海藻糖对乳酸菌保护机制的研究进展
2021-12-17杨友馨王瑞雪王俊国
杨友馨,王瑞雪,王俊国
(内蒙古农业大学食品科学与工程学院,乳品生物技术与工程教育部重点实验室,农业农村部奶制品加工重点实验室, 内蒙古 呼和浩特 010018)
海藻糖又称为漏芦糖、蕈糖,是由两分子葡萄糖通过α,α-1,1糖苷键连接而成的一种非还原性双糖,19世纪早期Wiggers在研究黑麦麦角的过程中分离提取出了海藻糖[1],之后被发现广泛存在于细菌、酵母、真菌、藻类以及一些昆虫、无脊椎动物和植物中[2]。海藻糖的结构式为α-D-吡喃葡糖基-α-D-吡喃葡糖苷,通常以二水化合物的形式存在,分子式为C12H22O11·2H2O,相对分子质量为378.33[3],结构式如图1所示。
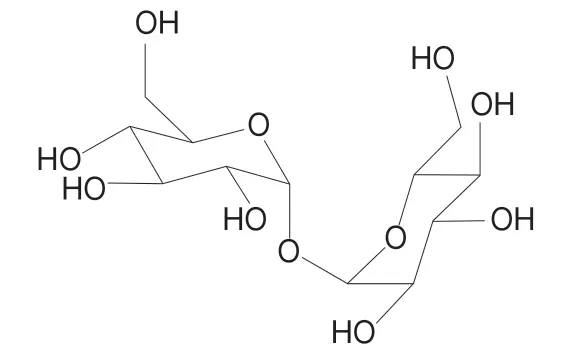
图1 海藻糖的结构式Fig. 1 Formula of trehalose
海藻糖的化学性质稳定,在低酸和高温等环境下的分解率仅为1%,能在高温、寒冷、高渗透压和失水干燥的环境中维持细胞生物膜的稳定性及蛋白质分子不变性,且海藻糖单体能稳定存在,仅能够被海藻糖酶分解。研究表明,许多动植物干燥失水后,由于体内含有海藻糖,其生物活性能够基本保留,复水后机体生物功能便可恢复正常[4]。大量学者研究证明,海藻糖在 医药[5-6]、农业[7]及食品[8]等领域都能对生物和生物分子起到保护作用,但关于海藻糖对于乳酸菌的保护作用仍缺乏系统的分析。因此本文结合近年来国内外学者的科学研究,从海藻糖对生物分子的保护机理出发,对乳酸菌应对脱水胁迫、冷冻胁迫、高温胁迫及氧化胁迫等恶劣环境下的保护作用进行综述,分析海藻糖对乳酸菌保护作用的机理,为今后选择合适的乳酸菌保护剂、探索提高乳酸菌菌剂活性的新方法提供一些理论参考。
1 海藻糖对生物分子的保护机理
海藻糖是自然界中化学性质最不活泼和最稳定的糖之一,这是因为两分子葡萄糖通过α,α-1,1糖苷键连接,使得海藻糖结构稳定。Nagase等[9]通过模拟计算证实在海藻糖的几种同分异构体中,α,α-1,1型结构的稳定性最强。与非还原糖蔗糖相比,海藻糖不易分解,不与蛋白质和氨基酸等发生反应,同时对于生物体的保护作用也更为显著[10]。但海藻糖对于生物体的保护机理目前还没有统一的解释,研究者根据海藻糖对生物膜及蛋白质的保护作用提出了以下4 种假说。
1.1 “水替代”假说
生物体内的蛋白质、糖类和脂质等大分子物质能够维持分子结构和功能特性的稳定原因是依赖于水分子外壳对其的包围与保护,大分子物质在冷冻干燥过程中失去活性的原因之一就是脱水引起的构象变化。如在脱水过程中,维持蛋白质空间结构的各种相互作用力被破坏,从而引起蛋白质变性。多羟基化合物(如海藻糖和蔗糖)中含有的羟基能替代水分子,通过氢键与蛋白质分子结合,并在蛋白质表面形成一层水化膜,抑制上述变化的发生,从而保持其蛋白质构象和分子活性稳定,这种作用被称为“水替代”假说[11]。上述提到的“替代”并非是完全替代,Pereira等[12]对磷脂膜在海藻糖存在与不存在的情况分别进行模拟,发现海藻糖通过氢键直接与磷脂膜相互作用,但磷脂双分子层表面的水分子并没有完全被海藻糖取代,只有1/5~1/4的氢键位点上的水分子被海藻糖取代。但海藻糖分子与磷脂膜头部基团之间的氢键仍然与保护效果密切相关。Crowe等[13]认为,当蛋白质的结构水被除去时,海藻糖能够通过羟基在干燥生物分子的蛋白质表面形成氢键,取代由水分子形成的氢键,使得生物分子在干燥缺水的条件下仍然能维持其原有结构及生物活性。
1.2 “玻璃态”假说
大部分糖类在水分含量较低时会形成非晶态结构。对于非晶态物质,随着温度的变化会呈现玻璃态、高弹态(橡胶态)和黏流态。其中处于“玻璃态”时,分子的运动能量较低,分子构象不易发生改变,分子性质最稳定,因此在乳酸菌冷冻、干燥及贮藏过程中,应当使其尽可能保持玻璃态[14]。玻璃态转变是非晶态聚合物的固有性质,玻璃态转变温度是指由玻璃态转变为高弹态或由高弹态转变为玻璃态所对应的温度。
由表1可知,与蔗糖和乳糖等糖类相比,海藻糖是玻璃态转变温度最高的二糖。干燥状态下,海藻糖紧密包裹住相邻的分子,被包围的分子难以实现构象上的转变,特别是当环境温度远远小于玻璃态转变温度时,能维持一定的空间结构,进而对分子起到保护作用[16]。此外,与蔗糖等不同的是,海藻糖的玻璃态不具有吸水性,而蔗糖的玻璃态吸水性很强,即使在温和、常见的环境湿度下,也很容易因吸水变得黏稠,被吸收的水分会溶解蔗糖表面,造成蔗糖玻璃态重结晶并呈现多孔结构[17]。这进一步证实海藻糖作为保护剂能维持乳酸菌制剂在运输及贮藏过程中的稳定性,避免因吸水造成乳酸菌损伤。

表1 不同糖类的玻璃态转变温度[15]Table 1 Glass transition temperature of different sugars[15]
1.3 优先排阻学说
优先排阻学说认为,海藻糖并非直接与不稳定的生物大分子(如蛋白质)直接作用,而是优先与其表面的水分子结合。研究表明,在含有蛋白质的溶液中加入海藻糖会使海藻糖被优先排斥分布于蛋白质周围,这使得蛋白质的疏水基团不被暴露于介质中,抑制了因空间结构改变引起的蛋白质变性[18]。同时,海藻糖的添加能够增强蛋白质的稳定性,提高蛋白质的变性温度。Lin Tiaoyin等[19]研究表明,在1 mol/L海藻糖存在的条件下,核糖核酸酶的变性温度提高10 ℃。此外该研究显示,加入海藻糖后蛋白质变性温度的提高与蛋白质-水界面表面张力的提高有关,变性温度提高10 ℃的同时,表面张力增加1.69 mN/m。Timasheff[20]研究表明,海藻糖在溶液中会与蛋白质等生物大分子表面的水分子优先结合,使得水分子从蛋白质的溶剂化层中被排除出来,这会导致蛋白质等生物大分子的溶剂化层半径和表观体积缩小,使分子之间的结构变得更加紧密、稳定,从而抵御外界恶劣环境的影响,更好地保护生物大分子。另外有研究表明,海藻糖与其他糖类相比具有更大的水化体积,至少是蔗糖和麦芽糖等的2.5 倍,正是由于这种特性,海藻糖可以取代溶液中更多的水分子,比其他糖类物质具有更强的稳定效果[21]。
1.4 分子间协同作用
许多研究表明,海藻糖在发挥应激保护作用的同时也会与其他分子发生协同作用。张玉华等[22]研究表明,将海藻糖和透明质酸复配后的混合物对双歧杆菌冻干后的保护效果更好。一方面海藻糖进入细胞内部与细胞膜磷脂和菌体蛋白极性基团结合,维持磷脂的“水化”状态和蛋白质二级结构的稳定性;另一方面,透明质酸作为高分子化合物,与海藻糖复配后,提高了混合物的玻璃态转变温度,在双歧杆菌细胞周围形成稳定的玻璃态外壳,即海藻糖的“水替代”作用与透明质酸的“玻璃态”作用有机结合,增强了对菌株细胞的保护作用。乳酸脱氢酶是乳酸菌胞内的一种关键酶,Miller[23]研究发现,海藻糖能与一些盐离子发生协同作用,在冷冻、干燥和贮存过程中增强对乳酸脱氢酶的保护作用,有助于保护乳酸菌制剂的产酸发酵能力。
2 海藻糖对乳酸菌的保护作用
海藻糖作为一种高效保护剂,最早是在耐干旱的植物中发现的,许多体内含有海藻糖的生物体,即使干燥脱水后仍能维持其生物活性。但绝大多数乳酸菌内源海藻糖含量甚微,不足以起到对乳酸菌的保护作用,因此国内外学者常常通过内源重组合成海藻糖或外源添加海藻糖作为培养基碳源或作为保护剂来提高乳酸菌的抗环境胁迫能力[24-26]。
2.1 抗脱水胁迫
为了延长乳酸菌发酵剂的保藏期,降低运输和贮藏成本,一般采用冷冻干燥或喷雾干燥技术除去菌株中的大部分水分,使其能长期稳定地贮藏在环境温度中。水分子是生物体的重要组成成分,一般占细胞总量的70%。细菌能够进行正常生理活动的最低水分含量为33.33%,但研究表明,为了满足乳酸菌制剂长时间贮藏的需求,乳酸菌发酵剂中残余水分含量应低于5%,水分活度低于0.25[27]。
由于水分子有助于维持蛋白质、DNA和脂质的稳定,并对细胞结构的稳定性有重要影响,因此当细胞干燥至低水分含量时,许多细胞成分会受到影响。特别是磷脂双分子层被认为是细胞中最敏感的部分,当细胞膜表面的水分子被除去时,会导致磷脂膜头部基团堆积,增加磷脂分子碳氢键间范德华相互作用力,破坏磷脂双分子层的稳定性,引起细胞膜从液晶相转变为凝胶相,进而导致菌株死亡[28]。因此学者普遍认为,造成乳酸菌损伤甚至死亡的主要原因之一就是由脱水引起的细胞膜损伤[29]。
海藻糖作为一种多羟基化合物,当乳酸菌细胞脱水时,海藻糖的羟基可以替代水分子与细菌的细胞膜和蛋白质相互作用形成氢键,在脱水过程中维持乳酸菌细胞结构及功能的稳定性,保证菌株细胞功能的正常 运行[30]。在脱水过程中,脂质膜经历了由液晶相到凝胶相的转变,在复水过程中,反向的转变过程会引起脂质膜的外泄。海藻糖的添加能够降低干膜的相转变温度,有效阻止这种脂质膜的相变及复水过程中伴随的细胞内容物外泄[31]。
公丕民[31]在研究喷雾干燥对保加利亚乳杆菌的损伤发现,当细胞内海藻糖含量达到3.47×10-7μg/CFU时,海藻糖便开始对喷雾干燥后的乳酸菌起到保护作用,当细胞内海藻糖含量达到10.09×10-7μg/CFU时,经喷雾干燥后的乳酸菌存活率接近100%,同时胞内负载海藻糖的乳酸菌经喷雾干燥后的菌粉产酸能力也得到改善。为了探究海藻糖的抗脱水能力,葛宇等[32]研究干燥过程对保加利亚乳杆菌菌体产酸能力和菌株存活率的影响,发现经海藻糖重悬的保加利亚乳杆菌经历了干燥脱水后,无论是产酸能力还是存活率均优于经其他类型保护剂保护的菌株,菌株的存活率高达83.67%,比蔗糖高 出近1 倍。
2.2 抗冷冻胁迫
在冷冻干燥过程中,冰晶形成引起的细胞膜破裂是造成乳酸菌死亡的主要原因。一般情况下,冰晶越大,细胞膜越容易破裂,越容易造成菌株死亡。反之,形成的冰晶越小,对细胞膜造成的机械损伤也就越小。在冻结过程中,冻结速度的快慢对菌株的存活率有重要影响。在慢速冷冻过程中,细胞外的自由水会率先形成冰晶,导致胞外溶质浓度升高,为了维持细胞内外的渗透压平衡,细胞内的水分外渗,细胞外形成较大的冰晶,对细菌造成损伤;当冻结速度过快时,胞内的水分还未外渗就被冻结成冰晶,致使细胞内形成的冰晶过大,进而对细胞造成损伤[33]。
在低温环境下,磷脂分子由于脂酰链相互靠近产生疏水力,使磷脂分子由排列疏松的液晶态转变为有序的刚性凝胶态,造成磷脂膜流动性下降[34]。同时研究发现,在冷冻干燥过程中,关键酶的失活也是造成乳酸菌死亡的重要原因;随着水分的冻结,细胞间隙中的溶质浓度逐渐升高,电解质浓度增加,高浓度的电解质会引起蛋白质空间结构的改变,造成蛋白质变性,进而影响酶活性[35]。
许多学者[36-37]发现,在冷冻干燥过程中添加海藻糖作为保护剂可以有效提高乳酸菌的冻干存活率。海藻糖可以显著抑制冰晶的形成,起到稳定细胞膜和蛋白质结构的作用。Sei等[38]分别对海藻糖溶液和水溶液中生长的冰晶进行观察,发现海藻糖溶液中的冰晶生长速率明显小于在纯水中的生长速率,当海藻糖溶液的浓度加倍后,冰晶的生长速率降低到原来的1/4;此外,海藻糖对冰晶的抑制作用大约是蔗糖的2 倍,说明在抑制冰晶生长方面,海藻糖比蔗糖更有效,认为二者之间的差别与海藻糖能比蔗糖结合更多的水分子有关。Gaber等[36]以蔗糖、海藻糖和脱脂乳等作为唾液乳杆菌的冻干保护剂,经冷冻干燥后发现,不添加保护剂的菌株冻干后的死亡率高达99%,当海藻糖单独使用或与其他保护剂复合使用时,唾液乳杆菌的冻干存活率显著提高。
2.3 抗高温胁迫
乳酸菌的耐热性较弱,通常当温度高于50 ℃就能引起乳酸菌失活,随着环境温度的上升,失活速率也会加快。采用喷雾干燥技术制备乳酸菌发酵剂时,出口温度一般为60~90 ℃[39],因此乳酸菌很容易受到热损伤,造成细胞壁、细胞膜、DNA和RNA等结构的损伤,以及蛋白质变性和酶失活。徐礼云等[40]采用8-苯胺-1-萘磺酸荧光探针法对细胞膜的流动性进行测定,发现当瑞士乳杆菌的生长温度由42 ℃升至47 ℃后,荧光强度升高,说明高温胁迫引起膜脂肪酸组成的变化,导致膜的流动性下降。研究[31]表明,细胞壁最容易受到温度影响,当温度超过35 ℃时就会引起细胞壁结构的变化,细胞膜大约在50 ℃时就会发生损伤,蛋白质的热变性温度为65 ℃。Teixeira等[41]探究不同温度条件下脱脂牛乳中保加利亚乳杆菌的生长情况时发现,当温度在64 ℃以下时会对细胞膜造成损伤,而当温度达到65 ℃以上时,就会引起核糖体和蛋白质的损伤。
Colao等[17]研究表明,海藻糖能够使蛋白质的变性温度升高,因此海藻糖在喷雾干燥过程中常常充当热保护剂,来维持细胞的热稳定性。王芳等[42]研究发现,经70 ℃处理后的乳酸杆菌存活率随处理时间的延长而下降,而添加5%海藻糖后,乳酸杆菌经70 ℃处理3 min后的活菌数是对照组的2.4 倍,处理5 min后的活菌数与对照组相比更高达100 倍。说明海藻糖能够显著提高乳酸杆菌的耐热性,提高蛋白质的变性温度,保护蛋白质空间结构不受损伤,进而提高高温干燥过程中菌株的存活率。Mahmud等[43]将菌株培养温度从30.0 ℃逐渐升高至38.0、40.0、41.5 ℃,检测海藻糖重组菌株对热应激的反应,结果显示,热胁迫导致所有菌株中的海藻糖含量增加,表明海藻糖的积累对于菌株在热胁迫下的高生长活性十分重要。
2.4 抗氧化胁迫
氧化损伤也是造成乳酸菌细胞死亡的一个重要因素。一方面,氧化损伤来源于环境中的氧气浓度过高,在高氧环境下,乳酸菌会产生大量的自由基并在细胞内积聚,这些自由基会与乳酸菌细胞内的氨基酸、脂质和核酸等发生氧化应激反应,造成乳酸菌细胞损伤。Ghandi等[44]分别采用空气、氮气或添加抗氧化剂作为雾化介质的方式观察喷雾干燥后的乳酸乳球菌存活率,发现当使用氮气雾化、添加抗氧化剂或二者同时使用时乳酸乳球菌的存活率明显提高。另一方面,乳酸菌细胞膜脂肪酸在贮藏过程中发生氧化也是细胞氧化损伤的重要来源。由于不饱和脂肪酸亚甲基上的氢原子较为活泼,因此易被自由基夺走形成脂质自由基,与氧气结合后形成脂质过氧自由基,导致细胞膜不饱和脂肪酸含量减少,细胞膜流动性下降,影响细胞膜结构和正常的生理功能[33]。
海藻糖具有保护细胞免受氧自由基侵害的作用[45]。在培养基中添加海藻糖会增强乳酸菌对H2O2的抗性,高浓度的海藻糖会抑制自由基对蛋白质氨基酸的破坏。 陈喜玲等[24]将大肠杆菌中的海藻糖基因(OtsB)导入缺失海藻糖基因的副干酪乳杆菌中,发现在H2O2胁迫下,重组菌株的存活率明显提高,表现出良好的抗氧化胁迫性,胁迫10 min后菌株存活率提高1 倍,胁迫15 min后菌株存活率提高3 倍。此外,海藻糖对降低干燥过程中的脂质氧化也起到重要作用。Nery等[46]研究表明,1 分子海藻糖能够与不饱和脂肪酸的1 个双键相互作用形成复合物,从而降低脂质氧化水平。
3 结 语
研究海藻糖对乳酸菌的保护作用,不仅有助于了解乳酸菌在不同环境胁迫下的损伤情况和海藻糖的作用机制,并且对今后探索更加经济、高效的乳酸菌保护剂提供了理论参考。目前关于海藻糖对生物分子的保护机理共形成了4 种假说:“水替代”假说、“玻璃态”假说、优先排阻学说和分子间协同作用学说。但海藻糖对于细胞的作用机理仍然存在很大争议,单一的作用机理似乎难以解释海藻糖的保护作用。
国内外学者从乳酸菌抗脱水胁迫、抗冷冻胁迫、抗高温胁迫和抗氧化胁迫等方面进行了研究,以探究海藻糖在抵御不同环境胁迫对乳酸菌造成损伤的保护作用。海藻糖保护乳酸菌免受氧化胁迫的研究中,多以内源导入海藻糖基因为主,添加外源海藻糖能否发挥抵御氧化胁迫的作用仍然值得探索。
近年来,海藻糖作为非特异性保护剂,越来越成为国内外学者研究的热点,其自身的优良特性及其在保护菌株免受恶劣环境胁迫方面的突出效果必将吸引更多关注。随着研究的不断深入,海藻糖作为菌株保护剂必将更加广泛应用于菌株发酵、干燥及运输贮存的过程中,也将会给社会各行业带来巨大的经济效益。
