胆汁酸受体与猪骨骼肌发育相关基因的关联分析
2021-12-17孙丽娟赵伟杰谭伟豪李永祥陈履双王丽娜
孙丽娟,罗 培,赵伟杰,谭伟豪,李永祥,陈履双,王丽娜*
(1.华南农业大学动物科学学院,广东广州 510000;2.广东省动物营养调控重点实验室 广东广州 510000)
骨骼肌作为人类和动物躯体的重要组成部分,约占人类和动物体总体重的40%[1]。骨骼肌的生长发育过程受到多种生长因子和遗传环境因素的共同影响,有关骨骼肌发育的分子机制尚不完全明了,与其相关的功能基因一直是研究热点。其中,生肌调节因子(Muscle Regulatory Factors,MRFs)基因家族控制着肌肉的发育,包括从前体肌细胞的定型、增殖及肌纤维的形成,直到个体出生后肌肉的成熟和功能的完善性整个过程[2]。已有研究发现,该家族编码4 种肌肉特异性转录因子,分别是生肌决定因子(Myogenic Determining Factor,MyoD)、肌细胞生成素(Myogenin,MyoG)、生肌因子5(Myogenic Factor 5,Myf5)和生肌因子6(Myo genic Factor 6,Myf6),它们是控制骨骼肌生成的关键调节因子,共同控制肌肉的生成[3]。MyoD和Myf5基因在成肌细胞增殖过程中表达,主要参与生肌过程,MyoG与Myf6主要负责肌肉分化[4]。MyoG在分化末期表达,Myf6基因主要在出生后表达,通过调控收缩蛋白和调节蛋白同工型在不同时期的表达使肌肉细胞分化为成熟的肌纤维。
成熟后的肌纤维根据骨骼肌肌纤维(MyHC)同工型表达模式所分出的4 种类型(I 型、IIa 型、IIb 型与IIx 型)表现出不同的生理功能与代谢特征,其中I 型和IIa 型肌纤维表现出氧化代谢特性,而IIb 型和IIx 型肌纤维表现出糖酵解特性[5]。不同类型的肌肉由于肌纤维类型的差异也往往会呈现出体积上的差异,I 型肌纤维直径较细,而II 型直径较粗。
然而,近年来的研究发现骨骼肌中也有胆汁酸受体的表达。胆汁酸受体主要包括法尼酯X 受体(Farnesol-X-Receptor,FXR)和G 蛋白偶联受体TGR5(the G-protein Coupled Receptor 5,GPBAR1)[6]。FXR 作为第一个被发现的胆汁酸天然内源性配体,其在体内糖脂代谢稳态中的作用已经被多项研究所验证。FXR 激动剂Fexaramine 可显著改善血糖水平,并减少饮食诱导的体重增加[7]。同时,FXR 激活后还可以通过调控肝脏脂肪代谢保护骨骼肌免于出现脂毒性的症状[8]。有研究发现,全身性基因敲除FXR 的小鼠和肠道缺乏FXR 的小鼠口服葡萄糖后,耐受量可显著改善且体重降低[9-10]。相关试验证明,FXR 可以介导CDCA(鹅脱氧胆酸)促进小鼠胚胎肝细胞的增殖和分化[11]。据相关研究报道,胆汁酸通过与肝细胞中的FXR 结合,发挥类固醇激素的作用,从而反馈调节自身的合成[12],在小肠中,FXR 被高水平胆汁酸激活后诱导FGF19(或者小鼠FGF15)的转录和分泌,FGF15/19 通过门静脉进入肝细胞,激活FGF 受体4(FGFR4)/β-Klotho 的复合物,形成的复合物能够抑制CYP7A1和CYP8B1基因的转录[13],且研究表明,给小鼠肌肉注射FGF19 可以诱导其骨骼肌肥大,调节骨骼肌质量并改善小鼠的肌肉消耗[14]。TGR5 作为研究最为深入的膜结合G 蛋白偶联受体,其在体内多个组织中均有表达,胆汁酸是目前已知的TGR5 的唯一内源性配体[15]。有研究发现,从TGR5-/-雄性小鼠肌肉中提取的肌管和分离出的肌纤维会产生萎缩状态,当用胆酸(CA)和脱氧胆酸(DCA)处理分离出的肌纤维培养物后,会增加其肌纤维直径和肌球蛋白重链(MyHC)蛋白表达水平以及降低Atrogin-1 和MuRF-1 等萎缩蛋白的表达水平[16]。Sasaki 等[17]发现TGR5 在骨骼肌中的功能主要是调节肌细胞分化,改善由肌肉萎缩所产生的肌肉重量降低。除此之外,有研究还发现胆汁酸通过TGR5 诱导Dio-2 表达,使T4 转化为T3 并且降低骨骼肌中的胰岛素抵抗水平[18-19]。
目前,大多数研究主要是关于胆汁酸受体在肝脏和肠道中的作用,而其在骨骼肌发育过程中的调控作用及机制研究还很少。有研究发现,肌肉特异性基因敲除TGR5的小鼠会表现出较低的肌肉质量和强度,同时也会降低骨骼肌中与肥大和分化相关基因的表达水平,而TGR5 激活则可增强肌细胞分化和肌肉肥大[17]。可见,胆汁酸受体很可能与骨骼肌生长发育有密切联系。
莱芜猪是我国优良的土种猪,肉质鲜美。但是与外来品种相比有生长速度慢、产肉率低等缺点。为了探究胆汁酸受体是否参与了莱芜猪和杜洛克肌肉生长发育差异的形成,本研究采用Western Blot、qRT-PCR 和ATP酶染色等技术检测了2 个品种断奶仔猪背最长肌和腰大肌中胆汁酸受体和肌肉发育相关基因的表达,然后进行相关性分析,从而探究它们之间的关联性,为进一步研究胆汁酸受体在骨骼肌发育过程中的作用机理奠定重要基础。
1 材料与方法
1.1 实验动物 选取12 头同一胎次、健康状况良好的21 日龄杜洛克(江西农业大学种猪场)和莱芜断奶仔猪(来自山东农业大学种猪场),公母各6 头,采集背最长肌、腰大肌和血液样本。所有组织样品采集后,经液氮速冻,保存于-80℃备用。
1.2 主要仪器 多功能酶标仪,美国Gene 有限公司;电泳仪,美国Bio-Rad 公司;垂直电泳槽,美国Bio-Rad 公司;蛋白印记转移槽,美国Bio-Rad 公司;凝胶成像系统,美国UVP 公司;实时荧光定量PCR 仪,美国Life Technologies 公司;冰冻切片机,德国Leica 公司。
1.3 主要试剂β-actin 抗体、Tubulin 抗体、山羊抗兔二抗、山羊抗鼠二抗,美国Bioworld Technology 公司;FXR 抗体,美国Abcam 公司;TGR5 抗体,美国Novus 公司;MyoD 抗体,正能生物科技有限公司;MyoG 抗体,美国Abcam 公司;RNA 快速提取试剂盒,广州美基生物科技有限公司;BCA 蛋白浓度测定试剂盒,美国Thermo Fisher Scientific 公司;Western Blot快速制胶试剂盒,美国Bio-Rad 公司;琥珀酸脱氢钠(SDH),北京索莱宝科技有限公司;腺苷三磷酸二钠盐(ATP),北京索莱宝科技有限公司。
1.4 实验方法
1.4.1 蛋白质免疫印迹(Western Blot)检测 取肌肉组织提取总蛋白,经样品蛋白质浓度测定(BCA 法)、样品测定、蛋白质变性、上样、电泳、转膜、封闭、孵育抗体、显影、定影后统计结果。
1.4.2 实时荧光定量PCR 测定基因mRNA 相对表达量将肌肉样品取出40~50 mg,提取总RNA,具体操作按Trizol 试剂盒说明书进行,用RNA 浓度仪(Thermo Nano Drop Lite)检测RNA 浓度。用反转录试剂盒(Vazyme 公司,R323-01,20 μL 反应体系)进行反转录,具体操作按说明书指示进行。按荧光定量试剂盒(Vazyme 公司,Q111-02,20 μL 反应体系)具体操作按说明书指示进行。扩增条件为:95℃预热10 min,循环1 次;(95℃,15 s;55~62℃,15 s;72℃,40 s),循环40 次,(95℃,60 s;55~62℃,30 s;95℃,30 s),循环1 次。每个样品作6 个重复,以β-actin作为内参基因,2-ΔΔCt法计算目的基因mRNA 相对表达量。引物序列由NCBI 数据库获得,使用Primer 6.0 软件设计相应特异性引物,引物由生工生物工程(上海)股份有限公司合成(表1)。
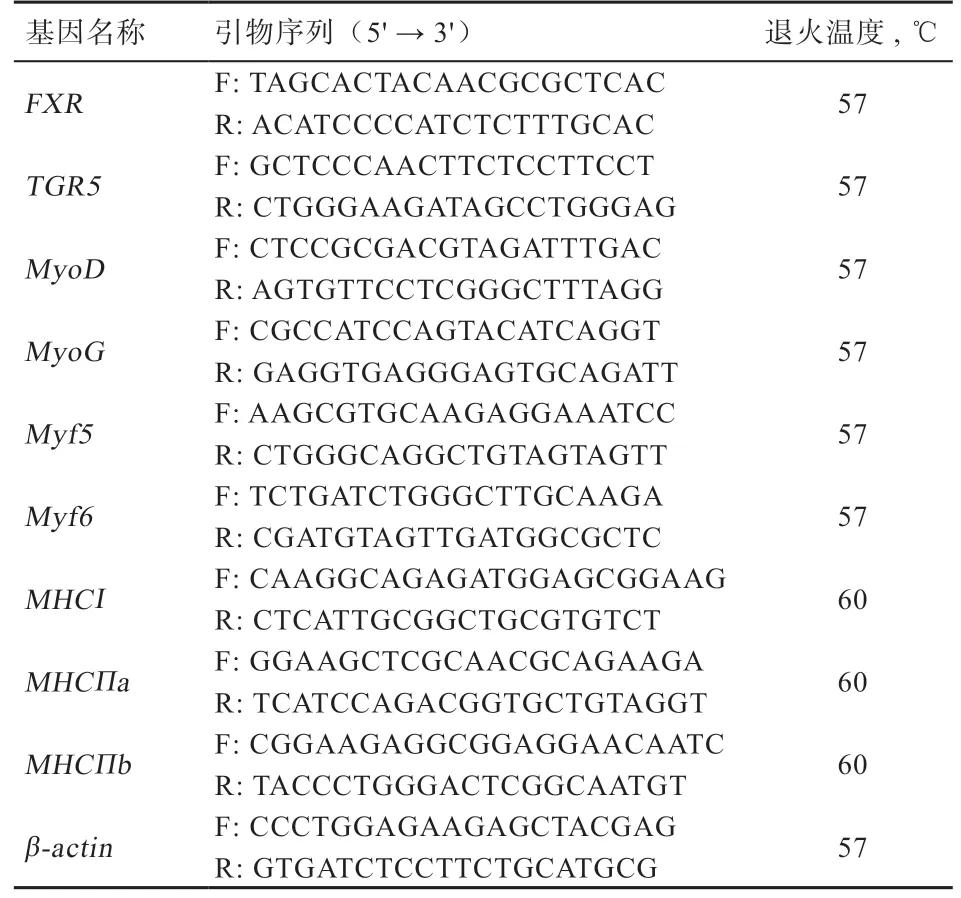
表1 实时荧光定量PCR 反应引物序列
1.4.3 ATP 酶酸染测定肌纤维类型 取新鲜肌肉组织放置到提前降温至-24℃的冰冻切片机中平衡30 min 后,经修剪、包埋、切片、染色、封片后,使用显微摄像系统进行切片观察和图像采集,采用Image-Pro Plus 6.0图像分析软件进行计数。
1.5 数据统计分析 实验数据使用GraphPad Prism 8.0统计软件进行统计及差异分析。采用t-test 检测比较两组间的统计学差异,所有数据均以平均值± 标准误(mean±S.E)表示,以P<0.05 表示差异显著,P<0.01表示差异极显著,P>0.05 表示差异不显著。
2 结果
2.1 杜洛克与莱芜断奶仔猪血清胆汁酸含量差异 本实验分别检测了杜洛克和莱芜猪断奶仔猪血清中的总胆汁酸含量和各种单体胆汁酸含量。结果显示,莱芜猪与杜洛克断奶仔猪血清总胆汁酸含量无差异(P>0.05)(图1A、C)。莱芜猪雌性断奶仔猪血清中石胆酸(LCA)含量低于杜洛克(P<0.05)(图1D)。
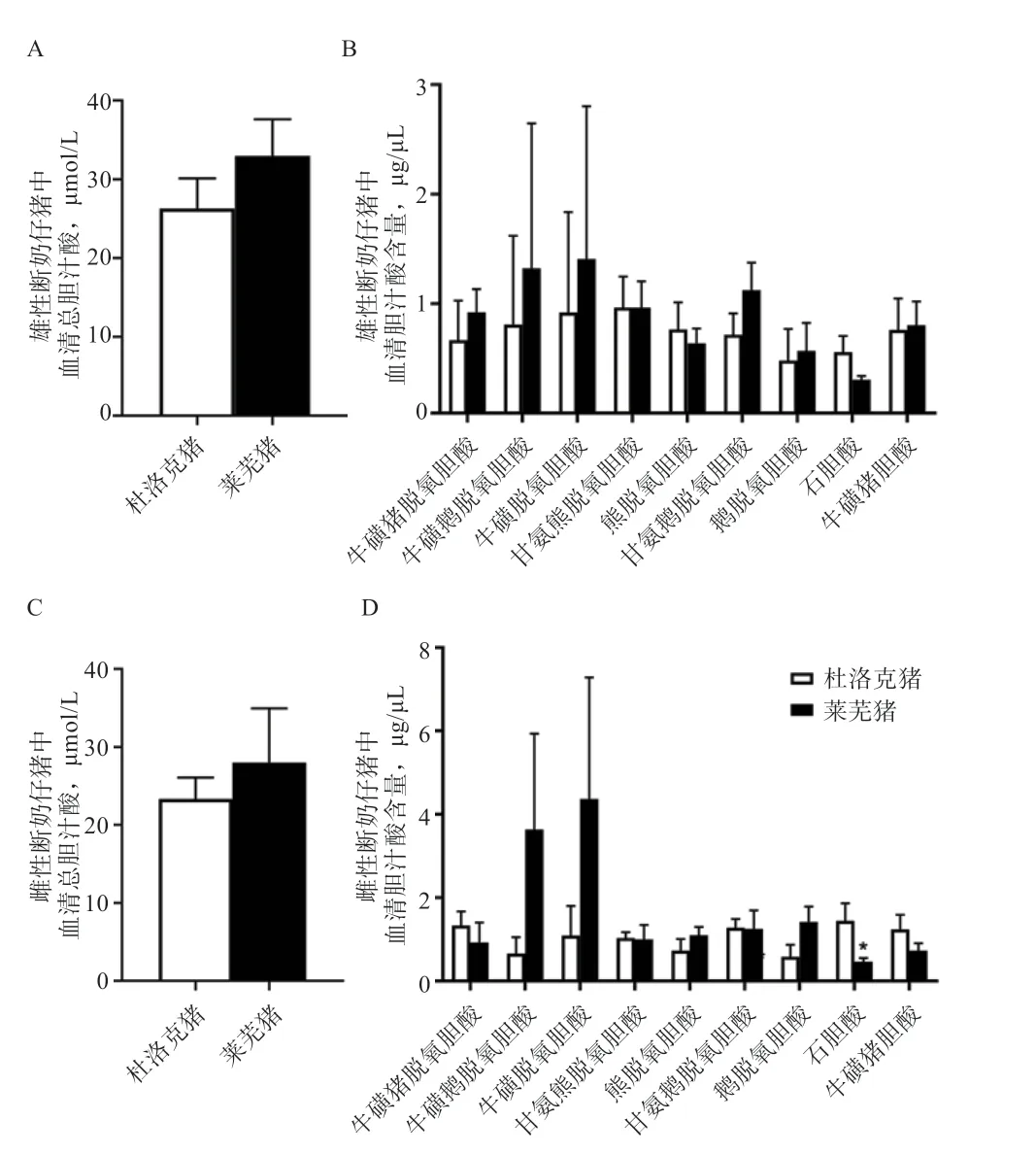
图1 杜洛克与莱芜猪断奶仔猪血清胆汁酸水平
2.2 杜洛克与莱芜断奶仔猪骨骼肌胆汁酸受体表达水平由图2 所示,在雄性断奶仔猪,杜洛克与莱芜猪背最长肌和腰大肌中FXR 和TGR5 的表达均无差异(P>0.05)(图2B、C、H、I);在雌性断奶仔猪,莱芜猪背最长肌中TGR5 蛋白水平和TGR5mRNA 水平低于杜洛克(P<0.05),FXR 蛋白水平有低于杜洛克的趋势(P=0.06),而FXRmRNA 水平低于杜洛克(P<0.01)(图2E、F),腰大肌中FXR的mRNA 水平高于杜洛克(P<0.05)(图2L)。
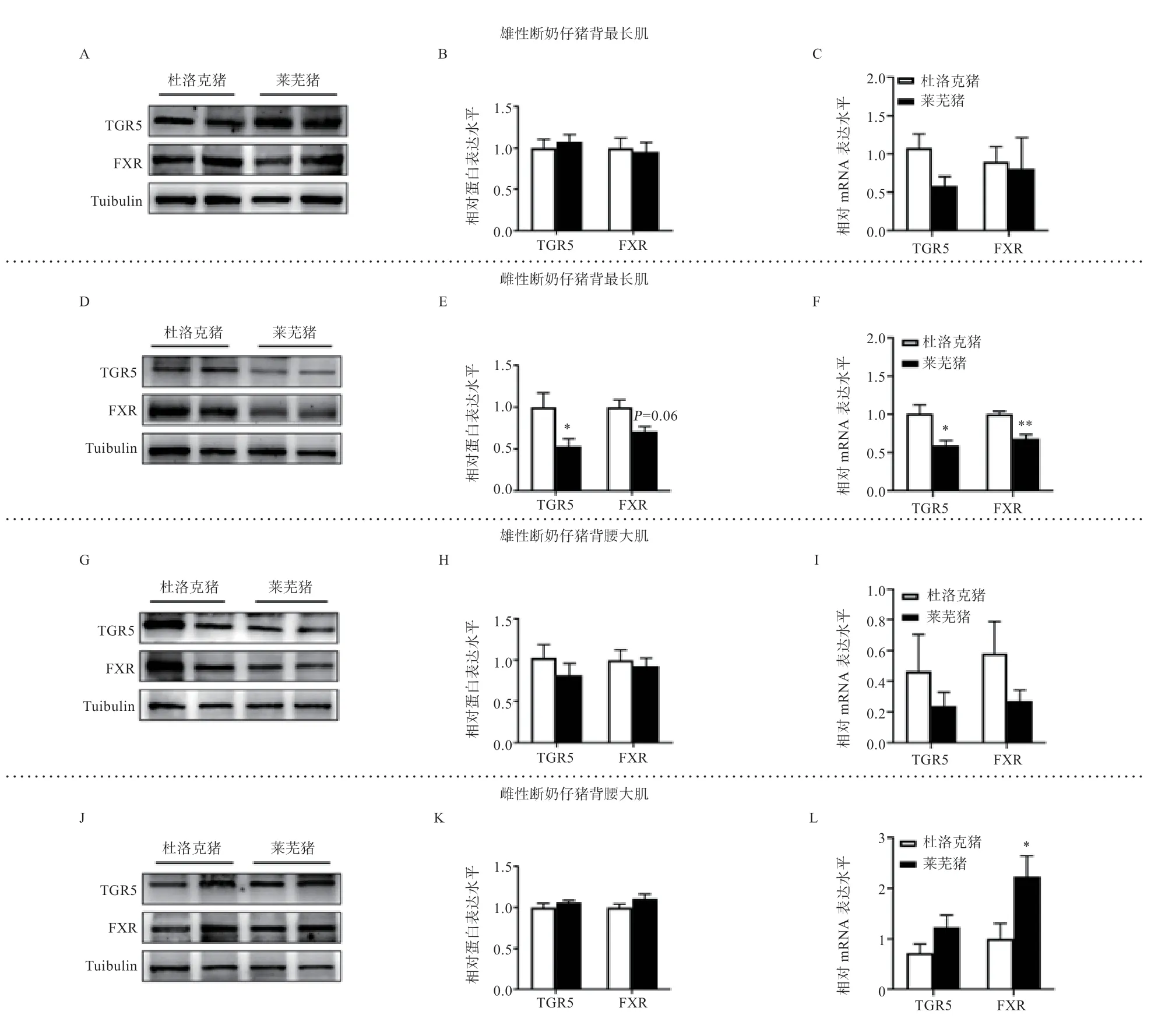
图2 杜洛克与莱芜猪断奶仔猪肌肉中胆汁酸受体表达
2.3 杜洛克与莱芜断奶仔猪骨骼肌肌纤维分型 如图3所示,在雄性断奶仔猪,莱芜与杜洛克背最长肌和腰大肌中3 种肌纤维类型的mRNA 水平均无差异(P>0.05)(图3A、B);在雌性断奶仔猪,莱芜猪腰大肌中MHCΠb的mRNA 水平低于杜洛克(P<0.05)(图3D),而莱芜猪与杜洛克背最长肌中3 种肌纤维类型的mRNA水平均无差异(P>0.05)(图3C)。
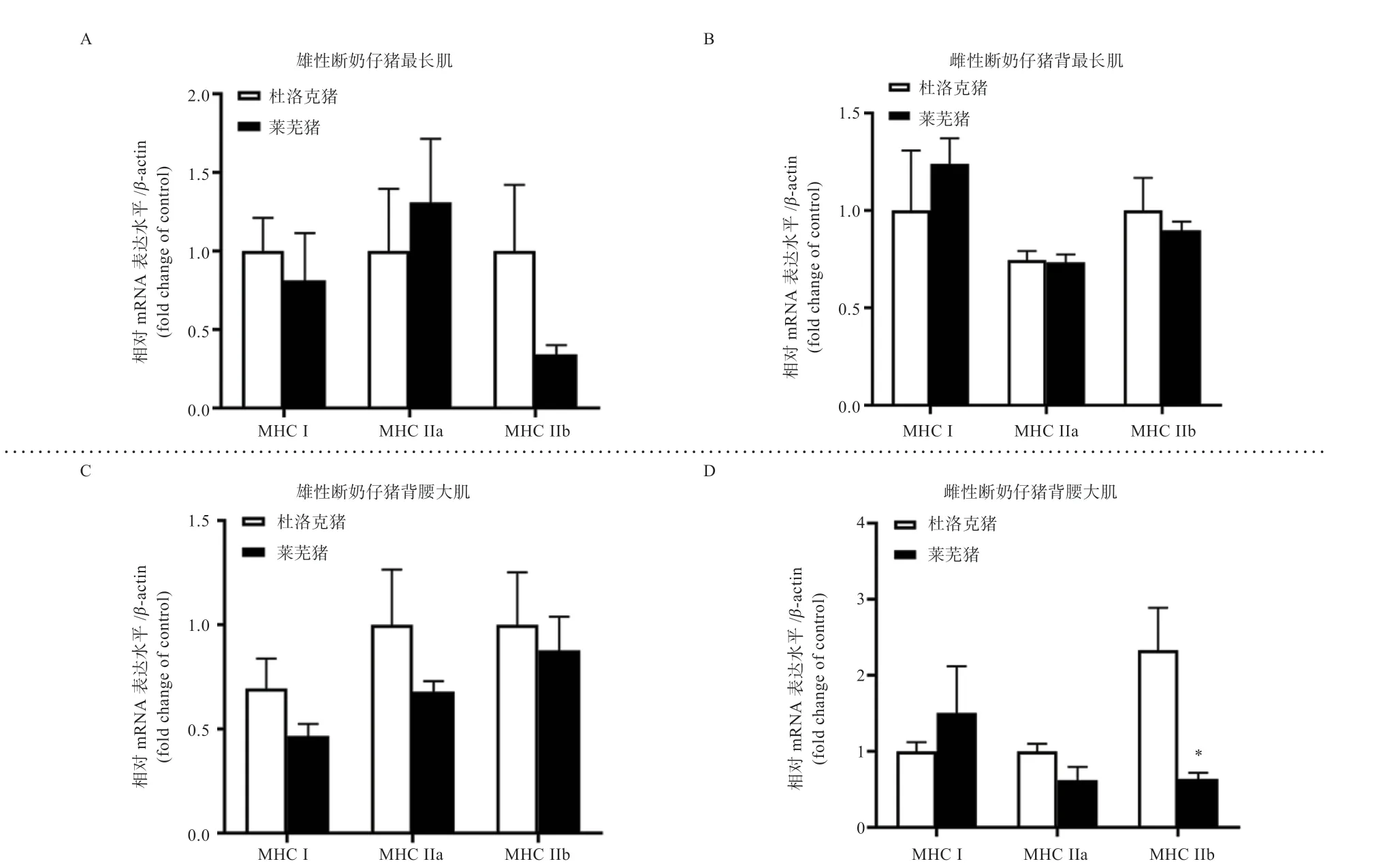
图3 杜洛克与莱芜断奶仔猪肌肉中肌球蛋白重链表达
在检测了3 种肌球蛋白重链的mRNA 水平基础上,本研究进一步采用ATP 酶染色进行验证。结果显示,雄性断奶仔猪中莱芜猪背最长肌中MHCI 的分布比例有高于杜洛克的趋势(P=0.088),而MHCIIb 的分布比例有低于杜洛克的趋势(P=0.061)(图4C);腰大肌中MHCI 的分布比例高于杜洛克(P<0.01),而MHCIIb 的分布比例低于杜洛克(P<0.01)(图4D);雌性断奶仔猪中杜洛克与莱芜猪腰大肌中3 种肌纤维类型的分布比例无差异(P>0.05)(图4E)。

图4 杜洛克与莱芜断奶仔猪肌肉中肌纤维类型
2.4 杜洛克与莱芜断奶仔猪骨骼肌生肌因子的表达 如图5 所示,在雄性断奶仔猪,莱芜背最长肌中MyoG的mRNA 水平低于杜洛克(P<0.05)(图5C),腰大肌中生肌因子的表达水平均无差异(P>0.05);雌性断奶仔猪中莱芜猪背最长肌中MyoD和MyoG的mRNA 水平低于杜洛克(P<0.05),而Myf6的mRNA 水平高于杜洛克(P<0.001)(图5F),腰大肌中Myf5和Myf6的mRNA 表达水平高于杜洛克(P<0.05)(图5L)。

图5 杜洛克与莱芜断奶仔猪肌肉中生肌因子
2.5 杜洛克与莱芜断奶仔猪骨骼肌胆汁酸受体和肌肉发育相关基因的关联分析 本研究发现,莱芜和杜洛克断奶仔猪肌纤维类型存在差异,并且跟性别有关。肌肉发育相关基因的差异主要出现在2 个品种的雌性断奶仔猪中,同时血清中LCA 水平和骨骼肌中胆汁酸受体的表达水平均在2 个品种的雌性断奶仔猪中出现差异(P<0.05)。基于此,针对雌性断奶仔猪骨骼肌中胆汁酸受体表达水平和生肌因子的表达情况进行了进一步的相关性分析。
结果发现,杜洛克猪骨骼肌中,TGR5在背最长肌中的表达与Myf5正相关(P<0.05),而在腰大肌中TGR5的表达与MyoG和Myf6正相关(P<0.05)(图6A、B)。然 而TGR5与Myf5、MyoG以 及Myf6在莱芜猪中无相关性(P>0.05)(图6C、D)。另一方面,FXR在杜洛克和莱芜猪的背最长肌和腰大肌中的表达跟4 个生肌因子的表达也出现相关性,但并无明显规律(图6A、B、C、D)。

图6 杜洛克与莱芜雌性断奶仔猪肌肉中胆汁酸受体和生肌因子的相关性
基于上述结果,快速生长的瘦肉型猪杜洛克猪肌肉中生肌因子的表达与TGR5 的表达存在显著相关性,而在莱芜猪似乎丧失了这种相关性。因此,推测胆汁酸受体TGR5 在莱芜猪和杜洛克猪肌肉生长发育特性差异形成过程中可能扮演了一定的角色。
3 讨 论
近年来的研究已经证实,胆汁酸作为一个重要的信号调节分子,能够激活胆汁酸受体并参与机体胆汁酸代谢、糖脂代谢、能量代谢以及机体的免疫调节等过程,具有广泛的生物学作用[20-21]。
本研究发现不同品种猪血清中胆汁酸组成存在差异,莱芜雌性断奶仔猪血清中LCA 含量显著低于杜洛克。与此同时,骨骼肌中胆汁酸受体的表达存在明显的品种和性别差异,TGR5 在莱芜雌性断奶仔猪背最长肌中的表达水平显著低于杜洛克,且背最长肌中FXR的mRNA 表达水平极显著低于杜洛克,而这些基因表达差异在雄性断奶仔猪骨骼肌中无显著性。很多研究表明,血清胆汁酸与各组织胆汁酸受体的表达存在相互调控的作用。Vassileva 等[22]认为胆囊膜上的TGR5 激活后通过介导尖端钠依赖性胆盐运输体(ASBT)促进胞外胆汁酸逆转运,使血清的胆汁酸浓度升高,反馈性地抑制了肝细胞中的CYP7A1,从而使胆汁酸合成减少,而血清及胆汁中胆固醇增多,故易形成胆结石。而Sinal等[23]已经证实FXR-/-小鼠能够诱导肝脏胆汁酸水平升高和肝脏损伤,进而引起肝脏脂肪变性、炎症和纤维化,有研究表明,在胆道阻塞时,胆汁酸浓度的上升会刺激FXR 的表达上调,并随阻塞时间延长,FXR 蛋白表达增高相对缓慢,同时FXR 的激动剂——CDCA 可以减轻胆道阻塞后大鼠肝脏损伤[24]。Kobayashi 等[25]对55 位中年的非酒精性脂肪肝(NAFLD)患者进行血清胆汁酸及代谢指标测定,通过Spearman 试验显示,血清中CDCA 与下肢骨骼肌体积(SMV)呈正相关,CDCA 含量增加引起SMV 水平升高。除此之外,有研究报道血清胆汁酸对减重手术的代谢有促进作用,并且通过激活膜受体TGR5,胆汁酸能够增加能量消耗以及改善糖脂代谢[26]。由此可见,胆汁酸与胆汁酸受体之间存在同向调控。本研究中也发现莱芜雌性断奶仔猪血清中LCA 含量降低可能会导致肌肉中TGR5 和FXR 的表达水平降低。
肌肉发育相关基因的研究结果显示,在雌性断奶仔猪,只有生肌因子的表达水平出现显著差异,而肌纤维类型无差异。推测胆汁酸主要通过其受体影响生肌因子的表达以及肌纤维的发育。背最长肌中MyoD的mRNA 表达显著低于杜洛克,而背最长肌和腰大肌中Myf6的mRNA 表达显著高于杜洛克。近年来关于骨骼肌发育的相关功能基因的研究一直是热点,猪骨骼肌中肌肉发育基因的表达在决定肌肉生长发育和中间差异中起着重要作用[27]。Yoichiro 等[28]发现梅山猪在其妊娠第75 天及出生时骨骼肌肌纤维的总量远少于西方瘦肉型猪种——大白猪,而这种差异直接导致大白猪出生后肌肉发育能力远高于梅山猪。Zhonglin 等[29]通过基因表达长序列分析(LongSAGE)技术,获得了通城猪和长白猪性交后33、65 d 和90 d 的骨骼肌转录组图谱,发现长白猪的肌肉生长因子和蛋白质调控相关基因表达上调,同时定量分析结果显示,这些基因在通城猪肌肉的表达下调,且其肌肉生长速度较为缓慢。推测这些差异形成可能和肌肉中胆汁酸受体表达差异有关。
本研究发现,肌肉发育相关基因的表达差异主要出现在两品种的雌性断奶仔猪中,并且血清中LCA 水平和骨骼肌中胆汁酸受体表达水平均在两品种的雌性断奶仔猪中出现显著差异。相关性分析结果显示,快速生长的瘦肉型猪杜洛克猪肌肉中生肌因子的表达与TGR5的表达存在显著相关性,而在莱芜猪肌肉中似乎丧失了这种相关性。已有研究证明,循环总胆汁酸与骨骼肌体积有关联,12-羟基甾醇胆汁酸(包括脱氧胆酸)与骨骼肌体积减小有关[30],同时有关试验已经验证了在肉鸡日粮中添加60 mg/kg 和80 mg/kg 的胆汁酸可以显著提高腿肌重量[31],而在妊娠母鼠日粮中添加植物甾醇酯(PE)可以影响母胎间胆汁酸转运及脂质代谢,并且可能通过胆汁酸受体的作用促进子代肌肉发育[32]。据有关研究发现,TGR5KO 小鼠会表现出较低的肌肉质量和强度,并且会降低骨骼肌中与肌肉肥大和肌肉发育相关的基因的表达,反之,激活TGR5 可增强肌细胞分化和促进肌肉肥大[17]。这提示肌肉中胆汁酸受体可能与肌肉发育相关基因有关联性,而该推测有待后期试验的进一步验证。
4 结 论
综上所述,骨骼肌中的TGR5 可能促进肌肉的生长发育,胆汁酸受体调节网络的差异可能参与了不同品种猪肌肉生长发育表型差异的形成。
