Morphological and phylogenetic diversity of magnetotactic bacteria and multicellular magnetotactic prokaryotes from a mangrove ecosystem in the Sanya River, South China*
2021-12-09HaitaoCHENJinhuaLILongFeiWUWeiJiaZHANG
Haitao CHEN , , Jinhua LI , Long-Fei WU , Wei-Jia ZHANG ,
1 Beijing Key Laboratory of Biological Electromagnetism, Institute of Electrical Engineering, Chinese Academy of Sciences,Beijing 100190, China
2 France-China Joint Laboratory for Evolution and Development of Magnetotactic Multicellular Organisms, Chinese Academy of Sciences, Beijing 100029, China
3 Deep-Sea Microbial Cell Biology, Department of Deep Sea Sciences, Institute of Deep-Sea Science and Engineering, Chinese Academy of Sciences, Sanya 572000, China
4 Key Laboratory of Earth and Planetary Physics, Institute of Geology and Geophysics, Chinese Academy of Sciences, Beijing 100029, China
5 LCB, Aix Marseille University, CNRS, Marseille 13402, France
Abstract Magnetotactic bacteria (MTB) are morphologically and phylogenetically diverse prokaryotes commonly able to produce magnetic nanocrystals within intracellular membrane-bound organelles (i.e.,magnetosomes) and to swim along geomagnetic f ield lines. We studied the diversity of MTB in the samples collected from a mangrove area in the Sanya River, Hainan, South China, using microscopic and microbial phylogenetic methods. Results of microanalysis and observation in microscopy and energy dispersive X-ray spectroscopy (EDXS) reveal a highly morphological diversity of MTB including unicellular cocci,vibrios, rod-shaped bacteria, and three morphotypes of multicellular magnetotactic prokaryotes (MMPs).In addition, analysis of the 16S rRNA gene showed that these MTB were clustered into 16 operational taxonomic units affi liated to the Alpha-, Delta-, and Gamma-proteobacteria classes within the Proteobacteria phylum. Meanwhile, by using the coupled f luorescence and transmission electron microscopy analysis, rodshaped bacteria, vibrio, and cocci were phylogenetically and structurally identif ied at the single-cell level.This study demonstrated highly diverse MTB communities in the mangrove ecosystem and provide a new insight into the overall diversity of MTB.
Keyword: magnetotactic bacteria; diversity; magnetosome; phylogenetic analysis; coordinated f luorescence in-situ hybridization transmission electron microscopy (FISH-TEM); mangrove
1 INTRODUCTION
Magnetotactic bacteria (MTB) are a group of morphologically and phylogenetically heterogeneous prokaryotes. They are capable of sensing and changing their orientation in accordance with the direction of the geomagnetic f ield, a behavior called magnetotaxis(Faivre and Schüler, 2008; Komeili, 2012). This unique capability is facilitated by special organelles that are intracellularly synthesized, membraneenclosed ferromagnetic nanocrystals of magnetite(Fe3O4) and/or greigite (Fe3S4), called magnetosomes(Bazylinski and Frankel, 2004). Because of the special properties of these magnetosomes, MTB have been exploited for applications in diverse disciplines from geobiology to biotechnology (Komeili, 2012; Li et al., 2013). In addition, the fossil magnetosomes preserved in sediments (called magnetofossils) could be used as potential proxies for reconstructing the paleoenvironment and paleo-climate changes and understanding the origin and evolution of early life(Pan et al., 2005; Kopp and Kirschvink, 2008; Li et al., 2020b).
MTB are widely distributed in various aquatic environments such as lakes, rivers, ponds, estuaries,lagoons, marine sediments, and even deep-sea extreme environments (Petersen et al., 1986; Lins et al., 2007; Jogler et al., 2010; Chen et al., 2015a; Li et al., 2016; Lin et al., 2017, 2018; Liu et al., 2017).Diff erent morphologies have been described,including single cells in forms of cocci, ovoid, beanshaped, rod-shaped, and spirilla, and groups of cell aggregates composing of up to over forty ovoid cells,which are known as multicellular magnetotactic prokaryotes (MMPs) (Abreu et al., 2007; Lins et al.,2007; Isambert et al., 2007; Pan et al., 2008; Lin et al.,2009; Wenter et al., 2009; Zhou et al., 2011; Qian et al., 2020). Phylogenetically, most known MTB are affi liated with the phyla Proteobacteria, Nitrospirae,Planctomycetes, the candidate phyla “Omnitrophica”and “Latescibacteria” (Lefèvre et al., 2009; Lefèvre and Bazylinski, 2013; Lin and Pan, 2015; Dziuba et al., 2016; Lin et al., 2017; Liu et al., 2021). Despite of the abundant data regarding cellular and crystal characterization, and phylogenetic diversity of MTB(Lins et al., 2000; Zhou et al., 2011; Wang et al., 2012;Chen et al., 2015b; Lin et al., 2017), only a few MTB strains have been successfully isolated and cultivated and the physiological characters of certain phylogenetic group of MTB remain obscure (Schüler and Köhler, 1992; Nakamura et al., 1993; Lefèvre et al., 2009, 2011; Schübbe et al., 2009; Zhu et al., 2010;Wang et al., 2013). Recently, f luorescence in-situ hybridization coupled with scanning (FISH-SEM) or transmission electron microscopy (FISH-TEM) was developed (Li et al., 2017). It enabled distinguishing MTB from other microorganisms in environmental samples at the single-cell level, and connecting phylogeny with the cell ultrastructure and magnetosome morphology of uncultured MTB strains(Li et al., 2017, 2019; Zhang et al., 2017; Koziaeva et al., 2020; Qian et al., 2020; Liu et al., 2021).
Previous studies have shown that diversity and communities of MTB, as well as the magnetic crystal morphology and spatial arrangement of magnetosomes, varied depending on environmental factors, such as the strength of the earth’s magnetic f ield, geographic latitude, temperature, salinity, pH,and nutrients (Lefèvre and Wu, 2013; Pósfai et al.,2013; Lin et al., 2014; Li et al., 2020a, b; Liu et al.,2021). Mangroves are peculiar ecosystems connecting terrestrial and marine environments in tropical and subtropical regions. The woody plants growing in coastal wetlands are periodically f looded by tidal water, which possibly result in a diverse and dynamic microbial community (Alongi, 2002). In general,study of MTB in mangrove ecosystem is lacked, with only limited reports of the presence of magnetite magnetic minerals and coccoid MTB in mangroves samples (Kannapiran et al., 1999; Maloof et al., 2007;Lin et al., 2012). To better understand the phylogenetic diversity and characterization of MTB in mangroves,we collected MTB from mangrove estuary waters in Sanya, southern China. Optical and electronic microscopes were used to analyze their abundance and morphology. Taxonomic identif ication and phylogeny study of collected cells were carried out by analyzing the 16S rRNA gene sequences. Additionally,we characterized the morphology of bacterial cells and magnetosomes of certain phylogenetic group of MTB with FISH-TEM analysis.
2 MATERIAL AND METHOD
2.1 Sampling and magnetic collection
Sediment samples were collected from the tidal f lats of mangrove located at the mouth of the Sanya River (Sanya, China; 18°15.242′N, 109°30.585′E) in spring of 2014 and 2015. To carry out enrichment of magnetotactic bacteria, approximately 300-mL sediments were transferred to 0.5-L plastic bottles covered with ~200 mL of seawater collected from the surface layer (~3-10 cm) at the sampling sites. A piece of magnet was stick to the bottle wall approximately 2 cm below the interface between the sediment and surface water with the southern pole facing interior. The bottles were covered loosely and stored at room temperature under dim light for a week for the enrichment. The pH, temperature, and salinity of sediments measured in-situ were 8.2, 23 °C, and 21.33, respectively. The concentrations of NH4+, NO2ˉ,NO3ˉ, SO42ˉ, and PO43ˉ in pore water were determined by using Elemental analyzer (Hach DR890, USA) to be 8.4, 0.02, 1.23, 2017, and 7.3 mg/L, respectively.Collection of MTB was purif ied by race-track method(Wolfe et al., 1987; Li et al., 2010).
2.2 Optical microscopy
The quantity and community of MTB in the microcosms were periodically checked with hanging drop method using an optical microscope at a 40×magnif ication (Olympus BX53, Japan). Based on these microscopic images, cell size of MMPs was measured with software ImageJ 1.51i.
2.3 Transmission electron microscopy (TEM)
The purif ied sample was deposited on a carboncoated copper grid supported by formvar for TEM observation. A JEM-2100 TEM equipped with an Oxford SDD detector (X-MaxN 80T) and an accelerating voltage of 200 kV was used for morphological and cellular structure studies. Under the same conditions, the chemical composition and structure of magnetic crystals were analyzed by energy dispersive X-ray spectroscopy (EDXS), and high-resolution TEM (HRTEM). The crystal length(along the long axis) and width (perpendicular to the long axis) of the magnetosomes were measured from the TEM images by software ImageJ 1.51i. Three MMPs cells of each morphological type were analyzed for the characterization of the magnetosome crystals.
2.4 PCR amplif ication, 16S rRNA gene sequencing,and phylogenetic analyses
16S rRNA gene sequences of the samples were amplif ied as described previously by Chen et al.(2015a). With bacteria-specif ic primers 27F and 1492R, PCR reactions were performed using a T100TMThermal Cycler (Bio-Rad, America). The cycling conditions were 4 min at 94 °C for initial denaturation;30 cycles of 1 min at 94 °C, 45 s at 55 °C, and 1.5 min at 72 °C; and a f inal extension of 10 min at 72 °C.PCR products were purif ied, ligated into the pMD19-T cloning vector (TaKaRa, Japan) and transformed intoEscherichiacoliDH5α to construct a 16S rRNA gene clone library. Positive clones were conf irmed by PCR amplif ication with primer M-13 and RV-M. 60 clones were sequenced by Shanghai Invitrogen Biotechnology Co., Ltd. Using BLAST search program on the NCBI website, the 16S rRNA gene sequences retrieved in this study were f irst analyzed.The related sequences were then aligned with their close relatives using the ClustalW algorithm, and a phylogenetic tree was subsequently constructed using the neighbor-joining method in MEGA version 6.0(Kumar et al., 2016). The sequences were submitted in GenBank database with accession numbers MW356763-MW356778.
2.5 Coupled FISH-TEM analysis
In this study, the Alphaproteobacteria-specif ic probe ALF968 was used for f luorescence in-situ hybridization (Neef, 1997). The bacterial universal probe EUB338 was used as a control (Amann et al.,1990). The probe ALF968 was labeled at the 5'end with hydrophilic sulfoindocyanine Cy3, and the universal probe was labeled at the 5'end with f luoresce in phosphoramidite (FAM). Both probes were synthesized and f luorescently labeled by Invitrogen Co., Ltd. Coupled FISH-TEM analysis was based on the procedure described in Li et al. (2017). The collected MTB cells were f ixed with 4%paraformaldehyde as described by Pernthaler et al.(2002) and then washed in 1×phosphate buff er saline.By centrifugation, the cells were f ixed (dehydrated)for the second time with 50%, 80%, and 100%ethanol, respectively. Then, the cells were resuspended in hybridization buff er containing probe (50 ng/μL)for in-situ hybridization at 46 °C for 3 h. Afterwards,the samples were washed three times with elution buff er and water respectively. An amount of 5-μL washed cells was deposited onto letter-indexed carbon-coated copper grids, dried in air at ambient temperature. The results were recorded using a f luorescence microscope (Olympus BX51) and TEM(JEM-2100 TEM).
3 RESULT AND DISCUSSION
3.1 Morphologic and chemical features of MTB from Sanya mangrove
Living MTB collected from sediment samples were observed with optical microscopy. The cells abundance was approximately 103cells/cm3. The MTB collected from Sanya mangrove were morphologically diverse and consisted of both unicellular MTB and multicellular MMPs. The multicellular MMPs represented as an abundant population in the samples collected. They exhibited a spherical morphology (s-MMPs) with a diameter of 4.6±0.2 μm (Fig.1). Like s-MMPs observed in other marine environments, they showed classic escape motility or “ping-pong” motion that undergoes backward excursions followed by a forward movement (Lins et al., 2007; Wenter et al., 2009;Zhou et al., 2011).

Fig.1 MTB collected from the mangrove of the Sanya River
The s-MMPs could be divided into three diff erent morphotypes in shape, size, and composition of magnetosomes (Fig.2 & Table 1). Type-I likely contains only greigite-type magnetosomes on average size of 77±11 nm in length (the number of magnetosomes:n=263; Table 1). Type-Ⅱ likely contains only bullet shaped magnetite magnetosomes on average size of 78±18 nm in length and 34±4 nm in width (n=125). Type-Ⅲ contains both magnetitetype and greigite-type magnetosomes, the average sizes of magnetosomes are 80±19 nm for greigite crystals, and 88±19 nm in length and 34±5 nm in width (n=110) for bullet-shaped magnetite crystals(Fig.2g-i). The three diff erent types of s-MMPs were also reported in the lagoons of Brazil (Lins et al.,2007). Comparatively, the average size of magnetosomes formed by the Type-I was slightly larger than those observed in the lagoons in Brazil(70±8 nm) and Yuehu Lake (63.9±9.3 nm) (Lins et al.,2007; Zhang et al., 2014). The average size of bulletshaped magnetosomes produced by Type-Ⅱ was smaller than those in previously reported s-MMPs collected elsewhere (Lins et al., 2007; Wenter et al.,2009; Zhou et al., 2011, 2013; Zhang et al., 2014). To summarize, the s-MMPs discovered from the mangrove sediments of the Sanya River resembled s-MMPs in other ecosystems in cell size (Table 1) and movement behavior, but diff ered slightly in the morphology of magnetosomes (Lins et al., 2007;Wenter et al., 2009; Zhou et al., 2011). This agrees with current understanding that magnetosome biomineralization is biologically controlled, yet aff ected by environmental factors of habitats at certain degree (Li and Pan, 2012).
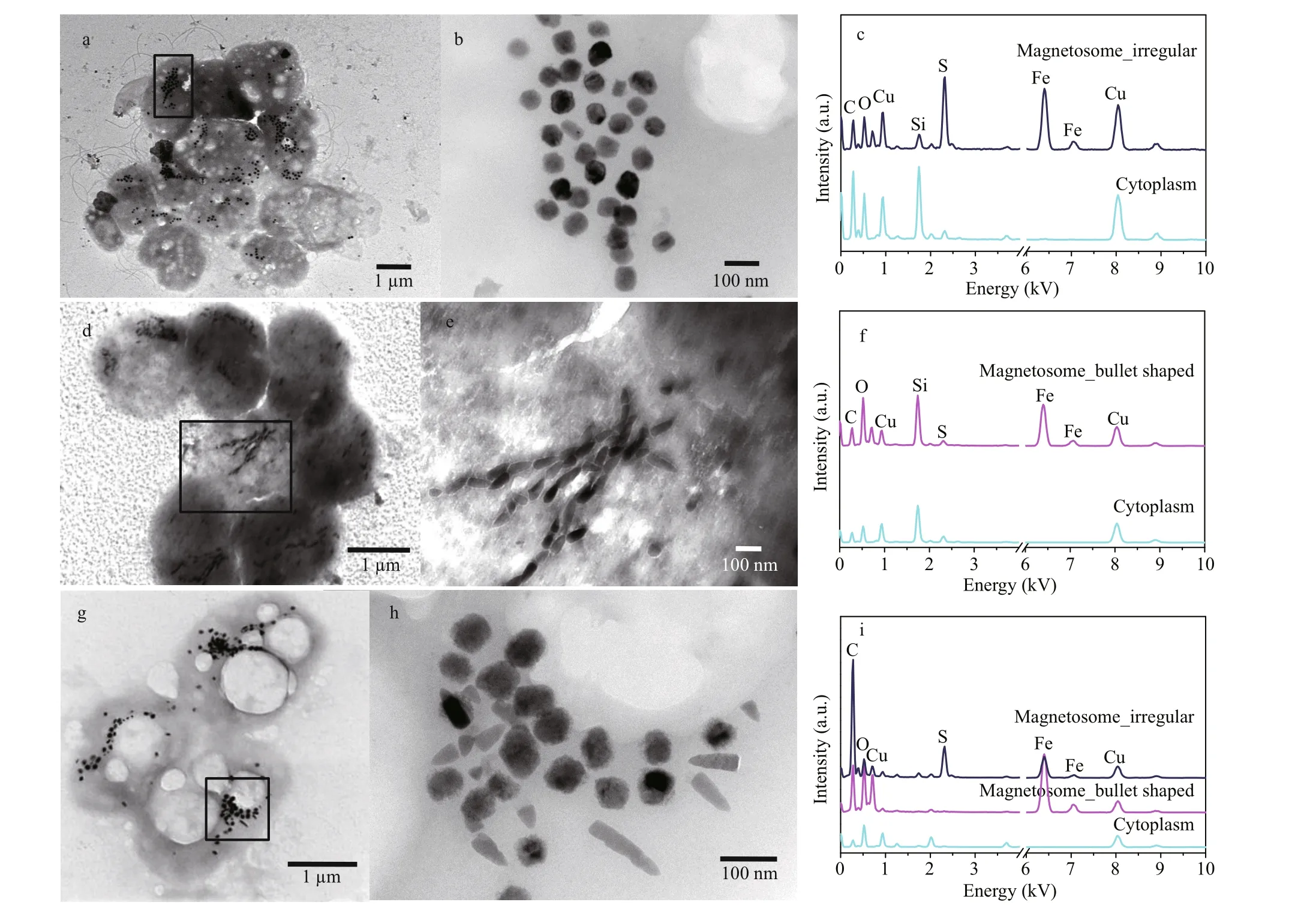
Fig.2 Morphological and chemical features of MMPs collected from Sanya mangrove environment
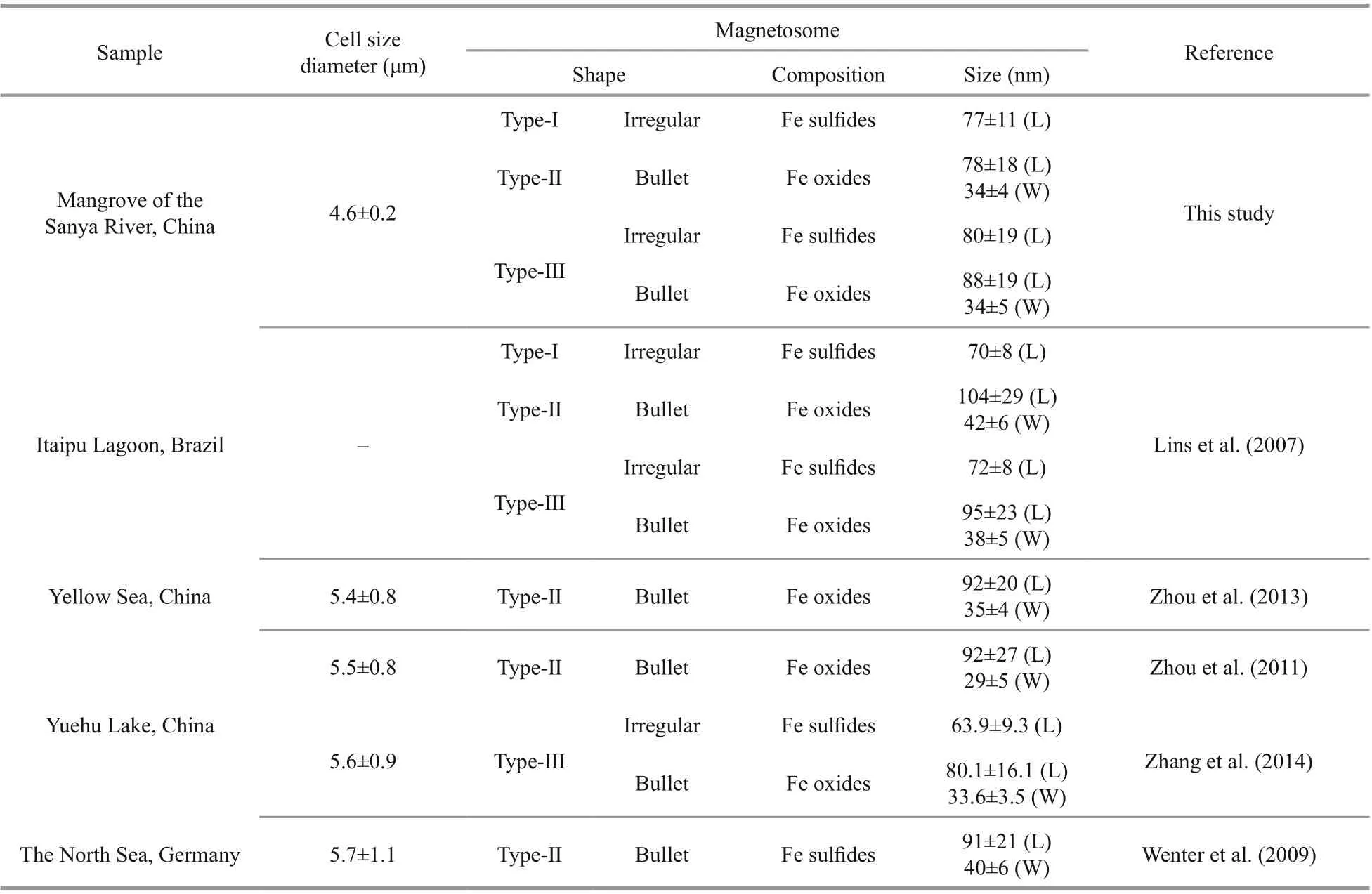
Table 1 Comparison of morphological characteristics of s-MMPs
As expected, TEM observations revealed a highly diverse unicellular MTB in terms of cellular and magnetic crystal morphologies. As shown in Fig.3,we found elongated prismatic magnetosomes in cocci(Fig.3a-h), vibrio (Fig.3j), and rod-shaped bacteria(Fig.3k-l); and bullet-shaped magnetosomes in magnetotactic cocci (Fig.3i). The magnetosomes are arranged in single chains (Fig.3a-b & j-k), double chains (Fig.3c, i, & l), multiple chains (Fig.3d, e, f, &g), or in scattered distribution (Fig.3h). Consistent with previous studies (Spring et al., 1993; Cox et al.,2002; Bazylinski et al., 2004; Lefèvre et al., 2011; Li et al., 2017, 2019, 2020a; Rivas-Lamelo et al., 2017;Schulz-Vogt et al., 2019), MTB collected from Sanya mangrove environments contain various micrometersized intracellular inclusions (e.g., sulfur-rich or polyphosphate particles, or polyhydroxyalkanoate granules) in addition to the nanometer-sized magnetosomes, indicating their involvement in driving biogeochemical cycles of key elements in nature (Li et al., 2020a).
Interestingly, we found a special type of large rodshaped bacteria whose irregularly shaped magnetosomes dispersed evenly in the cell (Fig.4a).EDXS microanalysis showed that the magnetosomes are rich in iron and sulfur (Fig.4b). HRTEM imaging further indicated that the mineral phase of irregular crystal was greigite (Fig.4c-d). Large rod-shaped MTB have been reported in the Great Boiling Springs geothermal f ield in Gerlach, Nevada, USA, and freshwater niche previously (Lefèvre et al., 2011;Wang et al., 2013). But, in both cases, the irregularly shaped greigite magnetosomes were aligned in multiple chains. Whether the large rod-shaped MTB present in Sanya mangrove belonged to a diff erent group, or exhibited a diff erent magnetosome spatial arrangement due to the changed environment parameters require further investigations.
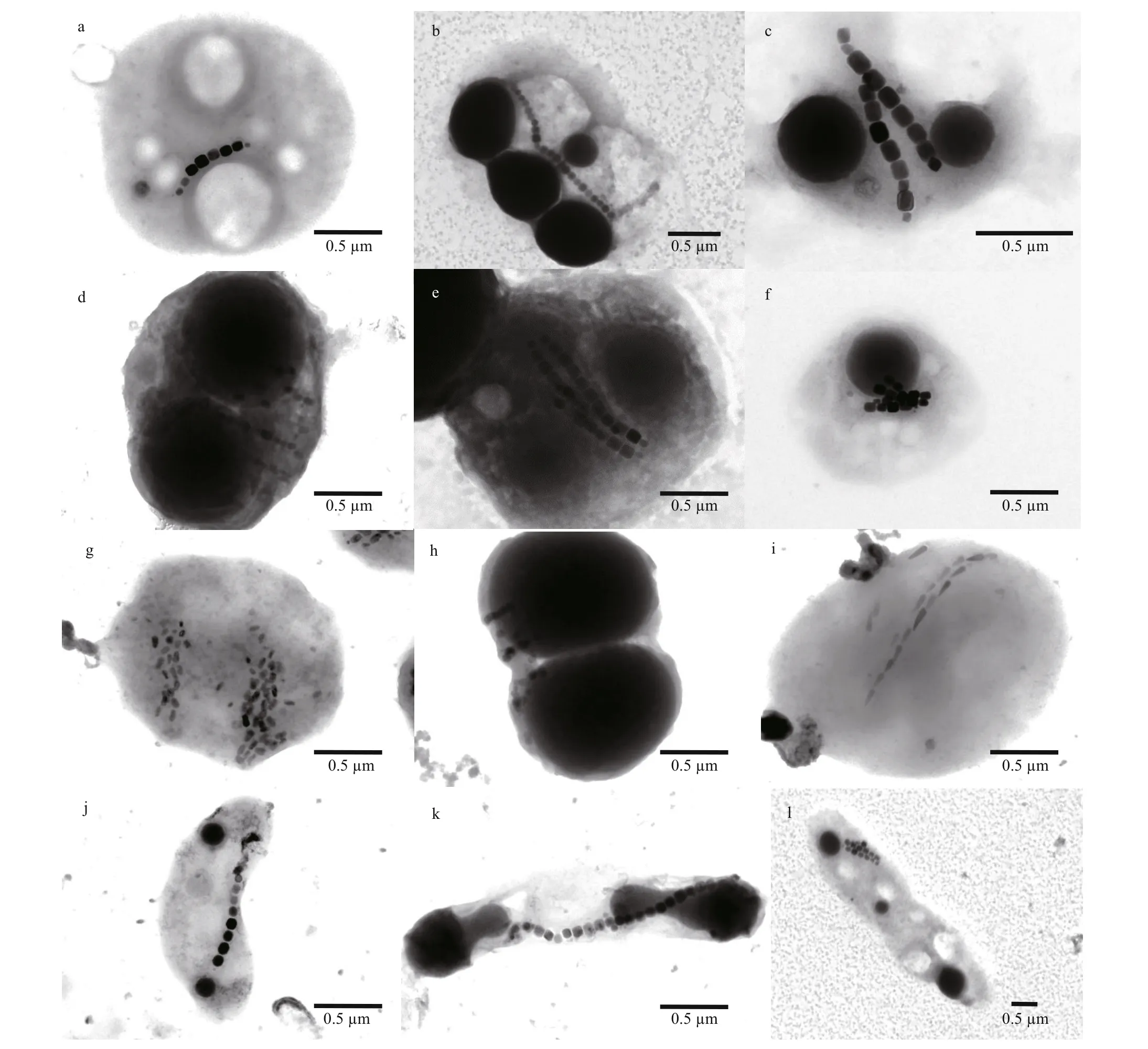
Fig.3 Transmission electron micrograph of unicellular MTB collected from Sanya mangrove environments
3.2 Phylogenetic diversity of MTB collected from Sanya mangrove environment
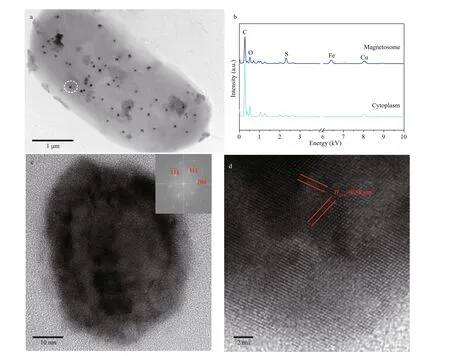
Fig.4 Large rod-shaped MTB collected from Sanya mangrove environments
Phylogenetic analysis based on 16S rRNA gene sequences was performed to determine the community structure of MTB. In total 16 operational taxonomic units (OTUs) were identif ied at the 98% similarity level, all affi liated with the Proteobacteria phylum(Fig.5). The OTU 1 and 2 shared 93.5% and 97.7%sequence identities to MMPs of the uncultured Deltaproteobacterium mmp2_9 (Simmons and Edwards, 2007), and 90.5% and 88.7% identities to theCandidatusMagnetoglobus multicellularis from Araruama Lagoon in brazil, respectively (Martins et al., 2009). Although our results showed the s-MMPs found in Sanya were similar to the MMPs of Lagoon in cell morphology and magnetosome characteristics,the relatively low similarity of 16S rRNA gene suggested that these two OTUs might represent novel MMPs species. The OTU 3 is 98.3% identical to the culturedDesulfamplusmagnetovallimortisBW-1 within the Deltaproteobacteria (Lefèvre et al., 2011;Descamps et al., 2017). Three OTUs (OTU 4 to 6)belong to the Alphaproteobacteria, which were most closely related to known magnetotactic cocci (90% to 98%). Additionally, ten OTUs (OTU 7 to 16) belonged to the Gammaproteobacteria, which accounted for more than 47% of all retrieved sequences. OTU 16 was 95.6% identical to the cultured Gammaproteobacteria BW-2 (Lefèvre et al., 2012).OTUs 14 and 15 were closely related to previously described Gammaproteobacterium GRS-1 MTB(Taoka et al., 2014). OTU 13 had a maximum similarity of 90.6% with the cultured Gammaproteobacteria SHHR-1 (Zhang et al., 2017).Sequences belonging to OTU 7 to 12 were highly divergent from known MTB species and might therefore represent novel branches. Notably, the coccoid MTB identif ied in the saline samples from the mangrove swamps (Wenchang, China) by Lin et al. (2012) were affi liated with the uncultivated AlphaproteobacteriaMagnetococcus. Comparing to that, the MTB in the mangroves of Sanya showed higher diversity in respect to morphology and phylogeny. This f inding expands the knowledge of the community composition and distribution of MTB in mangrove environments, and suggests the highly adapted MTB to complex environments.
3.3 Identif ication of uncultured MTB

Fig.5 Phylogenetic tree based on 16S rRNA gene sequences showing the relationship of the MTB obtained from the Sanya River sediment within its closest relatives
By the time this work was carried out, a large number of studies have shown that magnetotactic cocci belong to Alphaproteobacteria. Magnetotactic cocci with diff erent characteristics were observed in the sediments of Sanya mangrove. Along with that,several 16S sequences retrieved from this sample belonged to the cluster containingMagnetococcuswithin Alphaproteobacteria. To identify the morphology and magnetosomes characteristics of Alphaproteobacteria MTB in mangrove, we carried out FISH-TEM analyses using a probe specif ic to the 16S rRNA of Alphaproteobacteria group (ALF968)(Neef, 1997) and a universal bacterial probe EUB338(Amann et al., 1990) as a control. As shown in Fig.6,the Alphaproteobacteria-specif ic probe ALF968 hybridized with vibrio, cocci, and rod-shaped MTB from mangrove sediment. TEM analysis showed they synthesized elongated prismatic magnetite crystals (Fig.6c-f).
In addition, based on 16S rRNA gene analysis we found that ten OTUs may be belonging to Gammaproteobacteria. OTU 15 shared 92.4%sequence identities to Gammaproteobacterium GRS-1 MTB. To identify whether this group is a new genus of MTB, we designed a specif ic probe for OTU 15,but our results showed that cells recognized by this probe did not possess magnetosomes (data not shown). We cannot rule out the possibility that this bacterium was MTB but did not synthesize magnetosomes under current condition, but it is also possible that the OTU 15 may not be MTB even though its 16S sequence shows similarity with known MTB and the accuracy of judging the diversity of MTB based on 16S rRNA gene sequences is worth discussion.
4 CONCLUSION
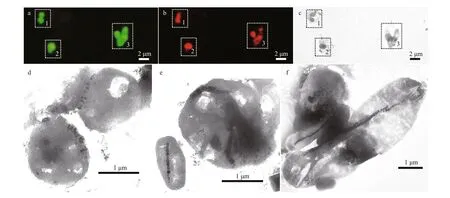
Fig.6 Correlative FISH-TEM analysis of uncultured MTB from the tidal f lats of mangrove located in the mouth of theSanya River in China based on in-situ hybridization with the Alphaproteobacteria-specif ic probe ALF968
Both microscopy and molecular analyses revealed a high diversity of MTB in Sanya mangrove environments. Additional to unicellular cocci, vibrios,and rod-shaped bacteria, three types of Desulfobacteraceae family belonging to Deltaproteobacterium of s-MMPs were observed:Type-I forming only irregular greigite magnetosomes,Type-Ⅱ forming only bullet-shaped magnetite magnetosomes, and Type-Ⅲ producing both bulletshaped magnetite and irregular greigite crystals.Consistent with morphological diversity, phylogenetic analyses based on the 16S rRNA genes sequencing revealed 16 OTUs of MTB that affi liated to Alphaproteobacteria, Deltaproteobacteria, and Gammaproteobacteria. This f inding suggests that the mangrove environments contain abundant and potentially novel MTBs that worth further investigations.
5 DATA AVAILABILITY STATEMENT
The data used in the paper are available from the authors.
6 ACKNOWLEDGMENT
We are grateful to Qunjian YIN for help in sample collection. The authors also thank the anonymous reviewers for valuable comments and the editors for careful editing.
杂志排行

Journal of Oceanology and Limnology的其它文章
- MamZ protein plays an essential role in magnetosome maturation process of Magnetospirillum gryphiswaldense MSR-1*
- Magnetotactic bacteria from the human gut microbiome associated with orientation and navigation regions of the brain*
- How light aff ect the magnetotactic behavior and reproduction of ellipsoidal multicellular magnetoglobules?*
- Biocompatibility of marine magnetotactic ovoid strain MO-1 for in vivo application*
- Determination of the heating effi ciency of magnetotactic bacteria in alternating magnetic f ield*
- An approach to determine coeffi cients of logarithmic velocity vertical prof ile in the bottom boundary layer*