甜角大小孢子发生和雌雄配子体发育及受精过程
2021-12-04夏富娴李梦雪李体初杨时宇王兵益
夏富娴,李梦雪,李体初,杨时宇,王兵益**
(1.中国林业科学研究院 资源昆虫研究所,云南 昆明 650224;2.南京林业大学,江苏 南京 210037)
甜角(Tamarindus indicaL.)又名罗望子、酸豆、酸梅,豆科(Leguminosae)酸豆属(TamarindusL.)的热带亚热带常绿大乔木,原产地为非洲,后传至其它大洲[1].甜角是单种属植物,包括两种果实类型,甜型和酸型.酸型甜角很早以前就被引种到中国,主要分布于云南、海南、广州、广西、福建、台湾等地;甜型是20世纪末才从泰国引种到国内,主要栽培区域在云南南部及干热河谷地区[2].甜角果实是特色热带水果,含多种营养物质,包括多种有机酸、糖类、维生素、蛋白质、游离氨基酸和矿质元素等,鲜食生津止渴,清热解暑,消食化积,可加工成果脯、蜜饯、饮品等[3].种子中富含罗望子多糖,可制成罗望子胶,是一种优质的食品添加剂[4].所以,甜角具有良好的综合经济利用价值和前景.
甜角的经济价值主要体现在其果实上,然而,作为新引进物种,甜角在种植过程中一直存在开花量大,但结实率和果实产量低的问题.基于课题组之前调查统计,甜角自然结实率仅为7%[5].成功的有性生殖过程保证了植物的结实,很多学者认为,胚胎发育进程受到阻碍是植物生殖失败的重要原因之一[6].目前,国内外甜角的研究主要集中在果实营养、化学成分分析、药理等方面[7-9].在胚胎学方面的研究还鲜有报道. 为此,本研究采用石蜡切片法和荧光显微观察法对甜角大小孢子发生和雌雄配子体发育及受精过程进行观察,旨在阐明甜角大小孢子发生和雌雄配子体发育过程,为甜角生殖生物学积累资料,并为了解其结实率低的原因和解决该问题提供细胞学证据,以期为后续的丰产措施提供理论基础.
1 试验地概况
实验所用材料为引种自泰国,种植于云南省昆明市中国林业科学研究院资源昆虫研究所元江试验站内(102°22′E,23°55′N,海拔390~420 m)22年生的甜角植株.试验站区属于北热带干热河谷气候类型,年平均温度12~24℃,最冷月平均气温7~17℃,最热月平均气温16~29℃,年平均降水量770~2 400 mm,土壤为砂红壤,呈微酸性至中性.
2 材料与方法
2.1 组织切片选取植株中不同发育阶段的花蕾和花朵,保存于FAA固定液中(V(甲醛)∶V(冰乙酸)∶V(70%乙醇)=1∶1∶18)固定24 h,4℃冰箱中保存备用.采用常规石蜡切片法制片[10]. 利用Leica RM轮式切片机切片,切片厚度为8~10μm,经番红固绿(Safranin-Fast Green)染色法染色,中性树胶封片后用光学显微镜观察并拍照.
2.2 花粉粒形态观察采集甜角花朵,用解剖针轻轻拨动花药壁,使花粉粒散落到导电胶上,离子溅射仪(JS-1600)喷金镀膜后,用TM3000扫描电镜进行观察并拍照.
2.3 甜角受精作用花粉管生长动态观察在开花前1 d对即将开放的200个花蕾去雄套袋并进行挂牌标记,于开花当天10:30—11:00进行授粉.于授粉后1、2、3、4、5、6、7、8、9、20、24 h分别采集雌蕊各10个样本放于卡诺固定液(V(无水乙醇)∶V(冰醋酸)=3∶1)中固定并带回实验室,用2 mol·L-1的NaOH溶液进行透明和软化3 h,再用苯胺蓝溶液(质量分数0.1%)染色1 h后压片,用OLYMPUS LX71型荧光显微镜观察花粉的萌发和花粉管的生长并拍照.
3 结果与分析
3.1 花药壁的发育甜角花两性,每朵花有3枚雄蕊,雌蕊1枚(图1(a)~(c)).切片结果显示花药有4个花粉囊,呈左右不对称型并排排列,相邻两个花粉囊之间被药隔分离(图2(b)).花药发育初期的结构比较简单,由外侧的表皮和内侧的分生组织两部分构成,随着花药的发育,在花药的4个角隅处有一群体积较大、细胞质较浓、细胞核明显的细胞,这就是孢原细胞(图2(a)).孢原细胞经过1次平周分裂后形成初生壁细胞和初生造孢细胞.初生壁细胞经1次平周分裂和多次垂周分裂后形成内外2层细胞,随着进一步发育内层细胞形成绒毡层细胞,外层细胞形成次生壁细胞.次生壁细胞随后发育形成花药的中层和药室内壁.5层花药壁形成,由外向内依次是表皮、药室内壁、中层(2层)和绒毡层(图2(j)).观察结果显示,甜角花药壁的发育类型属于基本型.

图1 甜角花部结构Fig.1 The floral structure of T.indica

图2 甜角小孢子发生和雄配子体发育Fig.2 The microsporogensis and development of male gametophyte of T.indica
在小孢子母细胞形成的时期,表皮细胞进行分裂并液泡化,这一时期细胞特征明显,其径向壁延长,垂周壁加厚呈长方形状,并且细胞排列紧密,核大(图2(h)).至后期小孢子细胞成熟后,部分表皮细胞逐渐解体,仅留下波浪状的角质层细胞(图2(j)).观察发现,甜角的绒毡层属于变形绒毡层,当小孢子母细胞处于减数分裂的过程中时绒毡层的细胞从花药壁脱离,成团移至花药腔内(图2(g)).当小孢子成熟时,同侧的2个花粉囊连通,在2个花粉囊相连处的药室壁渐渐变薄裂开,花粉散出(图2(k))
3.2 小孢子发生和雄配子体形成在花药壁发育的时候,小孢子同步发生,初生造孢细胞经过数次的有丝分裂,形成次生造孢细胞.这一时期的次生造孢细胞体积大,排列紧密,细胞质浓厚,形状多呈多边形.其进一步发育后形成小孢子母细胞,初期的小孢子母细胞排列紧密,发育到一定程度后因为胼胝质的沉积会使它们彼此分离开形成椭圆形、细胞核大、核仁明显、细胞质浓厚的单个游离小孢子母细胞(图2(c),(d)).随后,游离的小孢子母细胞进入减数分裂期,第1次减数分裂后形成2个子核,但这个时期并没有细胞壁的形成,细胞直接进入第2次减数分裂形成4核,这一时期因为彼此间有胼胝质的相互隔离,形成四分体. 甜角小孢子四分体的排列方式以四面体型为主,偶见左右对称型(图2(e)),胞质分裂为同时型.四分体存在的时间较短,发育至后期,彼此间的胼胝质逐渐解体,形成4个体积小,形状不规则的游离小孢子,在此期间,绒毡层细胞逐渐解体(图2(f)).随后,小孢子细胞进一步发育形成一个体积较大圆形有中央大液泡的细胞.在此期间,中央大液泡会将细胞核挤向细胞壁的一侧,此时进入单核靠边期;单核靠边期的小孢子细胞的细胞核进行一次不均等的有丝分裂,形成一个圆的较大的营养核和一个小的生殖核.之后,细胞内营养物质不断积累,中央大液泡逐渐消失,形成2-细胞型的成熟花粉粒,此时观察到绒毡层细胞并未完全降解(图2(i)).
通过对甜角小孢子母细胞发育过程的观察,发现同一花药不同药室间以及同一药室内的小孢子母细胞发育进程具有不同步现象(图2(h)).在甜角雄配子体的发育过程中,观察到小孢子在发育过程中出现细胞核降解、细胞质浓缩及细胞皱缩的败育现象(图2(i)).在观察中还发现很多花粉囊中花粉粒数目很少,这部分花粉囊中单个花药腔内约只有8~10粒花粉(图2(k)),此外,还观察到部分花粉囊为空室(图2(l)).
3.3 花粉形态观察通过扫描电镜观察,甜角花粉为单粒花粉(图3(a)). 赤道面观为长球形(图3(b)),极面观呈3裂钝三角形(图3(c)、(d)).于甜角的开花期随机选取若干花粉粒进行测量,并参照G.Erdtman对花粉形状类型的划分标准(P*E值表示花粉粒的大小,P代表极轴长,E代表赤道轴长;P/E值在1.33~2.0之间属于长球形,P/E值大于2.0属于超长球形)[11].结果显示甜角P为(49.77±1.24) μm,E为(33.32±0.98) μm,P*E值为1 658.34 μm2,P/E值为1.49,所以甜角花粉粒的形状是长球形或近长球形.萌发沟3裂延伸至两极、沟痕整体呈长条形,宽度1.8~2.4μm.花粉粒外壁瘤状纹饰紧密交错排列分布.

图3 甜角花粉形态扫描电镜观察Fig.3 Observation of pollen morphology by scanning electron microscope
甜角为单属种,甜角花粉形态的研究对酸豆属花粉形态特征具有代表性.所以将甜角花粉粒的P和E与豆科中其他5个属花粉特征资料进行了比较分析[12-14].发现,酸豆属花粉的P和E与其他5属相比较存在显著差异,且酸豆属P值明显大于其他5属(图4,不同属之间相同字母表示差异不显著p>0.05,不同字母表示差异显著p<0.05),说明同科不同属之间花粉形态差异较大.

图4 酸豆属与豆科其他属花粉极轴和赤道轴长比较Fig.4 Comparison of pollen polar and equatorial axis lengths between Tamarindus and other Leguminous genera
3.4 大孢子发生和雌配子体的形成甜角为单心皮雌蕊,子房一室,边缘胎座,厚珠心,胚珠弯生,着生在腹缝线的边缘上排成纵列.胚珠原基开始发育时,由薄壁组织细胞组成的珠心组织形成,近珠心基部形成内、外珠被原基,进而形成内、外双珠被(图5(a)).随后,伴随细胞的分裂生长在珠心表皮下形成一个体积大、原生质浓、有大细胞核与周围细胞差异明显的孢原细胞(图5(b)).孢原细胞进一步快速分裂发育形成一个体积更大、原生质更浓厚的大孢子母细胞(图5(c)).随着胚囊进一步发育大孢子母细胞分裂后形成4个大孢子.4个大孢子中只有靠近合点端的1个大孢子继续发育形成功能大孢子,其余3个大孢子退化消失.之后,功能大孢子生长膨大经有丝分裂后形成二核胚囊(图5(d),(e)).在此时期,二核胚囊的2个核因中央大液泡的作用逐渐移向胚囊的两端(图5(f)).二核胚囊再经过有丝分裂形成四核胚囊(图5(g)),四核胚囊继续进行分裂形成八核胚囊(图5(h)).
八核胚囊在较短的时间里继续生长发育形成细胞壁,此时胚囊发育成熟.观察发现,甜角胚囊发育为单孢子-蓼型胚囊,为比较原始的胚囊发育类型,是由1个功能大孢子发育形成7胞8核胚囊,包括珠孔端的2个助细胞和1个卵细胞、1个二倍体中央细胞和合点端的3个反足细胞.2个助细胞呈对称排列,具大液泡.卵细胞比助细胞小,易于辨认.二倍体的中央细胞液泡化明显(图5(i)).3个反足细胞体积较大,具2核.实验观察到甜角八核胚囊存在时间较短,胚囊成熟后不久3个反足细胞退化消失(图5(j));2个助细胞在受精前有开始退化解体的趋势,最后消失只留下卵细胞和中央细胞(图5(k)).观察中发现胚囊有败育现象(图5(l)).

图5 甜角大孢子发生和雌配子体发育Fig.5 The megasporogenesis and development of female gametophyte of T.indica
3.5 受精作用甜角花粉粒萌发时,在荧光染色下显示黄绿色荧光,随着花粉管的伸长,花柱内壁以及子房中也会呈现黄绿色荧光.授粉后30 min,柱头表面粘附大量花粉粒,但花粉粒未见明显变化(图6(a));授粉后1 h,部分花粉粒在柱头表面水合开始萌发,可观察到零星未穿过柱头表面的萌发的花粉管(图6(b));授粉后2 h,萌发的花粉粒数量明显增多,并有少量的花粉管穿过柱头表面的乳突细胞开始进入花柱,但荧光现象不强且主要集中在柱头处(图6(c));授粉后3 h,大量花粉粒已经萌发,荧光现象明显,许多花粉管穿过柱头表面沿引导组织开始生长(图6(d)),在花柱中可明显看到带有生殖核的花粉管(图6(e));授粉后4 h,柱头表面花粉粒基本已全部萌发,花粉管以束状形式向花柱基部延伸,少量花粉管已生长至花柱长度的3/4(图6(f));授粉后5 h,可观察到柱头表面几乎已无花粉粒萌发,花柱中花粉管的荧光宽度约为上一时间段的2倍,整个花柱内壁十分明亮(图6(g)),表明该时间段为花粉管伸长的最旺盛时期,此时,胚囊中中央细胞紧紧挨着卵细胞,卵细胞挨近中央细胞;授粉后6 h柱头和花柱中荧光现象开始减弱,花粉管离开引导组织,穿出隔膜进入子房腔内,沿着珠柄生长,伸入子房,从珠孔进入胚囊内部,释放出2个精细胞;然后,一个精细胞会向中央细胞靠近,另一个精细胞会向卵细胞靠近,精核紧贴卵细胞,其精核内容物会逐渐进入卵细胞内,此时精子与极核也完成融合,形成一个较大的细胞完成双受精过程,甜角的受精模式为珠孔受精(图6(h)).花柱中束状的花粉管缓慢呈现扭曲、断裂、不连续状态,柱头表面附着的花粉粒逐渐失去荧光效果,此状态为受精结束后花粉管解体的初期,甜角授粉12 h后花柱中仅可见极少量的、扭曲的花粉管,且花粉管解体、撤退的态势越发明显,柱头表面的花粉粒数量明显减少,此状态为受精结束后花粉管解体的末期(图6(i));授粉后24 h,柱头及花柱中的荧光消失,花粉管完全解体.
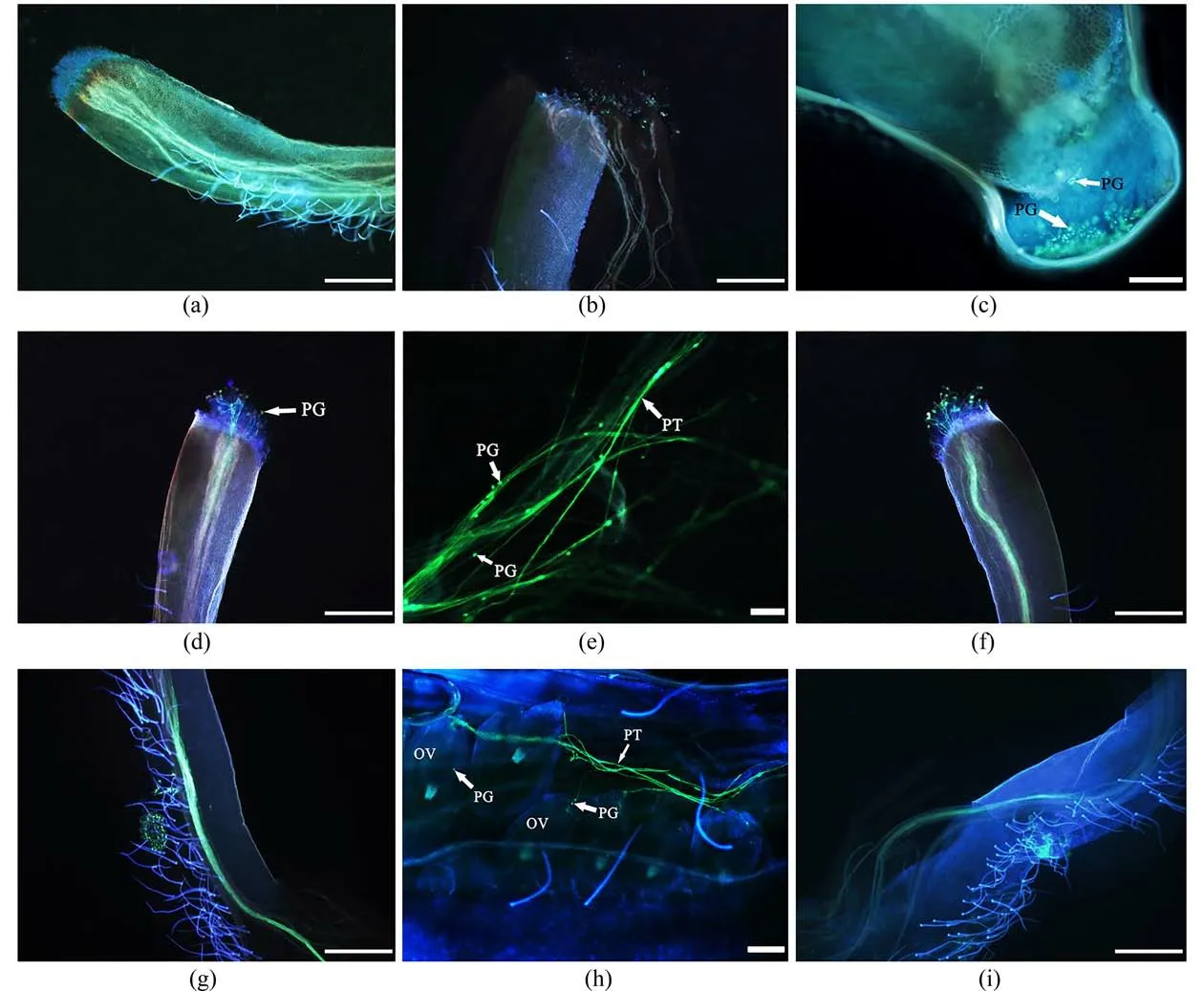
图6 甜角花粉萌发、和花粉管生长及受精过程Fig.6 Pollen germination, pollen tube growth and fertilization process of T.indica
4 讨论
植物胚胎学特征是植物系统进化过程中比较稳定的性状,是判定植物亲缘关系、系统进化进程的重要依据[15].比较酸豆属甜角与豆科中的豇豆属(VignaSavi)植物绿豆(V.radiata(L.)Wilczek)[16-17]和红小豆(V.angularis(Willd)Ohwi et Ohashi)[18],两型豆属(AmphicarpaeaElliot)的两型豆(A.edgeworthiiBenth.)[19],沙冬青属(AmmopiptanthusS.H.Cheng)的矮沙东青(A.mongolicus(Kom.)S.H.Cheng)[20],黄芪属(AstragalusL.)的黄芪(A.membranaceus(Fish.)Bunge)[21]的胚胎学特征,发现它们具备很多一致的特征,如花药具有4个花粉囊,花药壁由5层细胞构成,从外至内依次为1层表皮、1层药室内壁、2层中层和1层绒毡层,成熟花粉粒为2-细胞花粉,子房一室,边缘胎座,双珠被,厚珠心,胚珠弯生,胚珠多数,单孢原直接发育为大孢子母细胞,胚囊发育为8个时期,即孢原细胞形成期、大孢子母细胞形成期、大孢子母细胞减数分裂期、功能大孢子形成期、单核胚囊形成期、胚囊有丝分裂期、8核胚囊发育期和成熟胚囊期,胚囊发育类型为蓼型等.比较研究反映了豆科各属种之间系统发育上的一致性,说明他们有较近的亲缘关系.本研究认为酸豆属和上述几属的胚胎学特征虽然很多相一致,但在有些特征方面也有所不同.例如:甜角小孢子四分体多为四面体型,偶尔可见左右对称型,而上述其它植物四分体都为四面体型;经观察发现甜角花药壁两中层分别自内、外侧次生周缘细胞平周分裂形成的两层细胞,其发育类型为基本型,但黄芪药壁发育类型为单子叶型,红小豆为双子叶型. 其次,甜角小孢子胞质分裂是同时型,两型豆兼具同时型和连续型两种类型. 研究认为2-细胞花粉,双珠被,厚珠心,蓼型胚囊发育是比较原始的胚胎学发育特征,弯生胚珠和直生胚珠要比倒生胚珠进化[20].说明甜角在胚囊的形态发育中具有较明显的原始特征,但胚珠的发育特征比较进化.甜角既具有相对原始的特征,又具有相对进化的特征,镶嵌进化或异步演化明显,我们认为酸豆属以及豆科在被子植物中属于相对较原始的类群.
花粉是遗传信息的载体,具有较强的遗传保守性,其固有的形态特征在科与属之间对植物的系统分类地位具有一定实际意义[22].甜角为单属种,对其进行花粉形态特征的研究填补了酸豆属花粉形态学资料的空白和可为其准确分类提供重要依据.本研究中,甜角与豆科其他属的花粉形态特征有明显的差异性.甜角花粉外壁有瘤状纹饰紧密排列,沙冬青属、刺桐属(ErythrinaL.)、紫荆属(CercisL.)、皂荚属(GleditsiaL.)、槐属(SophoraL.)外壁都是网状纹饰,且沙冬青属花粉外壁具有多角形或圆形网眼.所以从孢粉学的观点看将甜角单独归为酸豆属是合理的.
甜角在减数分裂过程中,同一花药不同药室间以及同一药室内的小孢子母细胞发育具有不同步现象.这种现象在很多植物中都有发现,例如柿品种‘禅 寺 丸’(Diospyros kaki‘Zenjimaru’)[23]、楸 树(Catalpa bungeiC.A.Meyer)[24]、车桑子(Dodonaea viscosa(L.)Jacq.)[25]、‘早 籽 银 桂’(Osmanthus fragrans(Thunb.)Lour.‘Zao Zi Yin’)[26]、‘晚银桂’(Osmanthus fragrans(Thunb.)Lour.‘Wan Yin’)[27]等中也有出现.这种不同步现象使花粉散播的总时间延长,从而使传粉和受精的时间也有所延长的一种适应机制,可能是植物长期适应自然环境而形成的一种生殖策略.
成功的有性生殖过程保证了植物群体的结实,从而保证了植物群体的扩大和经济效益的提高.目前研究表明,胚胎发育进程受到阻碍是植物生殖失败的重要原因之一[28].本研究观察到甜角有空花粉囊现象,空花粉囊的出现可能是由于雄配子体在其生长环节出现异常状况,无法发育成成熟花粉所致.绒毡层在花粉发育的过程中起着重要的作用,它会随着花粉的发育通过溶解自身为花粉发育提供所需要的营养物质.绒毡层细胞无法正常生长、过早退化或后期退化不完全,都会使小孢子母细胞的发育出现异常,导致花粉败育[29].研究表明,毛白杨(Populus tomentosaCarrière)绒毡层细胞Ca2+-ATPase的分布异常会影响绒毡层细胞的正常PCD(programmed cell death)进程,使绒毡层细胞无法正常降解,导致花粉的败育;拟南芥(Arabidopsis thaliana(L.)Heynh.)在高温环境下,绒毡层PCD进程会出现滞后现象,因绒毡层后期无法彻底降解,导致不能产生正常的花粉粒[6,30].我们的研究发现,甜角花粉发育过程中小孢子在发育过程中出现细胞核降解、细胞质浓缩及细胞皱缩的败育现象.还观察到2-细胞花粉形成时期绒毡层出现降解不完全现象.因此推断,甜角胚胎发育过程中可能绒毡层生长发育异常,降解滞后使花粉发育过程中不能吸收足够的营养物质形成成熟的花粉粒,造成了花粉败育现象,从而降低了花粉数量,影响其后期的传粉生殖过程,推断这可能是甜角多花少实的原因之一.此外,本研究中甜角生长于干湿季分明的干热河谷地区,气候特殊,夏季高温干旱且漫长.因此,我们推测甜角绒毡层的生长发育异常,可能与其生长环境因素有关,这还有待于进一步研究.这将为干热河谷地区甜角的进一步开发利用提供一定的理论依据.
受精是植物生殖生长的重要过程,它不仅是植物界有性生殖最进化的形式,也是保证植物生殖成功的关键.植物的受精作用是一个动态的变化过程,每种植物均有不同的特性.用荧光显微镜是观察植物从花粉落到柱头开始萌发到花粉管生长伸长到达子房进入胚囊完成受精情况最直观和最有效的方式.本研究中花粉粒黏附在柱头上1 h后能正常开始萌发,授粉6 h后,花粉管生长延伸到达子房从珠孔进入胚囊完成受精.之前研究认为,胚囊中的助细胞对花粉管生长延伸至子房具有主要的定向指引作用,因为助细胞分泌的多肽LURES对花粉管具有吸引作用[31].本研究中观察到在受精之前2个助细胞退化消失.何孟元等[32]对豆科植物大豆(Glycine max(L.) Merr.)的胚胎学研究也发现受精前成熟胚囊中2个助细胞已经退化消失,有时只有1个退化,极少数出现2个助细胞都未退化的现象.杨维才等[33]对拟南芥的研究发现中央细胞中CCG(central cell guidance)基因的特异性表达是将花粉管从柱头导向进入胚囊所必需的,研究还认为中央细胞可通过助细胞并协同助细胞一起完成对花粉管的导向作用.甜角雌配子体发育后期两个助细胞退化,只留下中央细胞和卵细胞,推测其中央细胞在甜角受精过程中对花粉管的导向性具有重要作用.关于甜角具体的受精机制还有待进一步研究.
