春季低温对小麦花粉育性及粒数形成的影响
2021-12-17张玉雪苏盛楠李春燕丁锦峰朱朱新开郭文善
高 芸 张玉雪 马 泉 苏盛楠,3 李春燕,* 丁锦峰朱 敏 朱新开 郭文善
1 扬州大学江苏省作物遗传生理国家重点实验室培育点 / 粮食作物现代产业技术协同创新中心, 江苏扬州 225009; 2 太仓市农业机械技术推广站, 江苏太仓 215499; 3 太仓市城厢镇农业技术服务站, 江苏太仓 215499
温度是影响小麦生长发育的重要环境因素之一, 特别是春季3、4 月低温, 小麦正处于拔节、孕穗和开花的关键生育时期, 低温严重影响了小麦等农作物的生长[1-3]。我国黄淮平原冬麦区河南省和山东省霜冻害一般发生频率达 30%以上, 最高年份可达70%[4]。2016 年越冬冻害造成河南省柘城县小麦减产 5%~20%, 早春冻害造成减产5%~30%, 晚霜冻害造成减产15%~60%[5]。近55年来, 江苏省倒春寒平均每年发生3.8 次, 每次平均持续时间为3.2 d, 中级以上倒春寒一般发生率达22%以上[6]。2013 年3 月江苏省出现4 次寒潮和温度急剧下降的恶劣天气, 24 h 降温幅度高达21.2 ℃, 最低温度–0.7 ℃, 受灾严重地区兴化市小麦平均减产 25%左右[7], 可见春季低温是制约小麦生产的重要气候因素。
小麦生育中后期逆境胁迫会造成小麦每穗粒数和千粒重显著下降, 制约小麦产量的形成[8-9], 小麦在药隔形成期和孕穗期遇到低温影响更加显著, 每穗粒数的减少与温度的降幅和低温持续时间成正比[10-11]; 尤其是减数分裂期是小麦低温最敏感的时期, 该时段低温胁迫致每穗粒数减少是产量下降的主要原因[12]。
小麦幼穗发育的好坏决定了穗型的大小, 其发育过程受到外界温度的影响[13-14], 而粒数的多少往往与花粉育性和花后灌浆物质多少有关。导致小麦花粉育性降低的原因很多: 低温条件下小麦花粉母细胞减数分裂过程中出现非同源染色体配对的异常现象, 导致花粉败育[12]; 低温胁迫下植物体内活性氧过度积累, 花药抗氧化酶活性降低是导致小麦花药败育的又一原因[15-17]; 在不育系小麦方面的研究表明, 绒毡层程序化死亡过程的异常、花粉壁无法正常合成、以及花药中小孢子母细胞淀粉和脂类物质代谢异常等是温敏核不育小麦受温度影响致花粉败育的主要原因[18-20]。
一定强度的春季低温显著降低了小麦产量, 除穗数减少外, 每穗粒数下降也是减产的重要原因之一。但导致小麦粒数下降的原因是小孢子母细胞减数分裂的异常, 还是因为花药中营养物质代谢、绒毡层死亡过程的异常引起的花粉败育, 尚没有明确的结论。本试验以春性小麦品种扬麦16和半冬性小麦品种徐麦30为材料, 在小麦倒二叶出生期、孕穗期和开花期分别进行低温处理, 研究春季低温对小麦花粉育性和粒数形成的影响及其生理机制, 为提高小麦抗低温能力、确保小麦稳产提供理论依据和技术支持。
1 材料与方法
供试品种为春性品种扬麦16 和半冬性品种徐麦30, 试验于2015—2017 年在扬州大学江苏省作物遗传生理实验室盆钵试验场进行。试验用土为轻壤土, 两年度有机质含量平均为14.61 g kg–1, 碱解氮含量为52.35 g kg–1, 速效磷含量为37.35 g kg–1,速效钾含量为96.51 g kg–1。采用盆栽种植, 盆钵高26 cm, 上口直径26 cm, 底面直径18 cm, 底部一共8 个排水孔。每盆装土12 kg, 基施尿素0.85 g及复合肥(15% N、15% P2O5和15% K2O) 3.5 g, 浇等量水自然沉实后播种, 覆土1 kg。四叶期追施壮蘖肥, 每盆施尿素0.35 g; 叶龄余数2.5 时追施拔节肥, 每盆施尿素0.4 g 和复合肥3.33 g。2015 年于10 月31 日播种, 2016 年于11 月18 日播种, 每盆播种12 粒, 四叶期定苗, 每盆留生长一致植株8 株。在人工智能控温室中进行低温处理(温度误差±0.5 ℃, 大气相对湿度 70%, 光量子通量密度800 µmol m–2s–1)。
1.1 试验设计
于倒二叶出生期(倒二叶露尖1~2 cm)、孕穗期(剑叶全展, 剑叶与倒二叶叶枕距2~4 cm)和开花期(小麦中部小穗开出1~2 朵小花)选取生长一致的盆钵分别进行低温处理。倒二叶出生期、孕穗期、开花期昼夜低温分别为 5℃/–3℃、 8℃/–1℃和12℃/4℃。每个处理时间均设24 h 和48 h 两个水平,盆钵土壤水分为田间持水量的60%~70%, 处理结束后移至自然条件下生长至成熟, 以室外自然生长的植株为对照。
1.2 测定内容与方法
1.2.1 花粉母细胞减数分裂观察 取处于减数分裂期的麦穗(剑叶与倒二叶叶枕距为3~4 cm)中下部花药, 用卡诺固定液固定, 置–20℃冰箱保存。试验时将花药取出, 置于载玻片上, 加少许醋酸洋红,用解剖针迅速将花药捣碎, 盖上盖玻片, 在酒精灯上烤一下, 在显微镜下观察, 挑选适期的花粉母细胞。在盖玻片一侧加入适量的45%醋酸, 在另一侧放入吸水纸置换出染液, 使细胞质背景颜色变浅。在液氮中放置数秒(直到声音变弱), 用刀片迅速揭去盖玻片, 晾干。滴加4%多聚甲醛500 µL 静置5 min, 分别用 70%、90%和 100%酒精脱水。滴加DAPI10 µL, 盖上盖玻片, 压片, 荧光显微镜(Olympus BX60)放大倍数为100 倍进行观察, 使用Olympus cellSens Dimension 软件照相。
1.2.2 花药细胞化学观察 花药细胞化学观察方法参照李东霄[20-21]。小麦抽穗后, 取小麦中下部花药, 一半花药通过醋酸洋红压片确定二核期和三核期, 另一半用0.1 mol L–1(pH 7.4)磷酸盐缓冲液配置的含3%戊二醛固定液4℃固定8 h, 再用磷酸缓冲液漂洗3 次, 乙醇逐级脱水。用LR White 树脂渗透、包埋、聚合。选用Leica EM UC7 型(德国)超薄切片机切1.2 μm 的半薄切片, 高碘酸–希夫反应鉴别红色多糖物质, 考马斯亮蓝R-250 鉴别蓝色蛋白质类物质, 利用Leica DM 2500 型(德国)正置生物显微镜进行观察。
1.2.3 花粉活性观察 于开花期取麦穗中下部花药, 采用 I2-KI 染色制片, 采用蔡司显微镜(Axio Imager D2 型)观察花粉发育情况, 统计花粉败育率。
1.2.4 穗部性状调查 于成熟期每个处理取5 盆,调查每个处理每穗结实小穗数、退化小穗数、每小穗结实粒数和每穗粒数。
1.3 数据分析
本文数据采用Microsoft Excel 2010、DPS 6.55等软件进行数据的计算、绘图、统计分析。因2015—2017 两年度小麦生长季各项指标变化趋势基本一致, 本文重点以2016—2017 年数据进行详细分析。
2 结果与分析
2.1 春季不同时期低温对小麦花粉育性的影响
2.1.1 孕穗期低温对小麦花粉母细胞减数分裂的影响 与自然生长对照小麦花粉母细胞减数分裂后期I、末期I 和末期II 相比较(图1-1~3), 孕穗期8 ℃/–1℃低温处理48 h, 2 个品种小麦花粉母细胞减数分裂过程均观察到减数分裂异常情况。扬麦16 花粉母细胞减数分裂后期I 出现多条落后染色体, 组成染色体桥(图1-4); 在末期I 子细胞染色体排列散乱, 细胞周围出现微核(图1-5); 在末期II 细胞周围出现微核(图1-6)。徐麦30 花粉母细胞减数分裂期后期I 出现染色体桥和落后染色体(图1-7, 8); 在末期II 出现大小不一致的子细胞(图1-9), 表明孕穗期低温影响小麦花粉母细胞的减数分裂过程。
2.1.2 春季不同时期低温对小麦花药发育的影响
自然生长的2 个品种小麦花药中, 大液泡减小或者消失, 细胞质内含物积累正常, 在二胞花粉期,多糖物质淀粉和蛋白质含量逐渐增加(图2-A, D 和图4-A, D); 在三胞花粉期, 绒毡层降解情况较好,少有残留的痕迹(图3-A, D 和图5-A, D); 在小麦倒二叶出生期和孕穗期低温处理48 h 后, 处在二胞花粉期的花药, 大液泡分解程度较低, 细胞质内含物未增加, 颗粒物较少, 淀粉和蛋白质积累量低, 形似空壳, 花粉细胞出现较多不规则的畸形状态(图2-B, C, E, F 和图4-B, C, E, F); 三胞花粉期绒毡层细胞明显延迟分解, 有较多残留(图3-B, C, E, F 和图5-B, C, E, F)。倒二叶出生期低温处理后小麦花药中淀粉和蛋白质的积累量均低于孕穗期低温处理, 淀粉和蛋白质代谢异常的现象更显著(图3 和图5)。这些结果说明倒二叶出生期和孕穗期低温胁迫均会导致花药中淀粉、蛋白质代谢异常和绒毡层降解延迟, 影响了小麦花粉的正常发育。2 个品种均表现为倒二叶出生期低温处理对花药淀粉和蛋白质代谢的影响程度大于孕穗期低温处理。
2.1.3 春季不同时期低温对小麦花粉粒活性的影响
开花期通过碘染色观察花粉粒活性, 扬麦16 和徐麦30 在倒二叶出生期低温处理后败育率最高, 变化范围在23.6%~29.6%, 其次是孕穗期, 变化范围在13.6%~19.8%, 开花期败育率仅为7.0%~16.6%(表1)。3 个时期低温处理后花粉粒败育率均表现为徐麦30 高于扬麦16, 并且随低温处理时间的延长败育率增加。
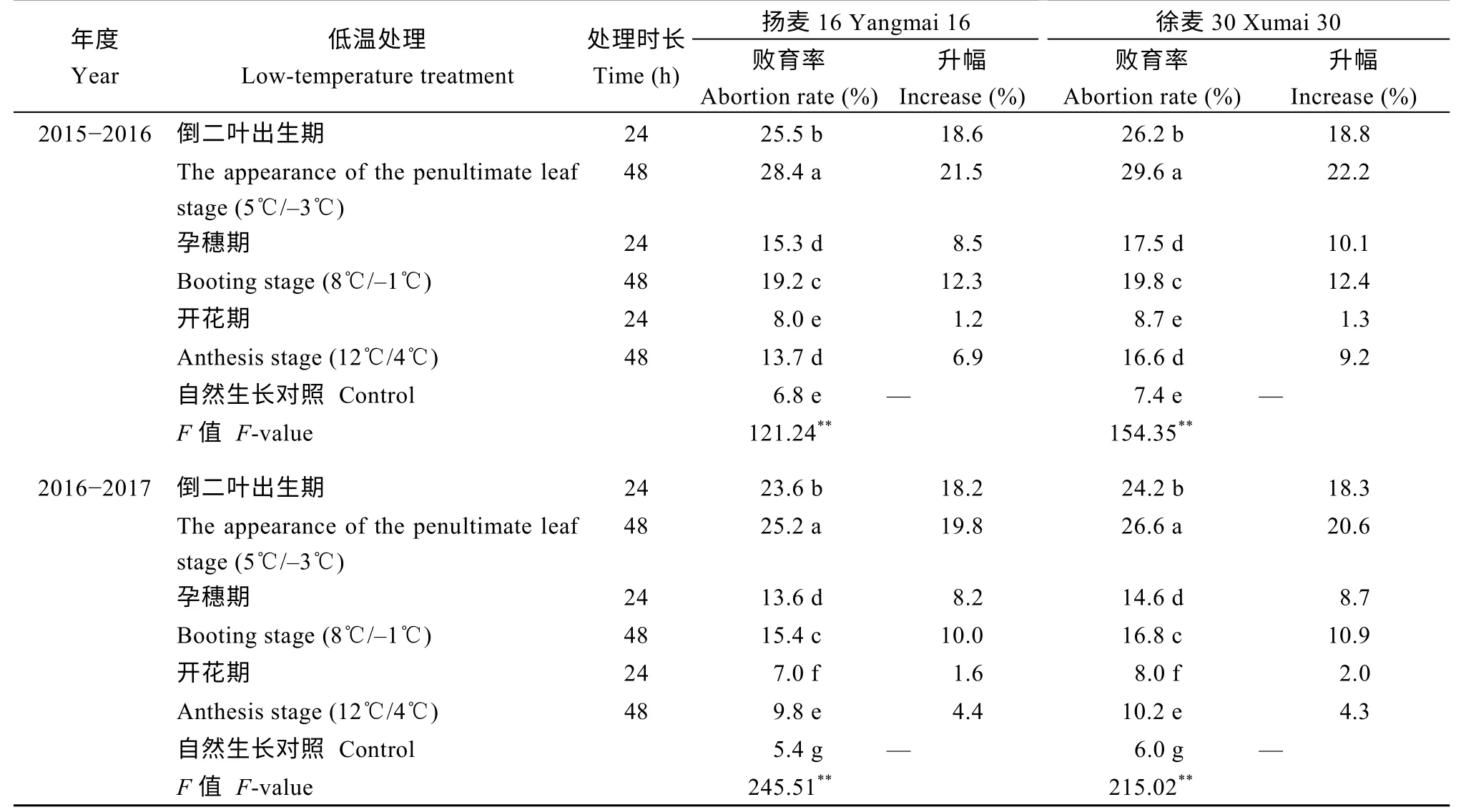
表1 春季不同时期低温对小麦花粉粒活性的影响Table 1 Effects of low temperature in spring at different stages on wheat pollen activity
2.2 春季不同时期低温对穗部性状的影响
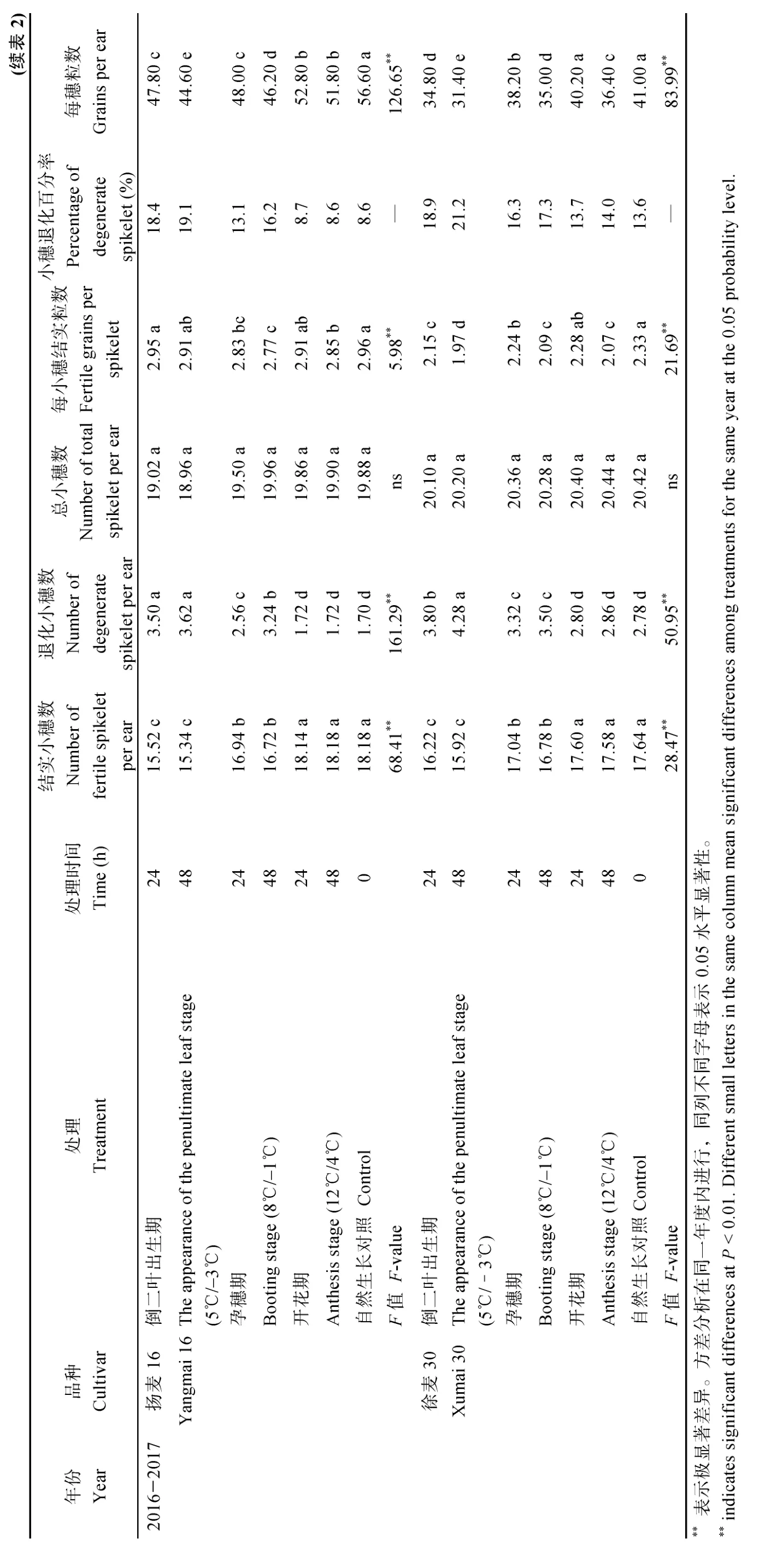
倒二叶出生期和孕穗期低温胁迫后, 小麦结实小穗数和每小穗结实粒数均显著低于自然生长对照, 且小穗退化百分率随着低温胁迫时间的延长呈上升趋势(表2)。在两年间, 扬麦16 倒二叶出生期低温处理24 h 和48 h 后平均小穗退化百分率分别为17.9%和18.7%, 孕穗期低温处理24 h 和48 h 后平均小穗退化百分率分别为14.0%和16.4%, 开花期低温处理扬麦16 退化小穗数略高于自然对照处理, 但无显著差异; 徐麦30 在两年间倒二叶出生期低温处理24 h 和48 h 之后平均小穗退化百分率分别为22.0%和24.0%, 孕穗期低温处理24 h 和48 h 后平均小穗退化百分率分别为19.9%和21.2%,徐麦30 开花期低温处理导致退化小穗数显著高于对照处理, 说明徐麦30 在开花期对低温反应更敏感。2 个品种3 个时期低温处理后, 每小穗结实粒数均低于自然生长对照处理, 且随低温处理时间的延长而减少。相关性分析表明, 2 个品种两年间花粉败育率与小穗退化数均成显著正相关, 2015—2016年, 扬麦16 相关系数0.935**, 徐麦30 相关系数0.780*, 2016—2017 年, 扬麦16 相关系数0.848*, 徐麦30 相关系数0.980**, 退化小穗数随着花粉败育率的增加呈上升趋势(图6)。
3 讨论
3.1 春季低温对小麦花粉育性的影响
花药的发育情况直接影响小麦粒数的形成。有研究表明, 不育小麦的花药异常情况大多出现在小孢子母细胞期、单核靠边期和二胞花粉期[22-23], 小麦剑叶与倒二叶叶耳距为0~4 cm 时幼穗中小孢子母细胞处于减数分裂的盛期, 对零度以上低温最为敏感, 低温导致后期出现落后的染色体、后期或末期染色体仍停留在赤道板上等异常现象[24]。本研究结果亦表明, 孕穗期低温对2 个品种小麦花粉母细胞减数分裂有显著影响, 低温处理后的小麦花粉母细胞出现染色体配对紊乱、减数分裂染色体不同步、子细胞差异明显等异常现象。赵莎等[25]研究表明,败育小麦农垦58 花药的绒毡层细胞提前启动细胞程序性死亡, 但解体过程缓慢, 使单核花粉无法获得所需的营养物质而败育。温敏核不育小麦花药在二胞花粉后期, 细胞质中内含物质不持续增加, 淀粉粒积累终止与花粉败育有关[20]。本试验研究结果表明, 倒二叶出生期和孕穗期低温处理后的花药中,绒毡层在三胞花粉时期仍有较多残留的痕迹, 说明绒毡层没有适时解体, 影响了花粉发育过程中营养物质的供给。同时亦观察到2 个时期低温处理均降低了花粉细胞内淀粉粒数量和蛋白质的积累量, 使得花粉在发育过程中没有充足的养分供应, 不能进一步发育为成熟的花粉粒。与温敏核不育花粉相比,本研究中低温处理仅使花粉的内含物质积累量下降,并没有出现非常严重的空泡化现象, 这也是低温处理后结实率并没有显著下降的原因。有研究表明,小麦拔节期、孕穗期低温显著降低叶片的光合能力[26-27], 减少植株干物质积累[28]。本研究中倒二叶出生期低温处理对花药中淀粉和蛋白质积累的影响大于减数分裂期低温, 这可能是由于倒二叶出生期低温处理对叶片伤害重, 影响了叶片的光合作用,也间接影响了花粉的营养供给, 导致花粉败育率显著增加; 同一时期随着低温处理时间的延长, 败育率增高。有研究指出, 冬性小麦低温诱导的抗冻害基因在从营养生长期转向生殖生长期的过程中表达量是显著下降的, 遇春季低温抗寒性下降[29], 本试验中2 个品种间徐麦30 败育率大于扬麦16, 这个结果与每穗结实粒数下降的结果一致, 进一步说明半冬性品种徐麦30 在小麦后期低温的敏感性高于春性品种扬麦16。
从花药原基分化到成熟花粉粒形成的过程中,花粉母细胞减数分裂和花药绒毡层正常发育是形成可育花粉的关键[30]。本试验研究结果表明, 孕穗期低温处理后出现了花粉母细胞减数分裂异常, 花药细胞畸形以及绒毡层分解迟缓等现象, 花粉育性亦降低。在拟南芥中RCK 基因发生突变后同源染色体联会和交叉紊乱, 二价体数目减少, 出现单价体,花粉母细胞减数分裂异常, 导致花粉败育[31]。在绒毡层发育的过程中, 多个受体蛋白激酶[32]、转录因子MYB (v-myb avian myeloblastosis viral oncogene homolog)结构域蛋白[33]、锌指蛋白中的PHD-Finger结构域蛋白[34]等均参与调控, 小麦雄性不育与育性相关基因表达也有关, 早在减数分裂期育性基因的表达就出现差异, 且外界温度对不育基因表达量有调控作用[35], 本试验中低温胁迫是否导致基因或者蛋白调控过程异常, 从而影响花粉母细胞减数分裂和绒毡层的程序性死亡的启动致花粉败育, 尚需从分子生物学角度开展深入研究。

?
?
3.2 春季低温对小麦每穗粒数的影响
春季低温造成穗数和每穗粒数显著下降[11,13,36]。拔节期和孕穗期低温对穗数和每穗粒数的影响超过对粒重的影响, 特别是在孕穗期低温处理后每穗粒数下降达55.1%, 是产量降低的主要原因[37]。Subedi等[38-39]研究结果表明, 抽穗后低温显著降了小麦结实率, 孕穗至开花期低温导致小麦每穗粒数显著下降, 产量下降了35%~78%。本试验研究结果亦表明,倒二叶出生期和孕穗期低温均显著降低了小麦结实小穗数和每穗粒数, 增加了退化小穗数, 且随着低温时间的延长, 小穗退化百分率呈上升趋势, 每穗粒数呈下降趋势。开花期低温略降低了结实小穗数和每穗粒数。相关分析表明花粉败育率与退化小穗数呈显著正相关, 这也进一步说明倒二叶出生期和孕穗期低温引起花粉败育是此期低温致每穗粒数下降的主要原因。生产中这2 个生育时期若发生低温天气值得关注, 后续将开展春季低温的避灾减灾调控途径研究, 以提高小麦抗春季低温逆境能力, 实现稳产增产。
4 结论
小麦孕穗期低温导致花粉母细胞减数分裂出现染色体配对紊乱等异常现象, 倒二叶出生期和孕穗期低温延迟了花药绒毡层的降解, 花药中多糖物质淀粉和蛋白质的积累量下降, 花粉发育因营养物质不足败育率增加, 开花期低温处理花粉败育率略有下降。3 个时期低温处理均降低小麦每小穗结实粒数和每穗粒数, 并随低温处理时间的延长呈上升趋势, 以倒二叶出生期低温处理穗粒数降低的幅度最大, 其次是孕穗期, 开花期穗粒数降低幅度最小。相关分析表明, 春季低温导致小麦花粉育性下降是每穗粒数降低的主要生理原因。
