利用分子标记辅助选育抗稻瘿蚊水稻新品系
2021-12-14张宗琼夏秀忠杨行海张晓丽徐志健李丹婷农保选
郭 辉,陈 灿,张宗琼,夏秀忠,杨行海,张晓丽,徐志健,李丹婷,农保选,冯 锐
(广西农业科学院水稻研究所/广西水稻遗传育种重点实验室,广西 南宁 530007)
【研究意义】亚洲稻瘿蚊(Orseoliaoryzae)是南亚、东南亚和我国南方稻区的主要灾害性害虫之一,以山区稻区发生较重。该虫主要以幼虫蛀食水稻生长点,使心叶停止生长,基部膨大,形成“标葱”,危害严重时水稻不能抽穗结实。我国稻瘿蚊年发生面积约98万hm2,损失稻谷约10亿kg[1-2]。稻瘿蚊为钻蛀性害虫,化学防治效果不理想,而挖掘抗虫基因、培育推广抗虫品种是防治稻瘿蚊最经济安全的手段[3]。分子标记辅助选择(Molecular marker-assisted selection,MAS)是通过分析与目标性状紧密连锁的分子标记来完成对目标性状的选择,可有效缩短育种周期和提高育种效率,已在水稻、玉米和小麦等粮食作物及多种经济作物上广泛应用[4-6]。在传统的稻瘿蚊抗性育种中,抗性鉴定易受虫源、时间和环境的影响,耗时费力。因此,通过分子标记辅助选择结合抗性鉴定,可缩短育种时间,减少抗虫鉴定工作量,提高育种效率,对水稻稻瘿蚊抗虫育种具有重要意义。【前人研究进展】自20世纪80年代以来,各国学者已报道了12个稻瘿蚊抗性基因[7-8],且除Gm9和Gm10基因外均已确定其在染色体上的位置。其中,Gm1基因是从材料W1263中鉴定到的显性抗稻瘿蚊基因,并定位到第9号染色体在标记RM23941与RM23956之间[9-10];Gm2、gm3、Gm6和Gm7基因成簇分布于第4号染色体长臂31.02~31.84 Mb之间0.82 Mb范围[10-14],并已确定隐性基因gm3的候选基因为LOC_Os04g52970[15];Gm4和Gm8基因都定位到第8号染色体,其中Gm4基因在Abhaya中发现,位于标记RM22551与RM22562之间0.33 Mb范围,并确定LOC_Os08g09670为其候选基因[16-17];材料Jhitpiti和Aganni都携带有Gm8基因,定位到标记RM22685与RM22709之间430 kb范围,并最终确定LOC_Os08g15080为其候选基因[18-19];材料ARC5984携带有Gm5基因,最新定位到第12染色体在标记Z57~Z64之间49 kb范围,并筛选到2个候选基因LOC_Os12g36830和LOC_Os12g36880[20-21]。从CR57-MR1523中鉴定到Gm11(t)基因,同样定位在第12号染色体,位于SSR标记RM28574和RM28706之间,分别相距4.4和3.8 cM[22]。gm12基因是材料MN62M携带的抗稻瘿蚊隐性基因,位于标记RM6800与S2_411840之间100 kb范围[8]。【本研究切入点】在已报道的抗稻瘿蚊基因中,仅Gm6基因来源于我国,也是目前我国唯一在生产上推广应用的抗稻瘿蚊基因。为防范抗性基因单一水稻品种大面积种植后出现抗性丧失现象,有必要进一步拓宽我国水稻抗稻瘿蚊基因的材料来源。【拟解决的关键问题】本研究课题组前期从广西稻种资源中筛选到1份全抗我国4种稻瘿蚊生物型的材料GXM-002-1,经遗传分析与基因定位,将抗稻瘿蚊基因定位到第12号染色体分子标记12M-25与12M-29之间,与Gm5基因位置相同[20-21],说明与抗稻瘿蚊基因Gm5是同一基因或等位基因。利用该抗源通过杂交和回交,结合分子标记辅助选择将其抗性基因转入优质恢复系R553,选育出农艺性状优良的抗稻瘿蚊新品系,为培育抗稻瘿蚊新品系提供良好种质资源。
1 材料与方法
1.1 试验材料
植物材料:供体亲本为高抗稻瘿蚊材料GXM-002-1,含有抗稻瘿蚊基因Gm5,来自本课题组前期收集的广西稻种资源;受体亲本为水稻优质恢复系R553,为本单位优质杂交稻组选育提供。抗蚊对照为多抗1号(含Gm6基因),为广东省农业科学院选育品种,感蚊对照为台中1号(即TN1,不含抗蚊基因),由广西农业科学院水稻研究所稻种资源中心提供。
供试虫源:在广西大新和马山县等地稻瘿蚊发生危害时期,采集刚出现的乙型标葱,在广西农业科学院水稻研究所温室内繁殖分型后以感蚊品种TN1饲养备用,生物型经鉴别为中国Ⅳ型。
1.2 试验方法
1.2.1 选育流程 试验于2017—2020年在广西南宁市进行。以抗稻瘿蚊材料GXM-002-1为父本,优质恢复系R553为轮回亲本进行杂交,并进行2次回交后再自交;从BC1F1代开始对后代进行标记辅助选择,从分离群体中选出含有抗稻瘿蚊基因且农艺性状优良的单株与轮回亲本继续回交,自交2次后选择抗性基因纯合且农艺性状优良的株系进行抗稻瘿蚊苗期鉴定(图1)。
1.2.2 分子标记 选用与基因Gm5连锁及在抗源GXM-002-1与轮回亲本R553间表现多态的分子标记12M-29(F:5'- AGGTGTAGTTCTTGGCAAAT-3',R:5'- TCTCTCTCGTGTTCTTGGAT -3')进行跟踪检测,在分离群体中选择携带目标基因的株系。引物由北京六合华大基因科技有限公司合成。
1.2.3 PCR反应体系和扩增程序 参照CTAB提取法从水稻叶片中提取DNA样品,参考李孝琼等[23]的方法进行PCR扩增、电泳和银染检测,略有改动。PCR反应体系10.0 μL:10×PCR Buffer(含Mg2+)1.0 μL,10 mmol/L dNTP 0.2 μL,5 U/μLTaqDNA聚合酶0.1 μL,10 μmol/L上、下游引物各0.2 μL,DNA模板1.0 μL,ddH2O调至总体积10.0 μL。扩增程序:95 ℃预变性5 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 45 s,进行32个循环;72 ℃延伸5 min,4 ℃保存。PCR扩增产物在8%聚丙烯酰胺凝胶上进行电泳分离和银染,用保鲜膜包胶保存并记录结果。

表1 水稻苗期稻瘿蚊抗性鉴定评价标准
1.2.4 稻瘿蚊抗性鉴定 参考国际稻瘿蚊苗期群体鉴定法进行稻瘿蚊抗性鉴定[3],将鉴别品种播于搪瓷育苗盆内,每盆播12行,每行20~25苗,每个品种播2行,设4个重复。以多抗1号为抗蚊对照,TN1为感蚊对照。在秧苗2~3叶期,将育苗盆移至盛水的水泥池中,盖上网罩,按每15~20苗接入经交配的雌蚊1头,在成虫产卵期和幼虫孵化期用湿布加盖保湿。接蚊后18~25 d,即乙型标葱出现后,调查各鉴定材料的抗感株数,其中,稻株心叶抽不出、叶鞘畸形膨大呈“大肚秧”或叶鞘伸长成管状“标葱”为感蚊株,稻株心叶正常生长为抗蚊株;统计各株系标葱率及抗性级别(表1)。以感蚊对照TN1标葱率在60%以上为试验有效标准。
1.2.5 田间农艺性状调查 2020年7月中旬播种,8月上旬移栽,移栽后常规田间管理,于成熟期选取中间2行的3~5株,考察株高、有效穗、穗长、每穗总粒数、结实率和千粒重等主要农艺性状。
1.3 统计分析
试验数据采用Excel 2010进行整理,采用SPSS 19.0进行差异显著性分析。
2 结果与分析
2.1 抗稻瘿蚊水稻改良株系的分子标记辅助选择和评价
2017年晚造,以R553与GXM-002-1进行杂交;2018年早造,以R553为母本与F1回交,晚造用分子标记从40株BC1F1中选择2株含抗性基因的优良单株与R553再回交;2019年早造,利用分子标记从32株BC2F1中选择2株含抗性基因的优良单株进行自交收种,晚造在苗期先用分子标记从251株BC2F2中检测到抗性基因纯合单株58株(部分标记检测结果见图2),全部移栽至大田,从中筛选得到农艺性状与R553相近的4个改良株系R553GM-15、R553GM-19、R553GM-31和R553GM-47。
2.2 水稻亲本及改良株系苗期的稻瘿蚊抗性表现
2020年早造收获BC2F3世代种子,在8月对亲本和4个株系的BC2F4世代进行稻瘿蚊苗期抗性鉴定(表2和图3)。由表2可知,R533对稻瘿蚊表现高感,GXM-002-1表现高抗,与抗蚊对照多抗1号相似;改良株系中的R553GM-15、R553GM-19和R553GM-31对稻瘿蚊的抗性达中抗水平,R553GM-47达抗水平。说明通过分子标记辅助选择,可获得稻瘿蚊抗性明显提高的株系。
2.3 水稻亲本及改良株系的主要农艺性状调查结果
2020年晚造,在正常田间水肥管理条件下,对亲本和4个改良株系的农艺性状进行调查,结果(表3)表明,R553GM-15和R553GM-31的株高与R553差异不显著(P>0.05,下同),R553GM-19和R553GM-47的株高极显著高于R533(P<0.05,下同);4个改良株系的有效穗数和千粒重与R553均无显著差异;除R553GM-19的穗长显著短于R553外,其余3个改良株系的穗长与R553均无显著差异;R553GM-31的每穗总粒数与R553无显著差异,其余3个改良株系的每穗总粒数均极显著少于R553(P<0.01,下同);R553GM-15的结实率与R553无显著差异,其余3个改良株系的结实率均极显著低于R533;4个改良株系的粒长均短于R533,其中,R553GM-15的粒长与R533差异不显著,R553GM-19与R553GM-31的粒长显著短于R553,而R553GM-41的粒长极显著短于R553;4个改良株系的粒宽均大于R533,其中,R553GM-15和R553GM-19与R533差异不显著,另2个改良株系的粒宽显著大于R533。可见,导入抗稻瘿蚊基因后改良株系各农艺性状中至少有1个株系与轮回亲本无显著差异,说明采用分子标记辅助将GXM-002-1的抗稻瘿蚊基因Gm5导入优质恢复系R553具有可行性。
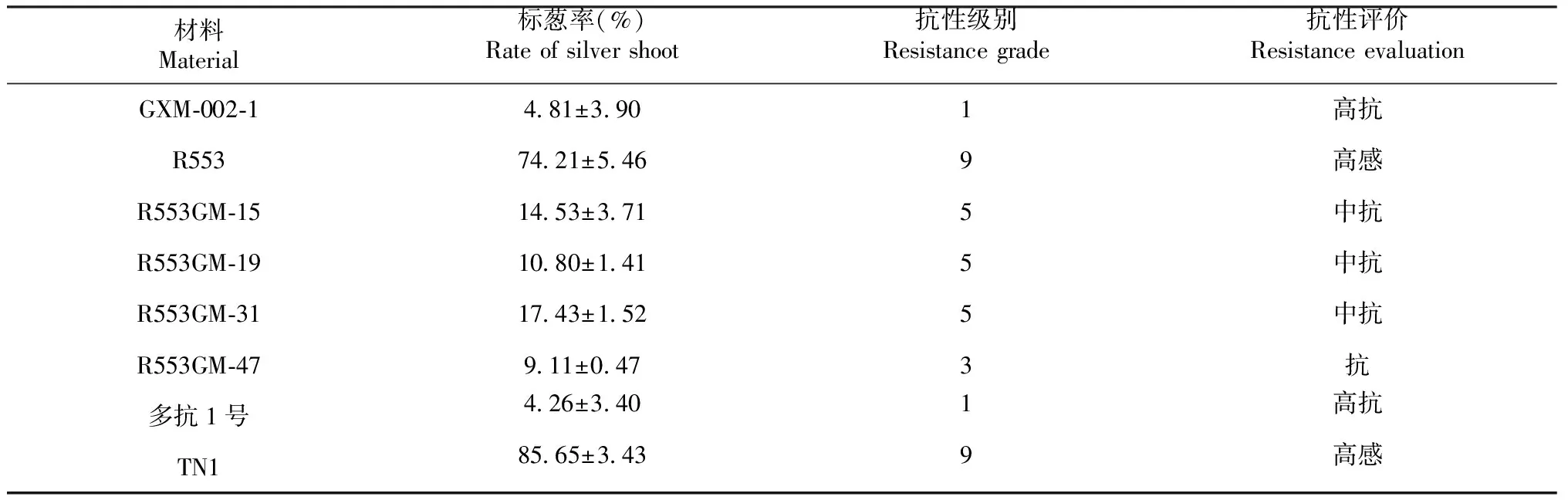
表2 苗期水稻的稻瘿蚊抗性评价结果
3 讨 论
分子标记辅助选择是一种高效的育种方法,可精确跟踪目标基因进行选择,已成功培育了大批优良品系。王春连等[24]筛选到与抗白叶枯基因Xa23紧密连锁的EST标记C189,其标记辅助选择的正确率接近100.0%。李进波等[25-26]利用EST标记C189进行辅助选择,也获得3份优良抗性杂交稻恢复系;通过与Bph14和Bph15基因紧密连锁的SSR标记MRG2329和MS5的辅助选择,获得多份抗褐飞虱株系,并获得6份聚合双基因的高抗株系。Qing等[27]采用分子标记辅助选择将抗褐飞虱基因Bph3导入桂恢582和桂7571,使其抗性明显增强。刘驰等[28]通过分子标记辅助选择培育出聚合了抗稻瘟病基因Pi-ta、Pib、Pi54和抗白叶枯病兼抗细条病基因xa5的多抗优质强恢复系桂恢663。在抗稻瘿蚊方面,黄显波等[29-30]、李云等[31]利用与Gm6基因连锁的分子标记检测明恢884、双抗明占、抗15-1、抗8和淦恢3等多个恢复系,确定其含有抗稻瘿蚊Gm6基因。林成豹等[32]采用连续回交结合分子标记辅助选择方法将Gm6基因导入元丰B,再与元丰A多代回转育成抗稻瘿蚊三系不育系BSC9247。Kumar等[33]利用分子标记辅助选择将Xa21与Gm4和Gm8基因分别聚合到恢复系RPHR-1005中,提高了RPHR-1005对白叶枯与稻瘿蚊的抗性。Zhou等[21]筛选出几个与Gm5基因紧密连锁的分子标记,可用于不同材料间的分子标记辅助选择。本研究利用与抗稻瘿蚊基因Gm5紧密连锁的在R553与GXM-002-1间有多态性的标记M12-29,采用杂交、回交方法,在分离群体中利用标记进行辅助选择,结合抗性鉴定与农艺性状选择,获得了对稻瘿蚊抗性提高的优良株系,与上述研究结果一致,可为水稻抗稻瘿蚊育种提供重要的遗传基因材料。

表3 水稻亲本及抗稻瘿蚊改良株系的农艺性状比较
农艺性状一直是水稻育种中优先考虑的重要因素,在分子标记辅助选择中,不仅要跟踪目标基因,更要保持好亲本的优良农艺性状。周元飞等[34]运用分子标记辅助选择将抗白叶枯基因Xa21转入轮回亲本9311,获得4个优良抗性株系,除株高和叶色与轮回亲本略有不同外,其他性状差异不明显,且与培矮64S配组的F1与两优培九各项指标非常相似。潘晓飚等[35]以明恢86、蜀恢527和浙恢7954为受体,以三黄占2号和IRBB23为供体,通过标记辅助聚合4个抗稻瘟病位点和Xa23基因,获得5个抗性改良恢复系,与原恢复系相比,单株产量无显著差异,穗长缩短,每穗粒数减少,千粒重下降,株高降低,但结实率有提高趋势;在与II-32A的测交种中,除明浙-G1-G2-G8-Xa23/II-32A的单株产量显著提高外,其余改良恢复系/II-32A的单株产量与原恢复系/II-32A的单株产量相仿。王飞名等[36]利用标记辅助改良黄华占,获得3个抗旱且携带Bph15基因的纯合株系,这3个株系的产量、每穗总粒数、每穗实粒数、有效穗数和结实率等与黄华占均无显著差异,株高极显著增高,其中有2个株系的穗长和千粒重存在显著差异;3个株系的株高与轮回亲本均存在极显著差异。本研究中,4个改良株系中有2个株系的株高与轮回亲本相近,另2个株系的株高显著高于轮回亲本,理论单株产量、有效穗数和千粒重与轮回亲本无显著差异,与上述研究结果相似。此外,本研究中4个改良株系的其他农艺性状如每穗总粒数、结实率、穗长、粒长和粒宽等性状中,至少有1个株系与轮回亲本无显著差异,表明在抗性改良中,仍会产生较大的遗传累赘效应,需通过多代回交或扩大选择群体来获得抗性与优良农艺性状俱佳的株系。
4 结 论
以高抗稻瘿蚊材料GXM-002-1为抗源供体亲本,以优质水稻恢复系R553为轮回亲本,通过回交育种结合分子标记辅助选择与抗虫鉴定,可获得稻瘿蚊抗性达抗或中抗、农艺性状表现良好的改良株系。
