呼吸代谢参与粉红单端孢侵染厚皮甜瓜果实不同阶段的防御反应
2021-12-02薛素琳南米娜DovPRUSKY薛华丽
薛素琳,南米娜,龚 迪,王 斌,姜 红,Dov PRUSKY,3,毕 阳,*,薛华丽
(1.甘肃农业大学食品科学与工程学院,甘肃 兰州 730070;2.甘肃农业大学理学院,甘肃 兰州 730070;3.以色列农业研究组织农产品采后科学部,以色列 里雄莱锡安 7505101)
粉红单端孢(Trichothecium roseum)是主要的采后病原真菌[1],不仅侵染杏[2]、甜瓜[3]和番茄[4]等多种果蔬,引起品质劣变,而且还会在果实体内积累单端孢霉烯族毒素,造成潜在的安全隐患[5]。该病原菌主要经由果实表面的伤口和自然孔口侵染[6],通过碱化果实伤口环境激活胞外酶致病[7]。有报道表明,果蔬的呼吸代谢在病原菌侵染过程中具有重要作用[8]。三羧酸(tricarboxylic acid,TCA)循环是重要的呼吸代谢途径,在提供能量、还原力以及碳骨架方面具有核心作用[9],磷酸戊糖途径(pentose phosphate pathway,PPP)是另一条重要的呼吸代谢途径,除了提供还原力外,还是核苷酸、芳香族氨基酸、苯丙烷及其衍生物碳骨架的重要来源[10]。Zhang Shen等发现,焦腐病菌(Lasiodiplodia theobromae)侵染激活了龙眼果实的TCA循环,导致还原型烟酰胺腺嘌呤二核苷酸(reduced nicotinamide adenine dinucleotide,NADH)显著积累,而PPP受到抑制,烟酰胺腺嘌呤二核苷酸磷酸(reduced nicotinamide adenine dinucleotide phosphate,NADPH)含量减少[11];在拟茎点霉菌(Phomopsis longanae)侵染龙眼果实期间也观察到类似的现象[12]。也有报道表明,灰霉病菌(Botrytis cinerea)侵染活化了苹果果实的PPP,促进了NADPH的合成[13]。同样,扩展青霉(Penicillium expansum)侵染苹果果实后激活了葡萄糖-6-磷酸脱氢酶(glucose-6-phosphate dehydrogenase,G6PDH),促进了NADPH合成[14]。尽管已有不同病原真菌侵染增强果实TCA循环和PPP的报道,但T. roseum侵染厚皮甜瓜后,果实TCA循环和PPP如何发生变化及其在不同侵染阶段的差异鲜见报道。因此,本实验用T. roseum接种厚皮甜瓜果实,观察常温条件下果实病斑直径的变化,测定侵染期间果实病健交界处组织TCA循环和PPP关键酶活力以及中间产物含量的变化,分析不同侵染阶段的差异,以期为揭示TCA循环和PPP在病原菌侵染果实中的作用提供理论依据。
1 材料与方法
1.1 材料、菌株与试剂
‘玉金香’厚皮甜瓜,于2017年6月14日(花后35 d)采自甘肃省兰州市皋兰县什川镇泥湾村。单果套发泡网袋后入包装箱(30 个/箱),当天运抵实验室,于(22±2)℃、相对湿度55%~60%下贮藏待用。
T. roseum为本实验室保存菌种。
烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NAD)、烟酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate,NADP)美国西格玛公司;植物NAD激酶(NAD kinase,NADK)、植物核酮糖-5-磷酸(ribulose-5-phosphate,Ru5P)酶联免疫分析试剂盒 上海杏宜生物科技有限公司。
1.2 仪器与设备
UV-2450型紫外分光光度计 日本岛津公司;3K30型高速冷冻离心机 北京东讯天地医疗仪器有限公司;CX21FS1C型生物显微镜 日本奥林巴斯工业有限公司;SW-CJ-2FD型超净工作台 苏净集团苏州安泰空气技术有限公司;LDZX-30KBS型立式压力蒸汽灭菌锅 上海申安医疗器械厂;SPX-30085H-II型恒温培养箱 上海新苗医疗器械制造有限公司。
1.3 方法
1.3.1 果实损伤接种
孢子悬浮液的制备参考Bi Yang等[15]的方法。取25 ℃下培养7 d含有T. roseum的PDA培养基,加入含体积分数0.05% Tween 20的无菌水约10 mL,用玻璃棒刮下PDA培养基上的T. roseum孢子,然后转入50 mL三角瓶中,在漩涡混合器上振荡15 s,再用双层纱布过滤,滤液用血球计数板计数算出孢子悬浮液的浓度,用无菌水稀释至1×106spores/mL。
果实损伤接种参考Bi Yang等[15]的方法并修改。果实先用自来水冲洗干净,然后用体积分数2%次氯酸钠溶液浸泡1 min,再用自来水冲洗后常温(22±2)℃下晾干。用体积分数70%乙醇溶液对果实接种点进行表面消毒后,用灭菌铁钉(直径3 mm)在果实赤道部位均匀打孔4 个(深3 mm)。孔内分别接入20 μL上述孢子悬浮液,以无菌水接种作对照组,晾干后于常温下贮藏。分别在接种后0、12、24、48 h和72 h测量病斑直径。每处理每一时间点用果实8 个,重复3 次。
1.3.2 生化指标测定取样
参照Lü Liang等[16]的方法,分别于接种后0、12、24、48 h和72 h用直径为10 mm的打孔器距离病斑边缘1 cm内打孔,切取皮下2~5 mm处果肉组织3 g,铝箔纸包裹,液氮速冻后于-80 ℃超低温冰箱中保存,用于测定生化指标。
1.3.3 相关代谢酶活力测定
1.3.3.1 异柠檬酸脱氢酶和苹果酸脱氢酶活力测定
参考Tang Mi等[17]的方法。取3 g冷冻组织,加入6 mL提取缓冲液(200 mmol/L Tris-HCl、10 mmol/L抗坏血酸、体积分数0.1% Triton X-100,pH 8.2)研磨。在4 ℃下匀浆,10 000 r/min离心30 min,上清液为粗酶液。
异柠檬酸脱氢酶(isocitrate dehydrogenase,IDH)活力测定反应体系:0.3 mL粗酶液、0.5 mL反应液(40 mmol/L羟乙基哌嗪乙硫磺酸、0.8 mmol/L NAD、0.2 mmol/L MnSO4,pH 8.2)、0.2 mL 2 mmol/L异柠檬酸钠,混合后于340 nm波长处测定吸光度,以每克鲜组织每分钟消耗1 mmol NAD表示1 个IDH活力,单位为mmol/(min·g)。
苹果酸脱氢酶(malate dehydrogenase,MDH)活力测定反应体系:0.3 mL粗酶液、0.5 mL反应液(40 mmol/L羟乙基哌嗪乙硫磺酸、0.8 mmol/L NAD、0.2 mmol/L MnSO4,pH 8.2)、0.2 mL 2 mmol/L苹果酸钠溶液,混合后于340 nm波长处测定吸光度。以每克鲜组织每分钟消耗1 mmol NAD表示1 个MDH活力,单位为mmol/(min·g)。
1.3.3.2 NADK活力测定
参照植物NADK酶联免疫分析试剂盒的方法。取3 g冷冻组织,加入6 mL磷酸盐缓冲液(pH 7.4)研磨。在4 ℃下匀浆3 000 r/min离心20 min,上清液为粗酶液。NADK活力测定反应体系:40 μL样品稀释液(0.01 mol/L磷酸盐缓冲液、体积分数0.5%牛血清白蛋白溶液,pH 7.2)、10 μL粗酶液、100 μL酶标试剂。37 ℃温育60 min,弃去液体,洗涤液清洗5 次,甩干后加入50 μL显色剂A、50 μL显色剂B混合,37 ℃避光15 min,加入50 μL终止液终止反应,于450 nm波长处测定吸光度。以每克鲜组织每分钟产生1 nmol NADP表示1 个NADK活力,单位为nmol/(min·g)。
1.3.3.3 G6PDH和6-磷酸葡萄糖酸脱氢酶活力测定
参考Mateos等[18]的方法。取3 g冷冻组织,加入6 mL预冷的提取缓冲液(100 mmol/L Tris-HCl、2.5 mmol/L MgCl2、1 mmol/L 乙二胺四乙酸、1 mmol/Lβ-巯基乙醇、5 g/100 mL聚乙烯吡咯烷酮,pH 8.0),在4 ℃条件下浸提20 min,匀浆液在10 000 r/min下离心20 min,上清液为粗酶液。
G6PDH活力测定反应体系:150 μL粗酶液、850 μL反应液(100 mmol/L Tris-HCl、10 mmol/L MgCl2、3 mmol/L NADP、3 mmol/L 葡萄糖-6-磷酸(glucose-6-phosphate,G6P),pH 8.0),混合后于340 nm波长处测定吸光度。以每克鲜组织每分钟产生1 nmol NADPH表示1 个G6PDH活力,单位为nmol/(min·g)。
6-磷酸葡萄糖酸脱氢酶(6-phosphogluconate dehy drogenase,6PGDH)活力测定反应体系:150 μL粗酶液、850 μL反应液(100 mmol/L Tris-HCl、10 mmol/L MgCl2、3 mmol/L NADP、3 mmol/L 6-磷酸葡萄糖酸(6-phosphogluconate,6PG),pH 8.0),混合后于340 nm波长处测定吸光度。以每克鲜组织每分钟产生1 nmol NADPH表示1 个6PGDH活力,单位为nmol/(min·g)。
1.3.3.4 核糖-5-磷酸异构酶活力测定
参考Xiong Yuqing等[19]的方法。取3 g冷冻组织,加入6 mL提取缓冲液(100 mmol/L Tris-HCl、1 mmol/L乙二胺四乙酸、1 mmol/Lβ-巯基乙醇,pH 7.4)研磨。在4 ℃下匀浆10 000 r/min离心5 min,上清液为粗酶液。核糖-5-磷酸异构酶(ribose-5-phosphate isomerase,RPI)活力测定反应体系包括0.1 mL粗酶液、0.3 mL提取缓冲液、0.1 mL 30 mmol/L核糖-5-磷酸,将试管短暂摇动并在室温下放置5~15 min。后依次加入0.5 mL 30 mmol/ L半胱氨酸,2 mL体积分数75%硫酸溶液和0.2 mL体积分数0.1%咔唑溶液,37 ℃下水浴30 min后,再在冰水中冷却2 min,540 nm波长处测定吸光度。以每克鲜组织每分钟消耗1 nmol核糖-5-磷酸表示1 个RPI活力,单位为nmol/(min·g)。
1.3.4 相关代谢产物含量测定
1.3.4.1 G6P和Ru5P含量的测定
G6P含量测定参考Elisa等[20]的方法。取3 g冷冻组织,加入6 mL提取缓冲液(0.2 mol/L Tris-HCl、体积分数0.05% Triton X-100,pH 7.5)研磨,在4 ℃条件下浸提10 min,10 000 r/min离心10 min,上清液为待测液。G6P含量测定反应体系:1 mL待测液、20 μL 30 mmol/L NADP、20 μL 0.5 mol/L MgCl2、10 μL 700 U/mL G6PDH,混合后于340 nm波长处测定吸光度。G6P含量单位为ng/g,结果以鲜质量计。
核酮糖-5-磷酸(ribulose-5-phosphate,Ru5P)含量通过酶联免疫吸附测定试剂盒测定。取3 g冷冻组织,加入5 mL生理盐水研磨,在4 ℃下3 000 r/min离心10 min,上清液为待测液。设置标准品孔和样品孔,标准品孔各加不同质量浓度(50、25、12.5、6.25、3.125 ng/mL)的50 μL标准品,样品孔加入10 μL待测液、40 μL样本稀释液(0.01 mol/L磷酸盐缓冲液、体积分数0.5%牛血清白蛋白,pH 7.2)。标准品孔和样品孔再加入100 μL辣根过氧化物酶标记的检测抗体混合,37 ℃水浴锅温育60 min,弃去液体,加入洗涤液,静置1 min,弃去洗涤液,如此重复5 次,加入试剂盒自带底物A、B各50 μL,37 ℃避光孵育15 min,加入50 μL终止液,混合后15 min内在450 nm波长处测定吸光度。Ru5P含量单位为ng/g,结果以鲜质量计。
1.3.4.2 NAD(H)和NADP(H)含量的测定
参考顾采琴等[21]的方法。取3 g冷冻组织,加入6 mL 0.1 mol/L HCl(对应NAD和NADP)或NaOH(对应NADH和NADPH)溶液研磨,在4 ℃条件下浸提10 min,10 000 r/min离心20 min,将上清液移至离心管中,用0.1 mol/L的NaOH(HCl)溶液调pH值至7.0,再在4 ℃条件下,10 000 r/min离心20 min,上清液为待测液。
NAD(H)和NADP(H)含量测定反应体系:待测液150 μL、200 μL混合液(含1 mol/L Tricine-NaH、40 mmol/L乙二胺四乙酸、4.2 mmol/L噻唑蓝、16.6 mmol/L吩嗪硫酸甲酯、25 mmol/L G6P)、150 μL 100 mmol/L NaCl,混合后37 ℃水浴5 min,取出后进行冰浴,加入100 μL G6PDH(对应NADP和NADPH)(乙醇脱氢酶(对应NAD和NADH)),再在37 ℃水浴40 min,最终加入200 μL 6 mol/L NaCl溶液终止反应,于570 nm波长处测定吸光度。NAD(H)和NADP(H)含量单位均为μmol/g,结果以鲜质量计。
1.4 数据统计与分析
上述各项测定至少重复3 次。全部数据用Excel 2010软件计算平均值和标准误,并用SPSS 19.0软件进行Duncan’s多重差异显著性分析(P<0.05表示差异显著)。
2 结果与分析
2.1 接种T. roseum后厚皮甜瓜果实病斑直径的变化
病斑直径增大是果实感病的直接表现。由图1可知,接种果实在接种后24 h内病斑直径无明显变化,接种后48 h接种果实与对照组病斑直径均增大,接种后72 h果实病斑直径显著高于对照组(P<0.05),是对照组的1.71 倍(图1)。
2.2 T. roseum接种对果实IDH、MDH、G6PDH和6PGDH活力的影响
IDH和MDH是TCA循环的关键酶,分别催化异柠檬酸和苹果酸生成α-酮戊二酸和草酰乙酸,同时合成NADH[22]。接种果实和对照组果实的IDH活力在接种后12 h迅速降低。接种果实的IDH活力之后缓慢上升,至48 h时达到峰值;对照组果实IDH活力在12 h后基本不变。接种果实的IDH活力在12 h时显著低于对照组,但在24 h后显著高于对照组(P<0.05),48 h时是对照组的1.63 倍(图2A)。接种果实的MDH活力在接种后呈现先升高后降低的趋势,对照组果实则是先迅速降低后缓慢升高。接种果实的MDH活力始终高于对照组(P<0.05),在12 h和24 h时分别是同期对照组的2.72 倍和2.14 倍(图2B)。
G6PDH和6PGDH是PPP的关键酶,分别催化G6P和6PG生成6-磷酸葡萄糖酸酯和Ru5P,同时调控NADP生成大量的NADPH以作为供氢体参与多种代谢反应[10]。接种果实和对照组果实的G6PDH活力在接种后24 h内相对平稳,接种果实的G6PDH活力显著高于对照组;24 h后接种果实的G6PDH活力迅速增加,在48 h和72 h时分别是同期对照组的1.83 倍和1.71 倍(P<0.05)(图2C)。接种果实和对照组果实的6PGDH活力在接种后迅速下降,接种果实的6PGDH活力在接种后24 h内显著低于对照组(P<0.05);但接种果实的6PGDH活力在接种24 h后迅速升高,48 h和72 h时分别是同期对照组的2.51 倍和2.98 倍(图2D)。
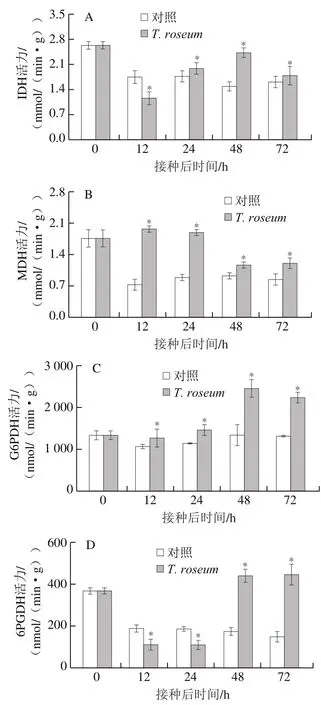
图2 T. roseum接种对果实IDH(A)、MDH(B)、G6PDH(C)和6PGDH(D)活力的影响Fig. 2 Effect of inoculation with T. roseum on the activity of IDH (A),MDH (B), G6PDH (C) and 6PGDH (D) in muskmelon fruit
2.3 T. roseum接种对厚皮甜瓜果实NAD(H)和NADP(H)含量的影响
NAD(H)和NADP(H)在能量形成和维持细胞的氧化还原平衡中具有核心作用[23]。在贮藏过程中,接种果实的NAD和NADH含量均先升高后降低,对照组果实这两种物质含量逐步升高,且接种果实的NAD和NADH含量总体显著高于对照组(P<0.05),在24 h时分别高出对照组50.37%和50.31%(图3A、B)。接种果实的NADP含量接种后48 h内基本稳定,但显著低于对照组(P<0.05);接种果实的NADP含量在48 h后含量迅速增加,在72 h时与对照组果实相比无显著差异;对照组果实的NADP含量逐渐升高,接种果实的NADPH含量在48 h内缓慢增加,且显著低于对照组,48 h后迅速增加,且在72 h时显著高于对照组(P<0.05)(图3C、D)。
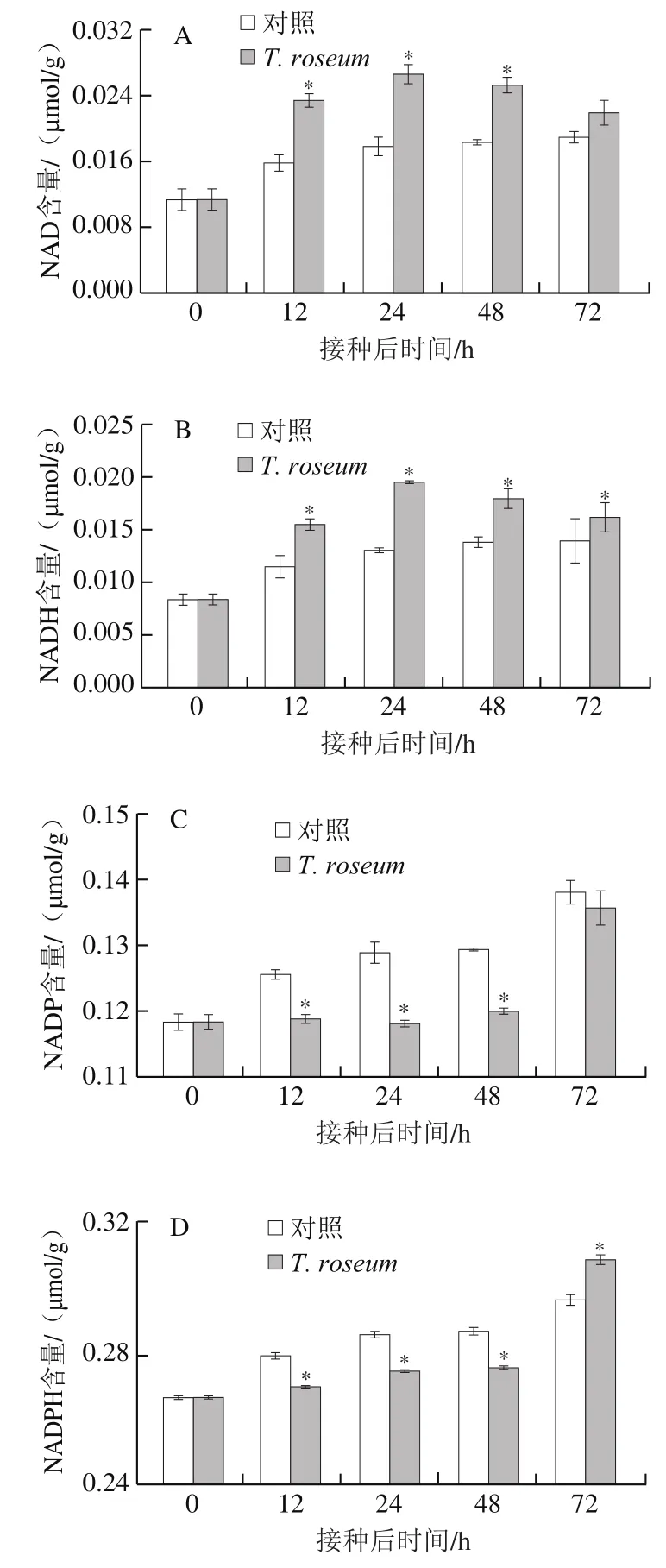
图3 T. roseum接种对果实NAD(A)、NADH(B)、NADP(C)和NADPH(D)含量的影响Fig. 3 Effect of inoculation with T. roseum on the contents of NAD (A),NADH (B), NADP (C) and NADPH (D) in fruits
2.4 T. roseum接种对厚皮甜瓜果实G6P和Ru5P含量的影响
G6P和Ru5P是PPP的重要中间产物,G6PDH催化G6P生成6-磷酸葡萄糖内酯,6PGDH催化6PG而产生Ru5P,这两个反应过程均伴有NADPH合成[24]。接种后果实的G6P含量呈现先略升高后降低再升高的趋势,虽然12 h和72 h时接种果实G6P含量高于对照组果实,但24 h和48 h时却显著低于对照组(P<0.05)(图4A)。接种果实和对照组果实的Ru5P含量在接种后48 h内逐渐降低,且接种果实的Ru5P含量显著低于对照组;接种果实Ru5P含量在接种48 h后有所升高,且在72 h时与对照组相比无显著差异(P>0.05)(图4B);说明接种显著降低了早中期果实Ru5P含量。
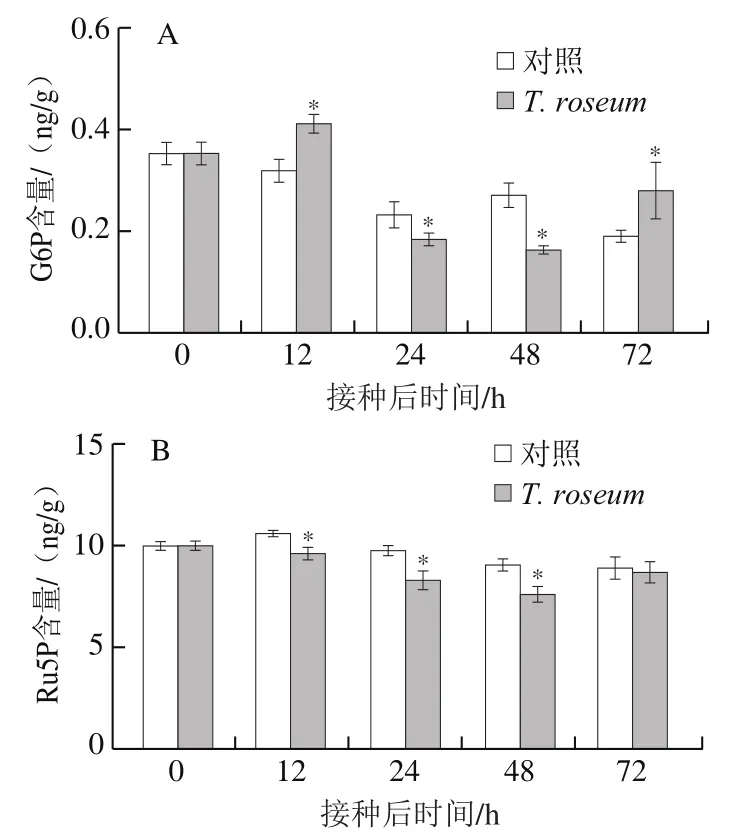
图4 T. roseum接种对果实G6P(A)和Ru5P(B)含量的影响Fig. 4 Effect of inoculation with T. roseum on the contents of G6P (A)and Ru5P (B) in muskmelon fruit
2.5 T. roseum接种对厚皮甜瓜果实NADK和RPI活力的影响
NADK通过催化NAD(H)与ATP或无机多聚磷酸提供的磷酰基生成NADP(H),维持生物体内的氧化还原平衡,保护组织免受氧化损伤[25]。接种果实的NADK活力在贮藏过程中先升高后降低,在48 h内显著高于对照组(P<0.05),12 h时是对照组的1.17 倍(图5A)。接种果实和对照组果实的RPI活力均呈先升高后降低的趋势,接种果实在48 h内显著低于对照组;48 h后接种果实的RPI活力有所上升,在72 h时与对照组相比无显著差异(P>0.05)(图5B);说明接种显著降低了早中期果实RPI活力。
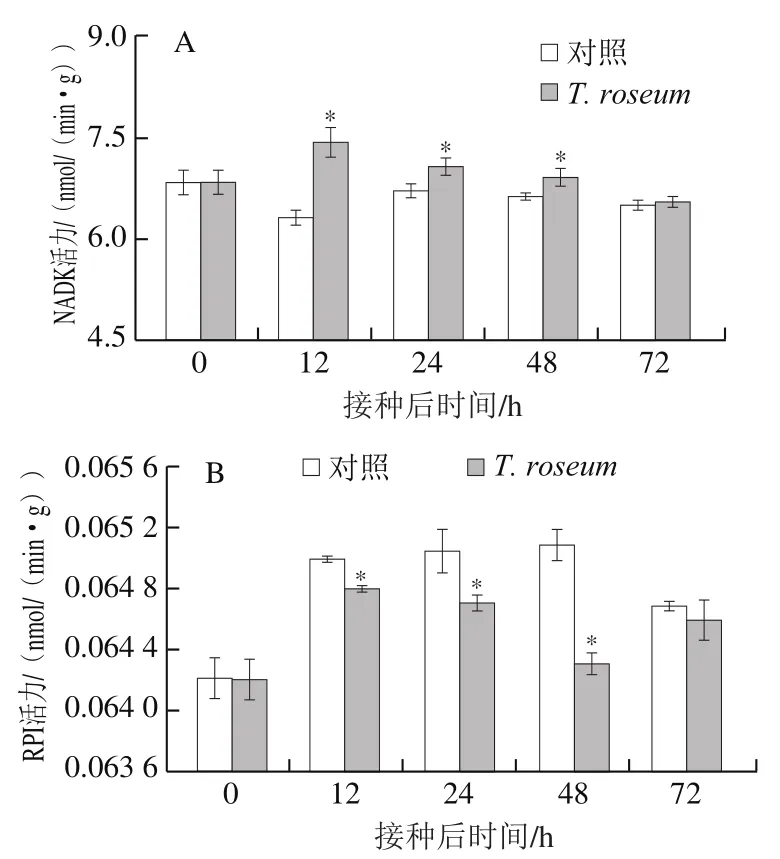
图5 T. roseum接种对果实NADK(A)和RPI(B)活力的影响Fig. 5 Effect of inoculation with T. roseum on the activity of NADK (A)and RPI (B) in muskmelon fruit
3 讨 论
呼吸代谢增强是病原真菌侵染植物后的典型反应[11]。但不同的呼吸途径在不同的侵染阶段表现各异,L. theobromae侵染龙眼早期诱导NAD(H)合成,促进TCA循环[11];P. longanae侵染荔枝果实后期会提高NADP(H)含量,提高PPP代谢强度[26]。
TCA循环除了能提供碳骨架外,还能合成ATP和NADH[22]。NAD和NADH是吡啶核苷酸的主要类型,起调节辅因子和电子转移的作用,参与钙信号转导[27];NAD和NADH通过多聚ADP核糖基化和蛋白质去乙酰化参与修复DNA[28]。随着质子跨线粒体膜转运,通过TCA循环将NAD转化为NADH[29]。NADH可通过氧化磷酸化进一步氧化为NAD,并伴随ATP的生成[30]。本研究发现,T. roseum侵染早期甜瓜果实的NAD和NADH含量大幅度升高。前人研究发现,大麦感染白粉病后,叶片中NAD含量增加了2 倍[31];NAD可诱导拟南芥PR基因的表达和对丁香假单胞菌(Pseudomonas syringae)的抗性[32]。IDH是TCA循环中的关键酶,催化琥珀酸生成α-酮戊二酸,同时产生NADH[33]。α-酮戊二酸是谷氨酸合成前体[34],谷氨酸在植物抵御病原菌侵染过程中具有重要作用[35]。本研究发现,T. roseum侵染早期甜瓜果实的IDH活力在早期(12 h)显著低于对照组。前人研究发现,过量ATP的产生可抑制TCA循环[11],T. roseum侵染12 h时,能诱导厚皮甜瓜大量ATP合成[16]。因此,认为IDH活力在侵染早期的降低可能受到ATP大量合成的影响。MDH是TCA循环中的另一关键酶[36],催化苹果酸生成草酰乙酸,促进细胞质和亚细胞器之间代谢物的交换和NADH的再生,且与能量代谢密切相关[37]。本研究发现,T. roseum侵染早期大幅度提高了甜瓜果实的MDH活力,该结果与Fusarium oxysporum侵染豌豆后导致的早期MDH活力升高的结果[38]基本一致。由此表明,通过TCA循环形成的能量和碳骨架在厚皮甜瓜果实抵御病原物侵染的早期发挥了重要作用。
PPP不仅能提供碳骨架,同时也是NADPH的主要来源途径[10],同时其中间产物如赤藓糖-4-磷酸是合成可溶性酚酸的前体,酚酸能够激活植物细胞体内信号参与对病原菌的抗性[39-40]。葡萄糖经己糖激酶催化生成的G6P是PPP的初始底物,G6P可为蔗糖、淀粉和细胞壁的合成提供前体物质[41]。NADPH可在NADPH氧化酶的作用下转移电子将O2催化还原为超氧阴离子,参与活性氧产生[42]。NADPH还是谷胱甘肽还原酶的辅酶,以维持还原型谷胱甘肽的正常水平,在清除活性氧的过程中发挥作用,以保护酶、硫氧还蛋白以及细胞膜免受氧化损伤[43]。RPI可催化PPP的非氧化阶段中Ru5P和R5P之间相互转化,在核苷酸、芳香族氨基酸、苯丙烷及其衍生物合成中具有重要作用[44],苯丙烷及其衍生物除具有直接的抑菌作用外,还积极参与了寄主的抗病反应[45]。本研究发现,T. roseum侵染厚皮甜瓜后期显著提高了G6PDH和6PGDH的活力,促进了G6P和NADPH的积累。NADPH含量在侵染的前期和中期显著低于对照组,而在后期显著高于对照组,可能是由于在前期和中期TCA循环旺盛,抑制了PPP[37-38]。后期接种甜瓜果实NADPH含量增加的原因与PPP激活相关[11,24]。由此表明,通过PPP形成的碳骨架和NADPH在厚皮甜瓜果实抵御病原菌侵染的后期发挥了重要作用。
NADK是生物体内唯一能够催化NAD+生成NADP+的酶,该酶以ATP作为磷酰基供体催化NAD(H)反应生成NADP(H)[46]。NADK的激活可提高或维持组织中NADPH水平,以发挥NADPH在抗病中的作用[11]。本研究发现,T. roseum侵染厚皮甜瓜果实后NADK活力先急剧升高后逐步降低,但始终高于对照组。由此表明,NADK催化的NAD+转化为NADP+在果实抗病的不同阶段均发挥了作用,但详细机制有待进一步揭示。
接种果实由于早期能量供应短缺导致病斑出现,为了弥补能量供应短缺,果实的TCA循环被激活,在一定程度上抑制了PPP[11]。接种后期由于防御反应需要大量的碳骨架,因此PPP增强,TCA循环受到了一定程度的抑制[14]。
综上,T. roseum侵染早期能诱导厚皮甜瓜果实大幅度提高TCA循环关键酶MDH的活力,促进NAD和NADH的积累;侵染的后期激活了果实的PPP,增加了G6PDH和6PGDH活力以及G6P、NADH含量。由此表明,TCA循环和PPP在病原菌侵染果实的不同阶段发挥作用,TCA循环更多参与了果实早期的抗病防御,而PPP在后期的寄主防御反应中发挥了更大的作用。
