内注射麻醉,麻醉后将大鼠俯卧位,四肢逐个固定于大鼠台上,防止大鼠移位。将左侧后肢腓肠肌处皮肤剪毛,直至可见大鼠肉色皮肤但不损伤皮肤为宜。用缓冲垫将左侧后肢垫起,以防止其悬空和下肢移位。将左后肢稍外旋,使其小腿后群肌置于胫骨、腓骨内侧,将固定器置于腓肠肌上方压紧,防止大鼠移动导致目标肌群移位。将实验打击器械中央杆上的500 g砝码(其中心穿过长为40 cm的圆形铁杆,铁杆末端对准固定器的中央)置于中央杆最高点处(25 cm处),从中央杆最高点处沿着铁杆自由下落,作用于下方的固定器上方,导致肌肉组织的一次性钝、挫伤,反复重复10次,造成大鼠组织急性钝、挫伤。造模后,不给实验大鼠任何治疗或处理,正常环境下常规饲养。
造模评价:造模后对大鼠进行行为学测定以初步判断造模成功与否,模型组及内热针组大鼠在造模后第1天精神状态均较萎靡,活动减少,足趾并拢卷曲,行走时有跛行倾向,左后肢拖地,重心偏右。可以此初步判断实验大鼠造模成功。经过15天,形成大鼠慢性软组织损伤动物模型。在进行取材后,取模型组及内热针组大鼠局部组织进行病理形态学观察,造模后左侧后肢腓肠肌肌肉肌腱接头处肌肉组织肌纤维部分断裂、结构紊乱、排列混乱、间隙变大、大量中央多角形萎缩的肌纤维、肌萎缩严重、有中央多核的胞浆呈泡沫状及炎性细胞浸润,以此证明造模成功(见图1)。否则排除,不纳入实验。

图1 慢性软组织疼痛大鼠左侧后肢腓肠肌肌肉肌腱接头处组织标本病理形态学改变(HE染色,× 200 Scale bar = 200 μm)Fig. 1 Pathomorphological changes of tissue specimens in muscle-tendon junction of left hind limb gastrocnemius in experimental rats with chronic soft tissue pain (HE staining, × 200 Scale bar = 200 μm)
处理方法:对照组:正常饲养,不采取任何干预措施。模型组:造模完成后正常饲养。内热针组:造模完成后,采用内热针治疗,常规麻醉固定,实验大鼠左侧损伤的组织皮肤消毒,参照华兴邦《常用动物动物腧穴图谱》定位,以“承山”为中心,与腓肠肌肌肉组织走行相平行的方向刺入损伤组织,针尖触及骨面,布针2~3枚,连接内热式针灸治疗仪,43℃恒温加热20 min后取针,按压止血并消毒。每周治疗1次,共治疗2次。
3.取材及指标检测
取材:将3组大鼠在内热针治疗完成后第4天用10%的水合氯醛(3 ml/kg)腹腔注射进行麻醉,将麻醉后的大鼠固定于解剖台上,固定方法同前。切开大鼠胸部皮肤,剪断剑突两侧肋骨,迅速暴露心脏。将动脉套管针从左心室插入升主动脉,止血钳固定套管针,在右心耳剪一小口,经套管针快速灌注4%多聚甲醛固定液,待右心耳处流出液清亮时再缓慢灌注500 ml固定液固定。灌流毕,取L4-6脊髓节段部分浸泡于4%多聚甲醛固定液,置于4℃冰箱过夜备用。
观察指标:观察大鼠行为学改变,机械缩足反射阈值 (mechanical withdrawal threshold, MWT) 及热缩足反射潜伏期 (thermal withdrawal latency, TWL)。采用Western Blot法观察慢性软组织疼痛大鼠L4-6脊髓节段离子型谷氨酸受体(iGluRs)相关亚单位NR1、NR2B、GluR1蛋白的表达。采用RT-PCR法观察慢性软组织疼痛大鼠L4-6脊髓节段离子型谷氨酸受体(iGluRs)相关亚单位NR1、NR2B、GluR1 mRNA的表达。采用电生理方法诱导检测脊髓背角神经元突触传递长时程增强 (long-term potentiation, LTP)。
指标检测:各组大鼠在造模完成后的第1天、6天、14天,分别测定大鼠的MWT。将待测大鼠置于透明的有机玻璃箱中,底为铁丝网。触觉用一匀速上升的触针测定。待其适应环境安静后,用仪器机械力触针垂直对准左后肢足底后外处。启动仪器,触针上升经底板网眼碰触测试部位,当触顶的机械力增大到一定程度时大鼠会因疼痛而抬起后足,仪器自动记录该部位受碰触后抬足或离开时触针所施碰触力单位。最大碰触力经测试定为50 g,运行时间5 s。共测量5次,每次间隔5 min,取5次数值的平均值。
在造模完成后的第1天、6天、14天,分别采用热板实验法检测各组大鼠TWL。将热平板温度设置为 (52±0.2)℃。待温度稳定后,将大鼠放置于热平板上,记录从接触平板到抬起右后足或舔右后足的间隔时间。测量时间上限值为30 s,共测量5次,每次间隔时间大于15 min,取5次数值的平均值。
NR1、NR2B、GluR1 蛋白及mRNA的表达:标本置于冰镇玻璃试管中,按每100 mg组织湿重3 ml的比例加入4℃的10%磺基水杨酸,在冰浴下用内切式匀浆机制备组织匀浆。采用分子生物学(Western Blot 和RT-PCR)方法,观察慢性软组织疼痛大鼠L4-6脊髓节段中枢敏化相关主要iGluRs相关亚单位NR1、NR2B、GluR1蛋白及mRNA的表达。
Western Blot法:用含1%苯甲基磺酰氟的RIPA蛋白裂解液提取细胞蛋白,将提取的蛋白溶液按照4:1比例加入5×蛋白上样缓冲液,沸水浴变性10 min,保存于-20℃冰箱备用。在10%十二烷基硫酸钠聚丙烯酰胺凝胶电泳上分离出目的蛋白,并转移到聚偏氟乙烯(即 PVDF)膜上,之后用 5%的脱脂牛奶封闭洗脱2 h,将PVDF膜在4℃一抗孵育过夜,次日室温下用TBST缓冲液洗涤3次。将PVDF 膜在二抗中孵育1 h,使用电化学发光法检测试剂盒曝光,上机分析目的条带的灰度值。其中NR1分子量为21 kDa,NR2B分子量为166 kDa,GluR1分子量为100 kDa,GAPDH分子量为36 kDa。
RT-PCR法:用Trizol裂解液提取各组细胞的总RNA,并将各组细胞 RNA终浓度调整为200 ng/μl。进行反转录,采用反转录20 μl体系,加入4 μl 5×Reaction Buffer缓冲液,1 μl Oligo (dT)18Primer (50 uM),1 μl Servicebio®RT Enzyme Mix, 12 μl RNase free water, 2 μl mRNA,混匀后在PCR仪上25℃保温300 s,42℃保温1800 s,之后85℃保温5 s终止反应,得到合成的cDNA。在无酶 PCR 管内加入5 μl 2×SYBR Green qPCR Master Mix, 0.2 μl Forward primer,0.2 μl Reverse primer, 2.6 μl Nuclease-Free Water, 2 μl cDNA。进行 PCR扩增,预变性95℃ 30 s。95℃ 15 s转60℃ 30 s 为1次循环,循环40次。熔解曲线为60℃~105℃。以GAPDH作为参照,检测各组细胞基因以及内参表达量,结果采用 2ΔΔCT法作定量分析。引物均由天一辉远生物科技有限公司提供,引物序列见表1。
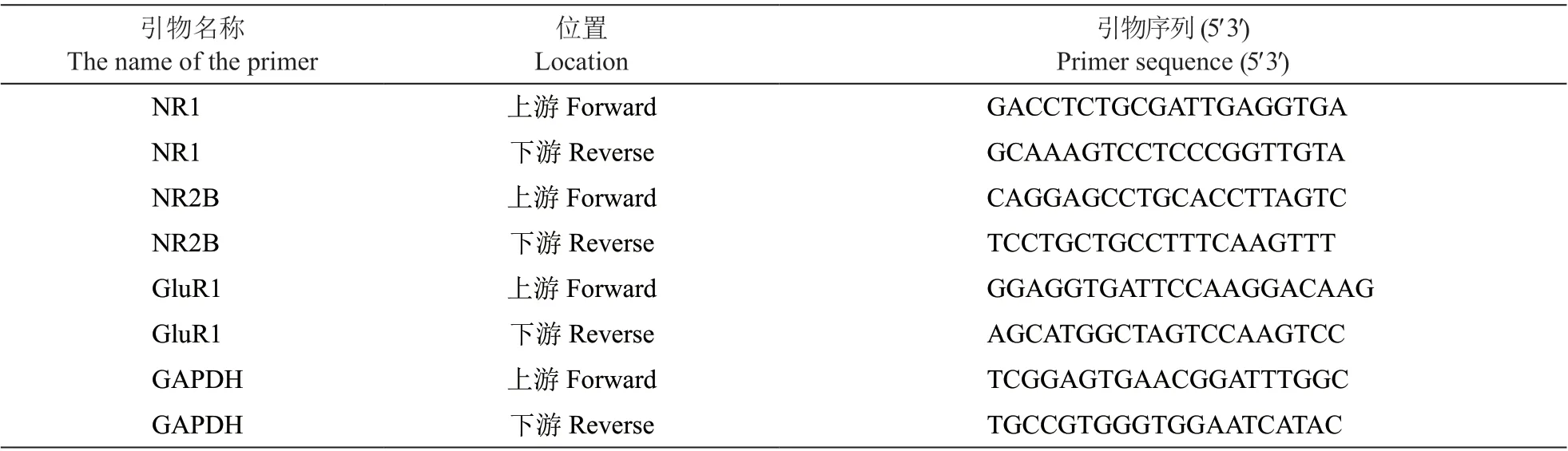
表1 目的基因及内参基因(GAPDH)所用引物及序列Table 1 Primers and sequences of target gene and internal reference gene (GAPDH)
脊髓背角C纤维、Aδ纤维及Aβ纤维群峰电位的记录和LTP的诱发:用钨丝微电极电极阻抗插入脊髓表面,在脊髓背角II层(插入深度为20~100 μm的范围内)寻找C纤维诱发场电位信号,在脊髓背角Ⅰ层(插入深度为0~20 μm的范围内)寻找Aδ纤维诱发场电位信号,在脊髓背角III-IV层(插入深度为100~220 μm的范围内)寻找Aβ纤维诱发场电位信号,用波宽0.15 ms,频率每5 min 1次的单脉冲波进行刺激,强度为引起群峰电位最大反应的60%左右时的刺激强度,在每层仔细寻找并记录最明显的脊髓背角C纤维、Aδ纤维及Aβ纤维诱发场电位信号, 为群峰电位。待群峰电位稳定10~15 min后,再行强直刺激,刺激参数为100 Hz、400个脉冲、60%最大刺激强度、波宽0.15 ms。给予强直刺激后群峰电位幅度增加20%,并持续30 min以上,即判定为长时程增强发生。整个实验过程中每 5 min观察记录1次,记录时间为2 h。电位经微电极放大器放大后,存入带有HXD-2000处理软件的计算机,再行分析处理。
4. 统计学分析
应用SPSS 22.0 统计软件进行数据分析,计量数据以均数±标准误(±SEM)表示,计量数据的组内比较采用重复测量方差分析。多组间计量数据比较采用单因素方差分析 (one-way ANOVA),方差齐时以Tukey 法和 SNK-q法多重比较,方差不齐时用 Dunnett T3检验。以P< 0.05为差异具有统计学意义。
结 果
1. 行为学观察
(1)大鼠行为观察:造模前各组大鼠精神状态可,步态正常,足趾伸展、运动及协调承重均正常。
模型组及内热针组大鼠在造模后第1天精神状态均较萎靡,活动减少,足趾并拢卷曲,行走时有跛行倾向,左后肢拖地,重心偏右。在内热针治疗完成后第4天对各组大鼠进行行为学观察,发现模型组大鼠精神可,但活动较对照组大鼠减少,足趾时有并拢卷曲,行走时稍有跛行倾向,左后肢无拖地,重心偏右。内热针组大鼠精神状态尚可,步态、足趾伸展、运动及协调承重与对照组相比未见明显改变。
(2)MWT:与对照组相比,模型组大鼠在相应各个时间的MWT均明显降低(P< 0.01),说明在慢性软组织疼痛模型造模成功后,大鼠MWT降低;内热针组的MWT在造模成功后的第14天较造模成功后的第1天明显升高(P< 0.01),较模型组明显升高(P< 0.01),与对照组第14天后相比差异无统计学意义,说明内热针治疗能有效提高机械痛阈(见图2)。

图2 各组慢性软组织疼痛大鼠MWT比较 (n = 15,±SEM)**P < 0.01,与本组造模后第1天相比; ##P < 0.01,与模型组造模后第14天相比;△△P < 0.01,与对照组同一时间相比Fig. 2 Comparison of MWT of each group of rats with chronic soft tissue pain (n = 15,±SEM)**P < 0.01, compared with the first day after modeling in this group; ##P < 0.01, compared with the model group on the 14th day after modeling; △△P < 0.01, compared with control group at the same time.
(3)TWL:与对照组相比,模型组大鼠在相应各个时间的TWL均明显降低(P< 0.01),说明在慢性软组织疼痛模型造模成功后,大鼠的TWL降低;内热针组的TWL在造模成功后的第14天较造模成功后的第1天明显升高(P< 0.01),较模型组明显升高(P< 0.01),与对照组第14天后相比差异无统计学意义,说明内热针治疗能有效提高热痛阈(见图3)。
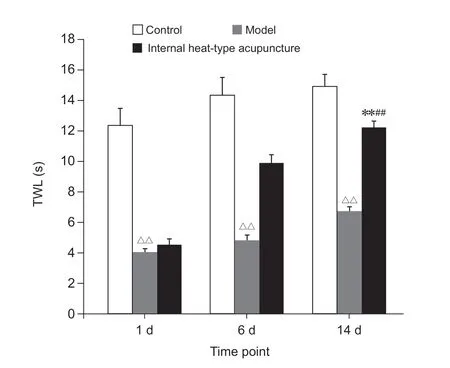
图3 各组慢性软组织疼痛大鼠TWL比较 (n = 15,±SEM)**P < 0.01,与本组造模后第1天相比; ##P < 0.01,与模型组造模后第14天相比;△△P < 0.01,与对照组同一时间相比Fig. 3 Comparison of TWL of each group of rats with chronic soft tissue pain (n = 15,±SEM)**P < 0.01, compared with the first day after modeling in this group; ##P < 0.01, compared with the model group on the 14th day after modeling; △△P < 0.01, compared with control group at the same time.
2. Western Blot法观察慢性软组织疼痛大鼠L4-6脊髓节段iGluRs相关亚单位NR1、NR2B、GluR1蛋白的表达
各组大鼠L4-6脊髓节段iGluRs相关亚单位NR1、NR2B及GluR1蛋白表达水平呈现内热针组<模型组,对照组与内热针组基本持平;与对照组相比,模型组NR1、NR2B及GluR1蛋白的表达明显升高 (P< 0.01);与模型组相比,内热针组NR1、NR2B及GluR1蛋白的表达明显降低 (P< 0.01);与对照组相比,内热针组NR1蛋白的表达明显升高(P< 0.01),但内热针组NR2B及GluR1的蛋白表达与对照组无明显差异(见图4)。
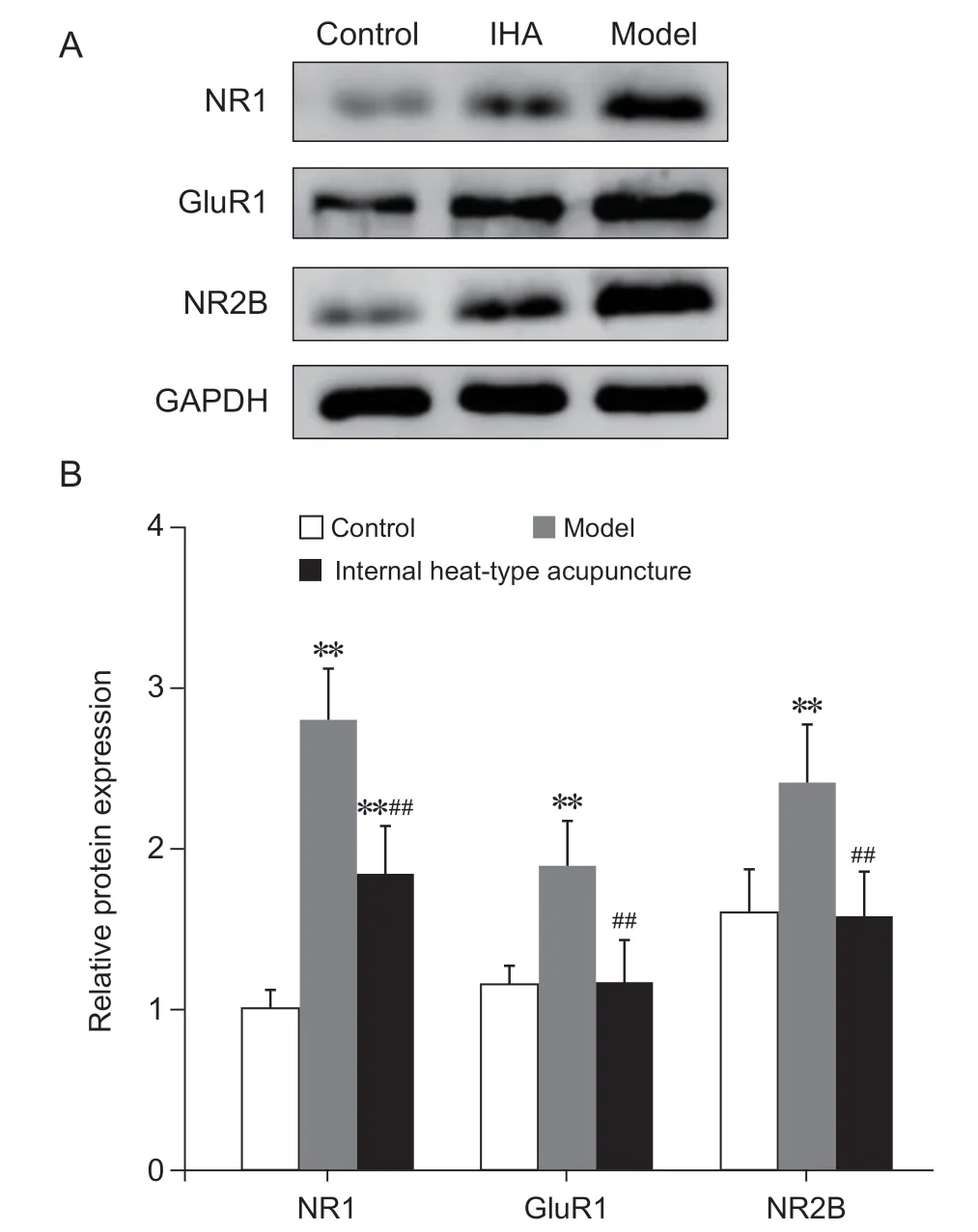
图4 各组慢性软组织疼痛大鼠L4-6脊髓节段NR1、NR2B及GluR1蛋白的表达(n = 5,±SEM)(A) WB检测L4-6脊髓节段NR1、NR2B、GluR1蛋白表达的条带图,GAPDH为内参条带;(B) 脊髓节段NR1、NR2B及GluR1蛋白表达的组间比较**P < 0.01,与对照组相比;##P < 0.01,与模型组相比Fig. 4 The expression of NR1, NR2B, and GluR1 proteins in L4-6 spinal cord segment of rats with chronic soft tissue pain in each group (n = 5,±SEM)(A) Showing the band of NR1, NR2B, and GluR1 protein expression in spinal cord segment L4-6 detected by WB, with GAPDH as the internal reference; (B)Comparison of protein expressions of NR1, NR2B and GluR1 in spinal cord segments between groups.**P < 0.01, compared with group control; ##P < 0.01,compared with group model.
3. RT-PCR法观察慢性软组织疼痛大鼠L4-6脊髓节段iGluRs相关亚单位NR1、NR2B及GluR1m-RNA的表达
各组大鼠L4-6脊髓节段NR1、NR2B及GluR1 mRNA表达水平为对照组<内热针组<模型组;与对照组相比,模型组NR1、NR2B及GluR1 mRNA的表达明显升高(P< 0.01);与模型组相比,内热针组NR1、NR2B及GluR1 mRNA的表达明显降低(P< 0.01);与对照组相比,内热针组NR1及GluR1 mRNA的表达升高 (P< 0.05),但内热针组NR2B mRNA的表达与对照组无明显差异(见图5)。
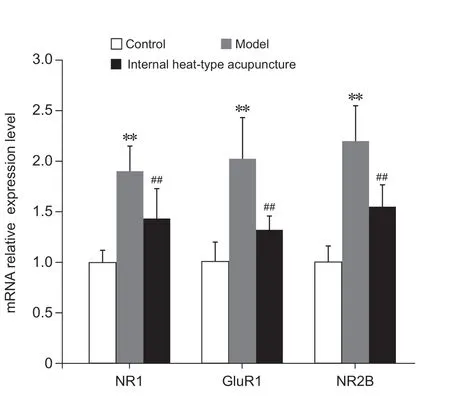
图5 各组慢性软组织疼痛大鼠L4-6脊髓节段NR1、NR2B及GluR1 mRNA的表达**P < 0.01,与对照组相比;##P < 0.01,与模型组相比Fig. 5 The expression of NR1, NR2B and GluR1 mRNA in L4-6 spinal cord segment of each group of rats with chronic soft tissue pain**P < 0.01, compared with group control; ##P < 0.01,compared with group model.
4. 脊髓背角神经元突触LTP情况
(1)脊髓背角C纤维LTP情况:以强直刺激脊髓背角各层的群峰电位幅度为100%计算,在对脊髓背角相关板层对C纤维诱发场电位信号,对照组未诱发出LTP效应,其在30 min时为刺激前的73.3%±12.8%;在60 min时为刺激前的72.1%±10.9%;在2 h时为刺激前的76.0%±16.8%。模型组脊髓背角强直刺激后其群峰电位幅度显著增高,在30 min时为刺激前的184.3%±22.1%,形成了长时程增强;在60 min时为刺激前的227.2%±58.9%;在2 h时为刺激前的169.9%±43.2%。内热针组未诱发出LTP效应,在30 min时为刺激前的74.3%±15.7%;在60 min时为刺激前的77.7%±20.5%;在2 h时为刺激前的71.1%±18.8%。模型组在强直刺激30 min、60 min及2 h时的群峰电位幅度明显高于对照组(P< 0.01);内热针组在强直刺激30 min、60 min及2 h时的群峰电位幅度明显低于模型组(P< 0.01);与对照组相比差异无统计学意义(见图6)。
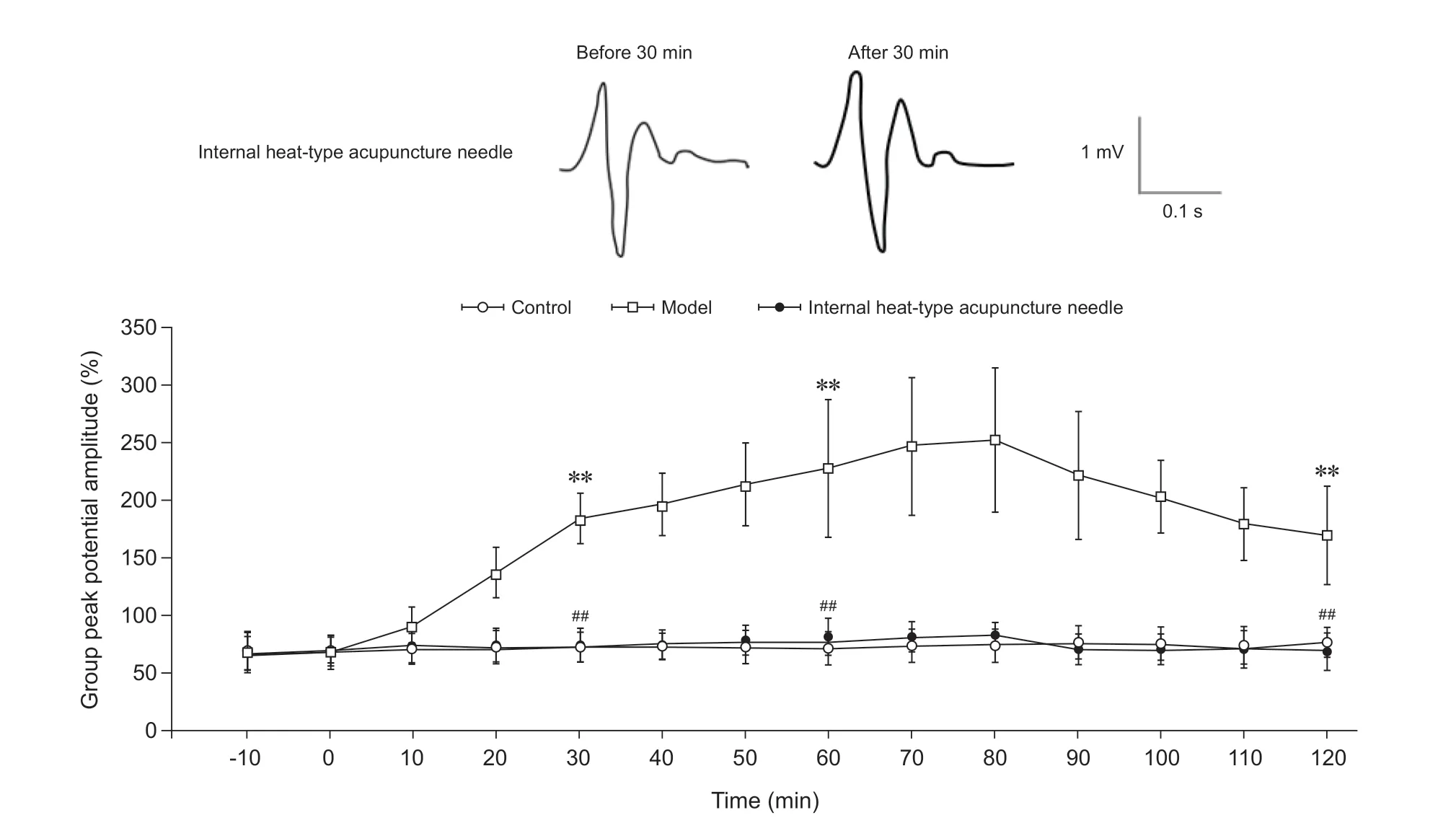
图6 各组大鼠脊髓背角C纤维长时程增强情况**P < 0.01,与对照组同一时间点相比;##P < 0.01,与模型组同一时间点相比Fig. 6 The LTP of C fibers in spinal dorsal horn of rats in each group**P < 0.01, compared to the group control at the same time point; ##P < 0.01, compared to the group model at the same time point.
(2)脊髓背角Aδ纤维LTP情况:以强直刺激脊髓背角各层的群峰电位幅度为100%计算,在对脊髓背角相关板层对Aδ纤维诱发场电位信号,对照组未诱发出LTP效应,其在30 min时为刺激前的59.8%±17.3%;在60 min时为刺激前的64.4%±19.0%;在2 h时为刺激前的62.9%±17.9%。模型组脊髓背角强直刺激后其群峰电位幅度显著增高,在30 min时为刺激前的141.0%±21.6%,形成了长时程增强;在60 min时为刺激前的178.3%±24.9%;在2 h时为刺激前的155.2%±20.1%。内热针组未诱发出LTP效应,在30 min时为刺激前的64.4%±13.9%;在60 min时为刺激前的72.6%±15.7%;在2 h时为刺激前的70.1%±16.5%。模型组在强直刺激30 min、60 min及2 h时的群峰电位幅度明显高于对照组(P< 0.01);内热针组在强直刺激30 min、60 min及2 h时的群峰电位幅度明显低于模型组(P< 0.01);与对照组相比差异无统计学意义(见图7)。
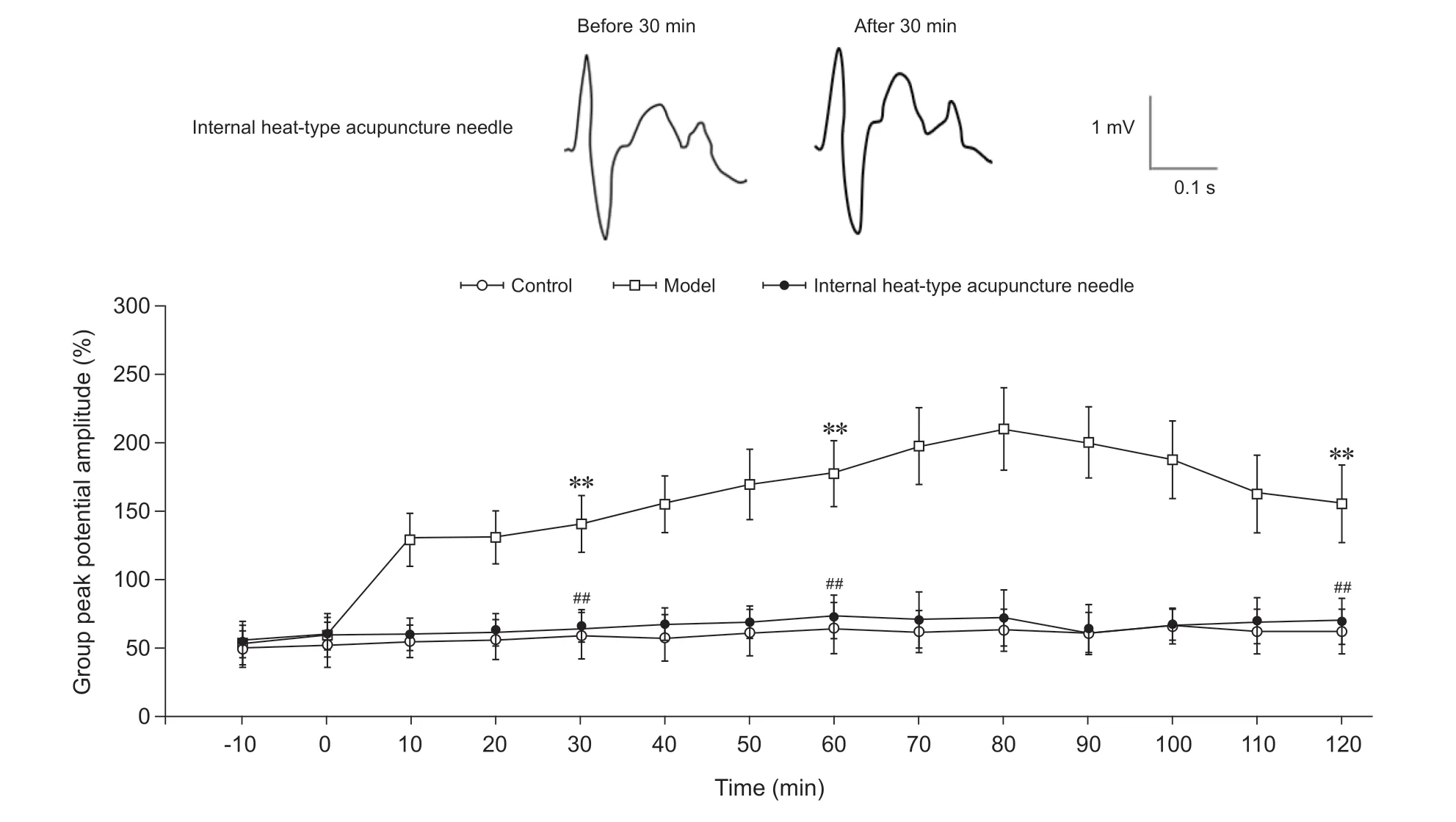
图7 各组大鼠脊髓背角Aδ纤维长时程增强情况**P < 0.01,与对照组同一时间点相比; ##P < 0.01,与模型组同一时间点相比Fig. 7 The LTP of Aδ fibers in spinal dorsal horn of rats in each group**P < 0.01, compared to the group control at the same time point;##P < 0.01, compared to the group model at the same time point.
(3)脊髓背角Aβ纤维LTP情况:以强直刺激脊髓背角各层的群峰电位幅度为100%计算,在对脊髓背角相关板层对Aβ纤维诱发场电位信号,对照组未诱发出LTP效应,其在30 min时为刺激前的67.3%±20.5%;在60 min时为刺激前的73.3%±16.8%;在2 h时为刺激前的72.9%±21.3%。模型组未诱发出LTP效应,且在脊髓背角强直刺激后其群峰电位幅度有所降低,其在30 min时为刺激前的70.7%±18.8%;在60 min时为刺激前的45.6%±11.0%;在2 h时为刺激前的49.8%±13.7%。内热针组在30 min时为刺激前的166.4%±31.3%,形成了长时程增强;在60 min时为刺激前的209.7%±42.4%;在2 h时为刺激前的158.9%±34.1%。模型组在强直刺激30 min、60 min及2 h时的群峰电位幅度与对照组相比差异无统计学意义;内热针组在强直刺激30 min、60 min及2 h时的群峰电位幅度明显高于模型组及对照组(P< 0.01,见图8)。
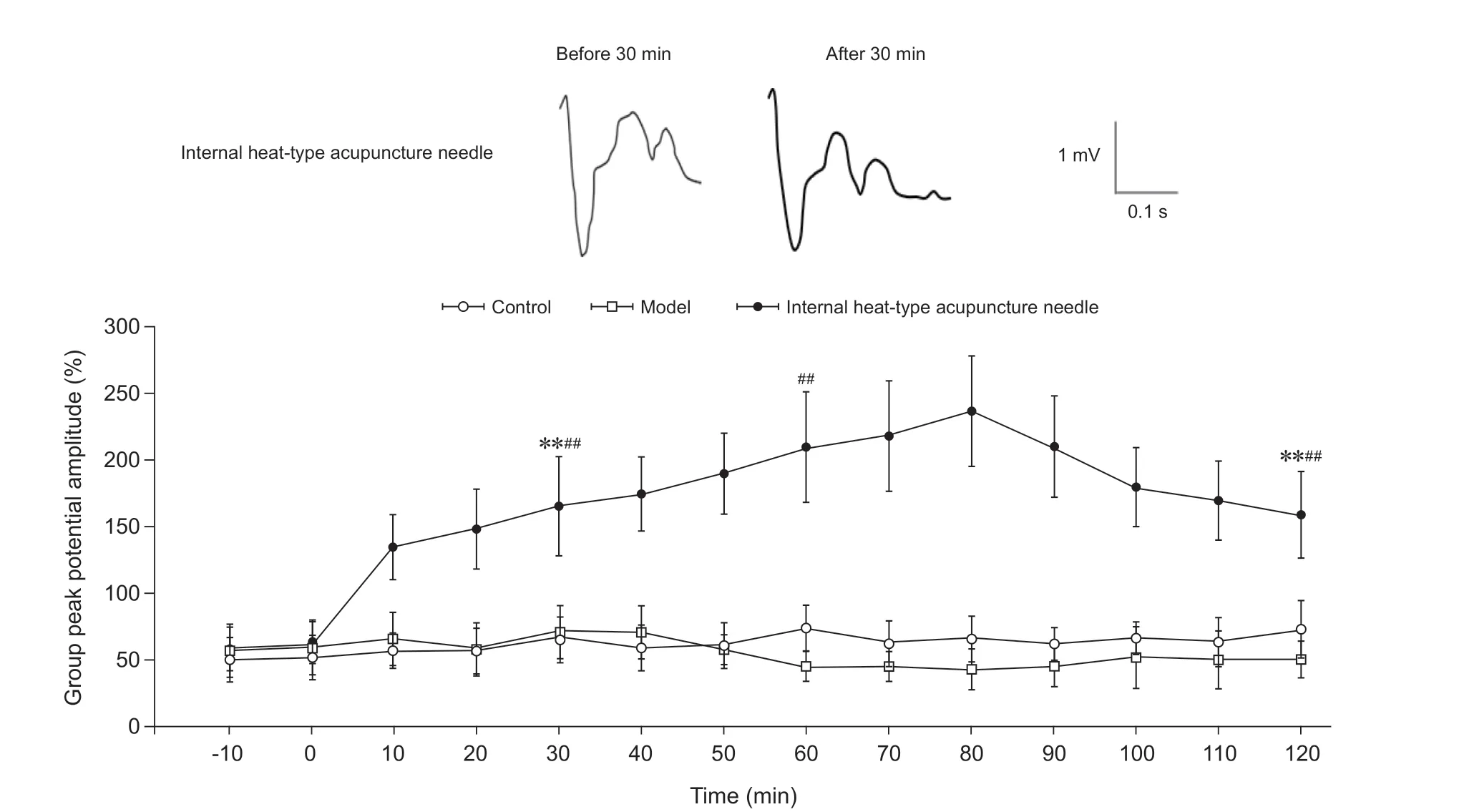
图8 各组大鼠脊髓背角Aβ纤维长时程增强情况**P < 0.01,与模型组同一时间点相比;##P < 0.01,与对照组同一时间点相比Fig. 8 The LTP of Aβ fibers in spinal dorsal horn of rats in each group**P < 0.01, compared to the group model at the same time point; ##P < 0.01, compared to the group control at the same time point.
讨 论
当机体发生慢性软组织疼痛时,躯体感觉通路被激活,除了外周炎性损伤,中枢系统也会发生一系列的复杂变化。其中中枢敏化是损伤后超敏感性疼痛的主要原因,也是慢性软组织疼痛中痛觉过敏的主要病理基础。中枢敏化是指脊髓背角伤害性突触信息传递增强导致痛觉敏感[7]。外周伤害性感受器的传入激发神经元兴奋性,释放兴奋性氨基酸、抑制性氨基酸和神经肽类神经递质,作用于脊髓神经元突触后受体,导致神经元突触后膜离子型谷氨酸受体 (iGluRs) 之NMDA 受体上调,而NMDA 受体的两个亚基NR1/2在脊髓背角NR1/2基磷酸化对脊髓中枢敏化至关重要。NR1/2 亚基磷酸化导致NMDA 受体通道Ca2+电流明显增加;钙离子活动增强进一步引发蛋白激酶C (PKC) 活化,激活下游的转录因子,使突触后表达更多的NMDA和AMPA受体,它们的过度激活导致了C类纤维传入末梢与脊髓背角伤害性神经元联结突触的可塑性,从而形成一个谷氨酸和天冬氨酸神经递质的正反馈回路,这种正反馈是神经元突触可塑性长时程增强效应LTP 形成的基础。而LTP是中枢敏化产生的重要基础[10~12]。
组织炎症和神经损伤等刺激使伤害性信息的初级整合中枢脊髓背角的神经组织结构发生变化,其中包括外周伤害性传入末梢在脊髓背角分布区域的变化[13]。正常生理情况下,外周Aβ类传入纤维终止于脊髓背角第III-IV层,传递低阈值的感觉神经信号(如触觉、压觉等);Aδ纤维多终止于第I、III层传递快痛信息;而C类纤维传入末梢则只终止在第II层即胶质层,主要传递慢痛信息。在针刺镇痛公认的闸门学说中认为,在脊髓背角内存在一种类似闸门的神经机制,粗纤维 (Aβ) 和细纤维(Aδ和C)的相对活动控制着痛觉信号的传入。Duan 等[14]的发现进一步充实了闸门学说。脊髓背角胶质中表达生长抑素 (somatostatin, SOM) 的兴奋性中间神经元和强啡肽 (dynorphin, Dyn) 的抑制性中间神经元,受来自躯体同一神经节段的 C/Aδ纤维和 Aβ纤维的拮抗性支配,C/Aδ纤维的传入激活 SOM+ 神经元,开放闸门,引起疼痛;Aβ纤维的传入激活 Dyn+ 神经元,抑制 SOM+ 神经元的活性,闸门关闭从而阻断疼痛传入。针刺通过激活 Aβ 纤维,关闭闸门起到对同源疼痛的抑制作用。
内热针疗法是一种新型治疗慢性肌肉损伤的方法,其有效地将传统的针刺治疗和热疗结合在一起,通过现代电子技术和工艺材料技术的改造,使其具有内部可加热、温度可控、热能集中等特点,在腰椎间盘突出症、腰椎管狭窄、顽固性腰腿疼痛、各型颈椎病、粘连性肩关节囊炎等慢性软组织疼痛的治疗中疗效显著,可以有效缓解各种慢性肌肉、软组织疾病引起的痛觉过敏。研究显示[15,16],针刺的刺激或艾灸的热能作用于穴位,通过分布于皮肤肌肉感觉神经元末梢的机械或温度感受器产生局部感受器电位并以电紧张的形式传播,当去极化达到阈电位水平,产生动作电位并沿感觉神经向中枢传导。在两个神经元之间,又借助突触结构相联系,根据信息传递媒质的不同,分为电突触或称缝隙连接和化学性突触。因此,电生理学方法和神经化学方法是研究针灸原理的两种重要的实验方法[17]。由于针灸效应的发挥需要从外周到中枢各级神经的参与,所以在体电生理学方法在针灸研究中又尤为重要[18]。而内热针来自于传统针灸,又结合了针刺及艾灸的治疗效应,更需要采用体电生理学的方法来揭示其治疗机制。
通过本实验研究发现,内热针组能降低慢性软组织疼痛大鼠L4-6脊髓节段离子型谷氨酸受体(iGluRs) 相关亚单位NR1、NR2B及GluR1蛋白及mRNA的表达;慢性软组织疼痛模型大鼠能诱发脊髓背角C纤维、Aδ纤维的LTP;不能诱发脊髓背角Aβ纤维LTP,甚至出现一定的长时程减弱。慢性软组织疼痛模型大鼠破坏了中枢神经系统原来的兴奋-抑制动态平衡,使神经元兴奋性和突触可塑性处于异常的活动增强或者减弱状态,导致了脊髓中枢的敏化,开放闸门,从而引发了疼痛。内热针治疗可有效抑制慢性软组织疼痛,可能与调节脊髓背角相关递质的表达,抑制脊髓背角C纤维、Aδ纤维的LTP,促进Aβ纤维的LTP,产生中枢敏化有关。本实验结果也与Xu等[18]和Zhu等[19]的研究结果相符。该研究在痛源一侧同神经节段水平,A 类纤维的激活就能产生明显的镇痛效应,蛇毒造成A纤维脱髓鞘损伤后,针刺镇痛效应显著削弱。而在痛源对侧或远离痛源部位针刺,只有激活C类纤维的的针刺强度才能明显抑制伤害性肌电反应,辣椒素阻断C纤维或切断高位颈髓造成脊髓化后该抑制效应消失,提示针刺通过兴奋外周不同直径的传入纤维,通过脊髓水平闸门控制或是触发DNIC系统的不同机制产生镇痛效应[19]。
本研究已初步通过电生理学方法和神经化学方法两种方法,推断内热针的治疗机制可能与其能调节L4-6脊髓节段相关神经递质的表达,抑制脊髓背角C纤维、Aδ纤维的LTP,促进Aβ纤维的LTP,激活Aβ纤维,关闭闸门,抑制慢性软组织疼痛有关。依据这一实验结果,是否可以推测内热针治疗缓解慢性软组织疼痛是通过L4-6脊髓节段iGluRs相关亚单位NR1、NR2B及GluR1等递质的介导,从而有效抑制脊髓背角C纤维、Aδ纤维的LTP,促进其对Aβ纤维的LTP,激活Aβ纤维,有效调节中枢神经系统的兴奋-抑制动态平衡?理论上来说,L4-6脊髓节段相关神经递质的表达与LTP的增强与减弱存在着相关性,但本研究尚不能确定其因果性,不能明确其间的传导反应过程,上下游转录因子、相关神经递质的正反馈回路等一系列信号传导级联反应,下一阶段需进一步基于脊髓中枢敏化系统,加入iGluRs抑制剂和激动剂对照,以判断iGluRs是否为介导脊髓背角相关纤维LTP的关键环节,从背根神经节和脊髓背角神经递质的上下游关系方面更深入地探讨其干预机制。
