施氮对镉胁迫下龙葵生长和光合特性的影响*
2021-12-09刘新亮唐星林黄文超骆昱春王丽艳
刘新亮,唐星林,黄文超,骆昱春,王丽艳
(江西省林业科学院,江西 南昌 330032)
镉(Cd)是一种重金属元素,对动植物有高度的毒害性。由于矿产开采、污水排放、农药及化肥施用等工农业生产活动,我国土壤Cd污染变得越来越严重[1]。土壤Cd易被植物根系吸收,并在植物器官内积累,对植物产生毒害[2]。Cd毒害会导致植物营养缺乏、叶绿素合成受阻、光合能力下降等,抑制植物正常的生长发育,严重时会导致植物死亡[2-3]。Cd还可以在作物可食用部分积累,进入食物链,威胁人类健康[1]。因此,土壤Cd污染防治已成为我国亟待解决的重大问题之一。植物修复技术是一项利用植物来稳定、提取土壤重金属的技术,具有成本低、无二次污染、环境友好等优点[4]。重金属超富集植物对重金属有超量积累作用,是土壤重金属植物修复的优良材料,但大多数超积累植物存在生长缓慢、生物量小等特点,以致植物修复效率不高,限制了其在土壤重金属植物修复工程中的应用[5-6]。合理施肥可以促进植物生长,增大植株生物量及重金属富集量,具有经济、高效等优点,是提高植物修复效率的重要技术[7]。氮(N)是植物生长发育过程必需的大量元素,对植物生长和光合作用有显著的调节作用[8-9]。Cd胁迫会抑制植株硝酸还原酶活性,导致植株N含量下降[10]。有研究发现施N有利于保护Cd胁迫下植物光合器官结构,保持叶绿素合成途径稳定,增大PSII最大光化学量子产量、电子传递速率、气孔导度和光合速率,进而促进植株生长和生物量积累[11-14]。由此可见,N在缓解植物Cd胁迫的毒害方面起着重要作用。
龙葵(Solanumnigrum)是茄科(Solanaceae)茄属(Solanum)1 a生或多年生草本植物,魏树和等[15-16]发现龙葵具有Cd超积累特性,是修复Cd污染土壤的良好植物材料。杨容孑等[6]研究表明Cd会显著抑制龙葵生长,降低各器官生物量,而施N可以提高龙葵叶绿素含量、抗氧化酶活性,增加地上部分生物量和镉积累量。光合作用是植物有机物积累和生长基础,研究施N对Cd胁迫下龙葵光合作用的影响,可以更好地揭示N素对植物Cd毒害的缓解作用。但施N对Cd胁迫下龙葵光合特性的影响仍不清楚。本研究以龙葵为试验材料,探讨了不同施N水平对Cd胁迫下龙葵生长及光合作用的影响,以期获得合适的N肥施用浓度,为植物高效修复Cd污染土壤提供指导依据。
1 材料与方法
1.1 试验材料
植物材料为龙葵1 a生幼苗。供试土壤为园土(黄壤),采自江西省林业科学院试验基地0~20 cm表层土壤,将土壤自然风干、去除杂物,粉碎,过5 mm筛。泥炭采用丹麦进口的Pindstrup基质。栽培基质为黄壤和泥炭的混合种植土(质量比1︰1),其物理化学性质为速效氮53.7 mg/kg、速效磷95.0 mg/kg、速效钾110.0 mg/kg、有机质75.58 mg/kg、pH值5.75。氮肥为尿素[CO(NH2)2];试验试剂为CdCl2·2H2O。
1.2 试验方法
试验在江西省林业科学院温室大棚内(28°45′N,115°49′E)进行,采用盆栽方法,塑料花盆高15 cm、上口径18 cm、下口径13 cm。试验设计如下,(1)对照(CK)为无添加;(2)CdN0为20 mg/kg Cd;(3)CdN150为20 mg/kg Cd+150 mg/kg CO(NH2)2;(4)CdN300为20 mg/kg Cd+300 mg/kg CO(NH2)2;(5)CdN450为20 mg/kg Cd+450 mg/kg CO(NH2)2(不含背景值,以Cd2+计)。将准备好的栽培基质与CO(NH2)2、CdCl2·2H2O充分混匀,平衡18 d,待用。2019年5月底,将龙葵种子播种于育苗穴盘。待幼苗长出5片真叶时,挑选长势较一致幼苗进行试验,每盆1株,每个处理重复6次。进行日常浇水管理,用托盘垫在盆下收集渗漏的Cd和营养物质,并将其倒回盆中。
1.3 测定项目与方法
1.3.1 气体交换参数
2019年8月晴天9:00—16:30,采用Ciras-3便携式光合作用测定系统(Ppsystems,美国)进行气体交换数据的测定。植物叶片先在样本室CO2浓度400 μmol/mol、叶室温度(26±2)℃、叶室空气相对湿度(60±5)%和光强1 200 μmol/(m2·s)等条件下,诱导30 min以上,直到光合速率和气孔导度相对稳定。其他环境条件不变,采用Ciras-3内置自动程序将光强梯度设置为1 400、1 200、1 000、800、600、400、300、200、150、100、50、20、0 μmol/(m2·s)(蓝光10%),依次测定叶片气体交换数据。每处理重复测量3株。从光响应曲线中获取1 200 μmol/(m2·s)下的光合参数:净光合速率(Pn)、胞间CO2浓度(Ci)、蒸腾速率(E)、气孔导度(Gs)、水分利用效率(WUE=Pn/E)。
1.3.2 直角双曲线修正模型
采用直角双曲线修正模型[17]对不同处理下龙葵叶片的光响应曲线进行拟合,获得最大净光合速率(Pnmax)、初始量子效率(AQY)、光补偿点(LCP)、光饱和点(LSP)和光下暗呼吸速率(Rd)等光响应参数,其表达式如下:
令Pn=0,计算的PAR值为LCP。
1.3.3 生长指标和生物量
气体交换参数测定结束后采集叶片并测定其比叶重(SLA)。于2019年8月8号试验结束取样时,采用卷尺测量植株株高和冠幅。将植株分为根系和地上部分,烘干至恒重后称取干重,计算根系、地上部分及单株的生物量和根冠比。植株地上部分粉碎后,采用凯氏定氮法测定地上部分氮含量(Nmass)[18]。每个处理重复测量3株。
1.4 数据处理
光响应曲线的拟合基于光合计算平台(http://photosynthetic.sinaapp.com/)进行。采用Microsoft Excel 2016和R语言(R-3.5.1)进行数据整理、分析和作图。采用SPSS 17.0进行方差分析和多重比较(Turkey HSD)。
2 结果与分析
2.1 施氮对镉胁迫下龙葵生长和生物量的影响
由图1可知,不同处理间株高和冠幅均存在显著差异(P<0.05)。与CK相比,CdN0处理龙葵株高和冠幅无显著差异,但冠幅有下降趋势。与CdN0相比,CdN150、CdN300、CdN450龙葵株高分别增大29.4%、43.0%和42.4%,冠幅分别增大56.8%、110.7%和122.1%(P<0.05)。龙葵株高和冠幅在CdN300与CdN450处理间差异不显著。

图1 施氮对镉胁迫下龙葵生长指标的影响
由表1可知,龙葵根系生物量、地上部分生物量、单株生物量、根冠比、比叶重和Nmass在不同处理间均存在显著差异。CdN0处理下龙葵根系生物量、单株生物量和比叶重均显著小于CK;与CdN0相比,CdN150、CdN300、CdN450龙葵根系生物量分别增大86.7%、116.7%和106.7%,地上部分生物量分别增大137.3%、209.6%和266.3%,单株生物量分别增大123.9%、185.0%和223.9%(P<0.05)。CdN450龙葵根冠比显著小于CdN0,CdN450龙葵Nmass显著大于CdN0(P<0.05),CdN450龙葵单株生物量、根系生物量、根冠比、比叶重、Nmass与CdN300无显著差异。

表1 施氮对镉胁迫下龙葵生物量的影响
2.2 施氮对镉胁迫下龙葵光响应曲线的影响
如图2所示,当光强较低时[PAR≤200 μmol/(m2·s)],不同处理下龙葵叶片Pn随PAR的增加而迅速增大,Pn对PAR的变化非常敏感。随PAR的增强,不同处理下龙葵叶片Pn表现出不同的响应规律。当200 μmol/(m2·s)
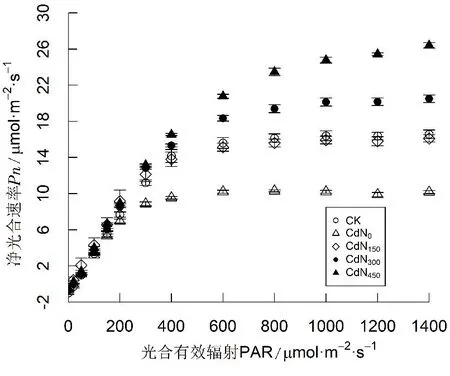
图2 施氮对镉胁迫下龙葵光响应曲线影响
由表2可知,不同处理间龙葵叶片Pnmax和LSP存在显著差异,AQY、Rd和LCP无显著差异。Cd胁迫下龙葵Pnmax和LSP均显著小于CK。与CdN0相比,CdN150、CdN300、CdN450龙葵Pnmax分别增大52.9%、93.9%和144.4%,LSP分别增大23.5%、30.9%和58.5%(P<0.05)。CdN450龙葵Pnmax和LSP显著大于CdN300。

表2 施氮对镉胁迫下龙葵光响应特性的影响
2.3 施氮对镉胁迫下龙葵气体交换参数的影响
由表3可知,不同处理间龙葵叶片Pn、Gs、Ci和E存在显著差异,WUE无显著差异。Cd胁迫下龙葵Pn显著小于CK。与CK相比,Cd胁迫下龙葵Gs和E无显著变化,但均有下降趋势。与CdN0相比,CdN150、CdN300、CdN450龙葵Gs分别增大58.3%、101.6%和159.7%,E分别增大74.6%、106.4%和116.9%(P<0.05)。CdN300和CdN450龙葵Gs分别比CdN0大212.5%和262.3%(P<0.05)。CdN450龙葵Ci显著小于CdN0和CdN300。CdN450龙葵Gs显著大于CdN300。
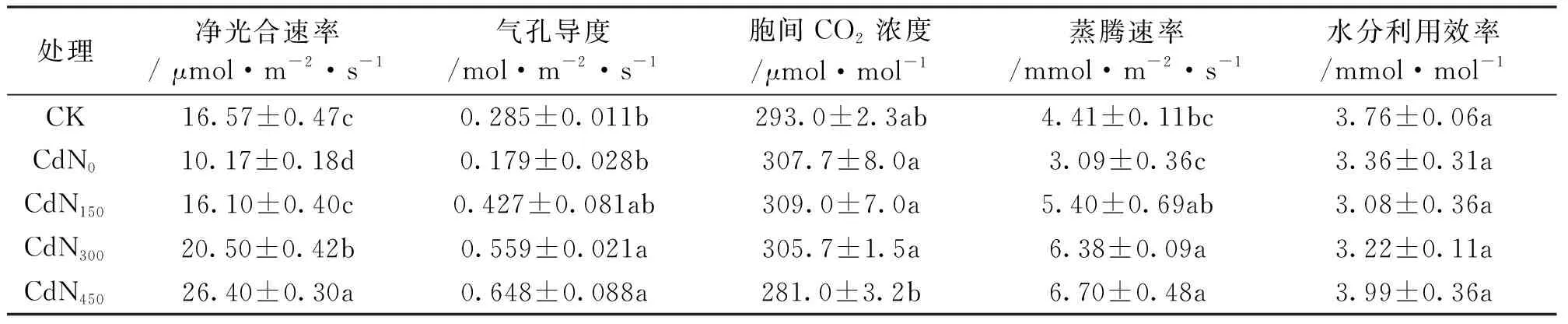
表3 施氮对镉胁迫下龙葵叶片气体交换参数的影响
2.4 参数间相关性分析
由表4可知,Cd胁迫及不同施氮处理下龙葵叶片Nmass与Pnmax(r2=0.78,P<0.01)、Gs(r2=0.60,P<0.05)、根系生物量(r2=0.58,P<0.05)、地上部分生物量(r2=0.82,P<0.01)、单株生物量(r2=0.80,P<0.01)呈显著正相关关系,Pnmax、Gs分别与地上部分生物量(Pnmax:r2=0.90,P<0.01;Gs:r2=0.91,P<0.01)和根系生物量(Pnmax:r2=0.80,P<0.01;Gs:r2=0.77,P<0.01)、单株生物量(Pnmax:r2=0.90,P<0.01;Gs:r2=0.91,P<0.01)呈显著正相关,Rd与根系、地上部分、单株生物量相关性不显著。
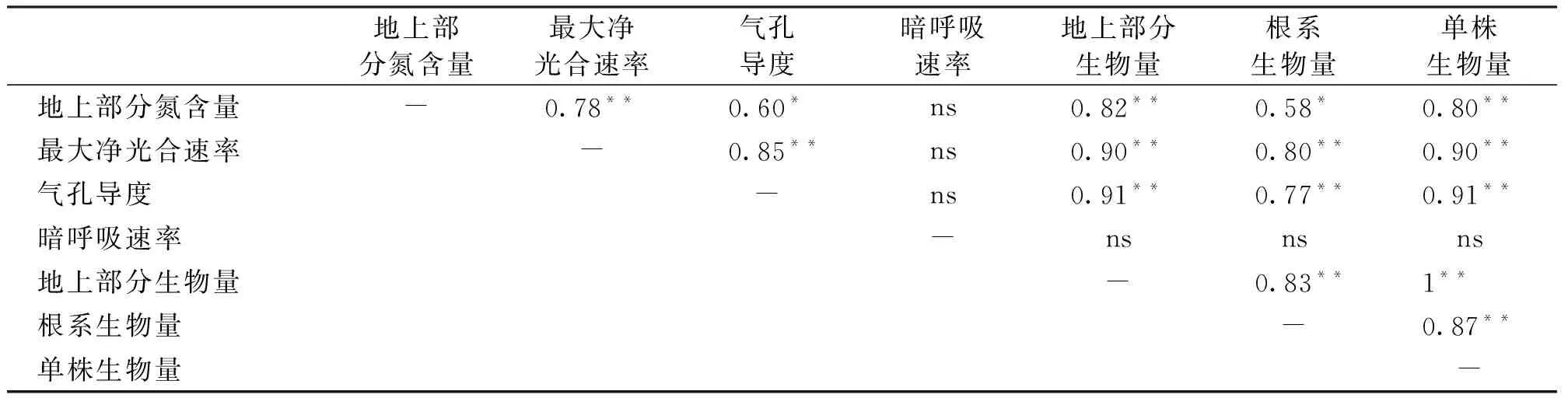
表4 研究参数间相关性分析
3 讨论与结论
光合作用是植物生长发育的基础。Cd胁迫导致植物光合作用下降的原因可分为气孔因素和非气孔因素,Gs和Ci同时减小则表明气孔因素限制占主导地位,若Gs减小,Ci增大则表明非气孔因素限制占主导地位[2]。本研究发现Cd胁迫下龙葵植株叶片Pn和Gs减小,Ci增大,表明光合限制中非气孔因素限制占主导地位,这与甜瓜(Cucumismelo)的研究结果一致[19]。Gs可以反映气孔导度对光合作用的限制。Cd胁迫下龙葵Gs下降说明气孔导度对光合作用的限制作用增大,这与南瓜(Cucurbitamoschata)[20]和番茄(Lycopersiconesculentum)[21]等植物的研究结果类似。Cd胁迫下Gs的下降可能与气孔密度、气孔大小的变化有关[22-23]。Yan等[24]发现施N可以增大马铃薯(Solanumtuberosum)气孔密度和气孔尺寸,进而增大Gs。本研究发现Cd胁迫下施N后龙葵Gs增大,说明施N有助于减小Cd胁迫下龙葵气孔导度对光合作用的限制,这与结缕草(Zoysiajaponica)[12]和小麦(Triticumaestivum)[25]等植物的研究结果类似。
光响应参数可以反映植物在逆境条件下的光合潜能和光能利用能力。其中,Pnmax反映了植物在大气CO2浓度下的最大光合能力,其值越大则植物叶片光合潜能越大。本研究发现Cd胁迫下龙葵Pnmax显著下降,说明Cd胁迫导致龙葵光合潜能下降,这与烟草(Nicotianatabacum)[26]和互叶醉鱼草(Buddlejaalternifolia)[27]等植物的研究结果一致。有研究发现Cd胁迫会导致植物叶绿素含量、PSII量子转换效率、电子传递速率、气孔导度、叶肉导度[28-29]等降低,这可能导致Pnmax降低。N对植物光合作用有显著的调节作用,施N可以增加叶片N素含量,提高叶片光合能力,进而增大植株生物量[8,30]。本研究发现Cd胁迫下施N后龙葵Nmass和Pnmax显著增大,且Pnmax与Nmass呈显著正相关,说明施N对Cd胁迫下龙葵光合能力有显著提升作用,与南方四季杨(Populusdeltoides×P.nigra)[31]、巨桉(Eucalyptusgrandis)[32]、小麦[25]等植物的研究结果一致,这可能与N素作用下植物叶绿素含量、电子传递速率和Gs等的增大有关[12-13]。LCP和LSP分别反映了叶片对弱光和强光的利用能力。本研究表明Cd胁迫下龙葵叶片LCP和LSP呈下降趋势,表明植株叶片利用弱光的能力有所增强,而利用强光的能力有所减弱,这与檫木(Sassafrastzumu)[2]的研究结果类似。有研究表明,N肥能显著提高黑麦草(Loliummultiflorum)LSP,增大其光能利用区间,促进植株生长[33]。本研究表明Cd胁迫下随施N水平的增大,龙葵LCP先升高后降低,LSP显著升高,说明其对弱光的利用能力先减弱后增强,而对强光的利用能力持续增强,光能利用区间增大,有利于植株生长。
Cd胁迫和施N对叶片光合作用的影响会反映到植物生物量累积上[34-35]。本研究发现Cd胁迫降低龙葵光合能力而抑制其根系与地上部分生长和单株生物量积累,这与烟草[36]和吊兰(Chlorophytumcomosum)[29]等植物的研究结果类似。古洪双等[31]研究表明施N可以减轻Cd对南方四季杨根、茎、叶生物量积累的抑制。毕景文等[7]认为植株生物量积累的增大对Cd有稀释效应,降低植株体内Cd的生理浓度,增强植物的耐Cd能力。本研究中Cd胁迫下施N后植株株高、冠幅、地上部分生物量、根系生物量和单株生物量均显著增大,说明N肥对Cd胁迫下龙葵的生长及生物量累积有显著的促进作用。其中,施N后龙葵根冠比显著降低,说明Cd胁迫下施N对龙葵地上部分生长的促进作用大于根系。这表明施N使得植物将营养物质更多地分配到地上部分,尤其是叶片,以更大程度地提高其植株光合效率[34]。
在本试验条件下,CdN300与CdN450处理间龙葵株高、冠幅和单株生物量差异不显著,但CdN450处理下龙葵地上部分生物量、净光合速率和最大净光合速率均显著高于CdN300处理,说明高N(CdN450)水平较中N(CdN300)水平更有利于促进龙葵地上部分生长,该水平下龙葵仍具有一定的生长潜力。综合来看,450 mg/kg CO(NH2)2施用量对Cd胁迫下龙葵光合速率和生物量积累的促进作用最大,是最适宜的施N水平。本试验中高水平施N量对Cd胁迫下龙葵生长的促进作用最大,而更高水平的施N量是否对Cd胁迫下龙葵生长具有更好的促进作用有待于进一步研究。
