山西黎城野生萱草群落优势种种间关系与生态位
2021-12-01李志娟李素清
李志娟,李素清
(1. 山西农业大学 园艺学院,山西 太原 030031;2. 山西大学 黄土高原研究所,山西 太原 030006)
种间关系和生态位是植物群落重要的数量和结构特征之一,其中种间关系是指物种间相互吸引或排斥的性质,包括种间关联性与相关性两方面[1]。生态位是指物种在群落中利用资源的能力,包括生态位宽度与重叠两方面[1,2]。对植物群落种间关系和生态位的研究,有助于揭示群落中物种共存、种间相互作用等生态过程[3],正确认识植物群落的结构、功能和演替动态,为植被的经营管理、物种资源保护和持续利用提供理论依据[1]。自20 世纪70 年代末以来,种间关系与生态位研究方法已被广泛应用于我国森林[4,5]、灌丛[6]、草地[7]、湿地[8]、苔藓[9]等群落研究中,但关于萱草种间关系与生态位的研究欠缺。
萱草(Hemerocallisspp.)是阿福花科(Aspho⁃delaceae)萱草亚科(Hemerocallidoideae)萱草属(HemerocallisL.)多年生宿根草本植物的统称[10]。全世界萱草属植物约14 种,主要分布在东亚至俄罗斯西伯利亚地区[11,12]。萱草花期长、耐旱、耐寒、耐贫瘠、生态适应性强[13],是一种重要的园林地被植物和药食同源植物,应用前景广阔[14]。我国是世界萱草属种质的自然分布中心,原产11 种[11]。其中,黄花菜,又名金针菜,原产中国,在河北、山西、山东和秦岭以南各地均有分布,花蕾为著名的“黄花菜”,可供食用[15];北萱草原产中国、俄罗斯和日本,在我国陕西、山西、甘肃、宁夏等地均有分布,生长在海拔500~2 500 m 的山坡、山顶或崖顶,也见于石砾地、杂草丛、路边、林下或林缘[16]。萱草在我国观赏栽培历史悠久,素有“母亲花”之誉[10]。尽管我国萱草野生种质资源丰富,但目前野生萱草资源产业化率低,具有自主知识产权的优良品种极度缺乏[17],以种群为单位对萱草属植物进行的研究欠缺[18]。因此,开展野生萱草群落优势种种间关系与生态位研究,对筛选、培育生态适应性优良的野生萱草,促进萱草产业化应用具有重要参考价值。
山西省黎城县是我国野生萱草的主要分布区之一,目前对该县野生萱草的研究主要集中在染色体数鉴定与核型分析[19]、萱草花芽分化过程中外部形态及营养物质特征等方面[20],关于野生萱草群落优势种种间关系与生态位的相关研究未见报道。基于此,本研究通过总体关联性、M. Gor⁃don 稳定性、种间联结性与相关性、生态位宽度和重叠测定方法,对分布于海拔1 070~1 530 m 的黎城野生萱草群落优势种种间关系与生态位进行分析,揭示其优势种的种间关系与生态位特征,旨在为该县野生萱草种质资源开发提供理论依据。
1 材料与方法
1. 1 研究区域概况
黎城县位于山西省东南部的太行山中南段(113°11′~113°35′E,36°23′~36°53′N),地形总趋势西北多山,东部丘陵,中南部地势较平坦,海拔600~1 500 m。为暖温带半湿润大陆性季风气候,年均温11. 5 ℃,年均降水量600 mm,无霜期约200 d,地带性土壤类型主要为褐土。地带性植被的代表种主要有油松(Pinus tabuliformis)、辽东栎(Quercus liaotungensis)、山杨(Populus davidiana)、野皂角(Cassia mimosoides)、白刺花(Sophora da⁃vidii)、白羊草(Bothriochloa ischaemum)等[21]。
1. 2 研究方法
1. 2. 1 植物样方调查
在山西省黎城县西北部洪井乡南峧一带(113°15′~113°18′E,36°34′~36°38′N)的太行山东麓野生萱草分布区坡度3°~70°、海拔1 070~1 530 m之间[19],根据海拔高度随机设置1 m×1 m 的草本样方。调查每个样方中野生萱草群落植物种的名称、盖度和高度,并记录样方的坡度、坡向等环境因子,同时采用手持GPS 仪测定每个样方的经纬度、海拔高度,共计72 个样方,统计72 个草本植物种。依据各物种在群落中的重要值及频率[1],得到野生萱草群落14 个优势种(表1),形成72×14 的优势种重要值数据矩阵,进行种间关系与生态位分析。

表1 黎城野生萱草群落14 个优势种的特征[22,23]Table 1 Characteristics of 14 dominant species in the wild Hemerocallis spp. community in Licheng County
1. 2. 2 总体关联性测定
将72×14 的优势种重要值数据矩阵转化为0、1 形式的二元数据矩阵,采用D. Schluter 提出的方差比率法(VR)测定群落整体关联性,用W值检验关联显著程度,若VR>1 则物种总体呈正关联,VR<1 则物种总体呈负关联[24]。 采用统计量W(VR×N)检验VR偏离1 是否显著,若W≤χ20.95N或W≥χ20.05N,则种间总体关联显著(P<0. 05)[1]。
1. 2. 3 群落稳定性测定
采用M. Gordon 稳定性测定方法,估测群落的稳定性[25,26]。
1. 2. 4 种间关联性测定
采用72×14 的0、1 形式的二元数据矩阵,进行χ2检验[1]。
1. 2. 5 种间相关性测定
采用72×14 的优势种重要值矩阵,计算Pear⁃son 相关系数和Spearman 秩相关系数[1]。
1. 2. 6 生态种组的判别
采用Spearman 秩相关系数进行聚类分析,划分生态种组[1,27]。
1. 2. 7 生态位宽度和重叠测定
采用Shannon-Wiener 指数、Pianka 指数[13],计算优势种的生态位宽度和重叠值。
1. 2. 8 生态位相似性指数测定
采用Schoener 生态位相似性指数(Cik),计算优势种的生态位相似性指数[6]。
1. 3 数据统计
采用Excel 2010 进行重要值、方差比率、群落稳定性、生态位相似性、宽度和重叠等数据计算与制图;采用SPSS 20. 0 软件进行χ2检验、Pearson 相关和Spearman 秩相关系数检验、聚类分析和回归分析[28,29]。
2 结果与分析
2. 1 总体关联性和群落稳定性
从黎城野生萱草群落14 个优势种总体关联性检验可知,总体关联性方差比率VR=0. 664<1,计算统计量W=47. 790,查表得出χ20.95(72)=52. 600,χ20.05(72)=91. 670,W<χ20.95(72),VR值偏离1显著,说明黎城野生萱草群落14 个优势种总体呈显著负关联(P<0. 01)(表2)。从M. Gordon 稳定性测定可知,该野生萱草群落稳定性曲线类型为y=-0. 009 2x2+1. 703 7x+19. 586,R²=0. 986 6(P<0. 001),其表征群落稳定性的交点坐标比值为34/66,距离坐标20/80 有一定的距离,说明该野生萱草群落结构不稳定(图1)。

表2 黎城野生萱草群落14 个优势种总体关联性检验Table 2 Overall interspecific association of 14 dominant species in the wild Hemerocallis spp. community in Licheng County

图1 黎城野生萱草群落的稳定性测定图Fig.1 Stability diagram of the wild Hemerocallis spp. com⁃munity in Licheng County
2. 2 种间关联性
从14 个优势种的χ2检验半矩阵图(图2)可知,91 个种对中呈正关联(34 对)、负关联的种对(57对)分别占总对数的37. 4% 和62. 6%,正负关联比(0. 60)小于1,其中呈极显著、显著正关联的种对(4 对)占总对数的4. 4%,包括赖草(L. calinus)(Ls)与细叶苔草(C. rigescens)(Cr)/铁杆蒿(A.gmelinii)(Ag)、乳浆大戟(E. Esula)(Ee)与鹅耳枥幼苗(C. turczaninowii)(Ct)、北萱草(H. escu⁃lenta)(He)与狗尾草(S. viridis)(Sv);呈极显著、显著负关联的种对(5 对)占总对数的5. 5%,包括黄花菜(H. citrina)(Hc)与北萱草/狗尾草(Sv)、赖草(Ls)与鹅耳枥幼苗(Ct)、细叶苔草(Cr)与艾蒿(A. argyi)(Aa)、艾蒿(Aa)与野艾蒿(A. lavan⁃dulaefolia)(Al)。总体上看,91 个种对中呈极显著和显著关联的种对数少(9 对),91. 1% 的种对种间关联未达到显著程度,说明黎城野生萱草群落优势种种间关联不紧密。

图2 14 个优势种的χ2检验半矩阵图Fig.2 Semi-matrix of χ2-test for 14 dominant species
2. 3 种间相关性
从Pearson 相关系数检验可知(表3),91 个种对中呈正关联(19 对)、负关联的种对(72 对)分别占总对数的20. 9% 和79. 1%,正负关联比(0. 26)小于1,其中呈极显著、显著正关联的种对(7 对)占总对数的6. 4%,包括北萱草(He)与龙芽草(A.pilosa)(Ap)、赖草(Ls)与乳浆大戟(Ee)/鹅耳枥幼苗(Ct)、茵陈蒿(A. capillaris)(Ac)与连翘幼苗(F. suspense)(Fs)、乳浆大戟(Ee)与鹅耳枥幼苗(Ct)、黄花菜(Hc)与艾蒿(Aa)、北萱草(He)与狗尾草(Sv);呈极显著、显著负相关种对有3 对,占总对数的11. 5%,包括黄花菜(Hc)与北萱草(He)/狗尾草鹅耳枥幼苗(Ct)、北萱草(He)与细叶苔草(Cr)。91 个种对中呈极显著和显著关联的种对少(10 对),89. 0% 的种对间关联程度不显著,表明黎城野生萱草群落优势种种间关系松散,该结果与χ2检验结果一致。
从Spearman 秩相关检验可知(表3),91 个种对中呈正关联的23 对,负关联的68 对,分别占总对数的25. 3% 和占74. 7%,正负关联比(0. 34)小于1,其中呈极显著、显著正关联的有9 对,占总对数的9. 9%,包括赖草(Ls)与乳浆大戟(Ee)/鹅耳枥幼苗(Ct)、乳浆大戟(Ee)与鹅耳枥幼苗(Ct)、北萱草(He)与龙芽草(Ap)/狗尾草(Sv)、细叶苔草(Cr)与铁杆蒿(Ag)、茵陈蒿(Ac)与连翘幼苗(Fs)、皱叶酸模(Rumex crispus)(Rc)与狗尾草(Sv);呈极显著、显著负关联的有7 对,占总对数的7. 7%,包括黄花菜(Hc)与北萱草(He)/铁杆蒿(Ag)/狗尾草(Sv)、赖草(Ls)与细叶苔草(Cr)、细叶苔草(Cr)与艾蒿(Aa)、艾蒿(Aa)与野艾蒿(Al)、野艾蒿(Al)与鹅耳枥幼苗(Ct),表明91 个种对中呈极显著和显著关联的种对少(16 对),82. 4% 的种对间关联程度不显著,优势种种间联结松散,该结果与χ2检验和Pearson 相关系数检验结果一致。

表3 14 个优势种的Pearson 相关和Spearman 秩相关系数Table 3 Pearson’s correlationand Spearman’s rank coefficients of 14 dominant species
2. 4 生态种组
生态种组划分有助于更深入的分析群落的结构和功能[1,6],同一组内的植物种,具有较大的关联性和相关性,在适应环境、资源利用能力等方面具有一致性[1]。采用Spearman 秩相关系进行平均联结(组内部)聚类[6,27,30],并综合物种的生态习性及分布生境,以显著负相关性作为划分种组的界限[7],可将黎城野生萱草群落14 个优势种划分为2个生态种组(图3),导致其生态种组分化的主要原因与物种的生物学特性和生态习性差异有关[6,18]。

图3 14 个优势种的Spearman 秩相关系数聚类图Fig.3 Clustering diagram of Spearman’s rank correlation coeffi⁃cients of 14 dominant species
Ⅰ黄花菜种组:由野生黄花菜(Hc)、细叶苔草(Cr)、艾蒿(Aa)、野艾蒿(Al)、茵陈蒿(Ac)、铁杆蒿(Ag)、皱叶酸模(Rc)、连翘幼苗(Fs)和狗尾草(Sv)组成,该种组优势种由旱生、中旱生到中生植物组成(表1),对水分的要求不高,能在较干旱的条件下生长,建群种野生黄花菜属夜间开花的类群[18],地上生物量大,耐旱、耐贫瘠,生态适应性强,分布范围广,从海拔1 070 m 的山麓地带到1530 m 山地东坡、东北坡均可生长。
Ⅱ北萱草种组:由野生北萱草(He)、龙芽草(Ap)、赖草(Ls)、乳浆大戟(Ee)和鹅耳枥幼苗(Ct)组成(表1),该种组优势种以中生植物为主,对水分的要求较高,建群种野生北萱草属白天开花的类群[18],主要分布海拔1 070~1 530 m 的山地东北坡土层较厚、土壤水分含量较高的林缘下,地上生物量与个体数量比黄花菜小。
2. 5 生态位宽度和重叠
从表4 可知,黎城野生萱草群落生态位宽度值最大的优势种为黄花菜(3. 851),其次为细叶苔草(3. 056)、野艾蒿(2. 983)、北萱草(2. 882)、乳浆大戟(2. 253)、铁杆蒿(2. 179)等;生态位重叠值≥0. 3 的种对(7 对)占总对数的7. 69%,其中黄花菜与细叶苔草(0. 443)/艾蒿(0. 361)、北萱草与龙芽草(0. 415)、乳浆大戟与鹅耳枥幼苗(0. 414)种对的生态位重叠较高,而黄花菜和北萱草的生态位重叠较低(0. 028),说明在黎城野生萱草群落中,黄花菜与细叶苔草/艾蒿、北萱草与龙芽草种对占据的生态空间资源相近,建群种黄花菜和北萱草的生态位宽度较大,生态适应性均较强,但二者占据的生态空间资源不同,对环境有不同的反应[31]。
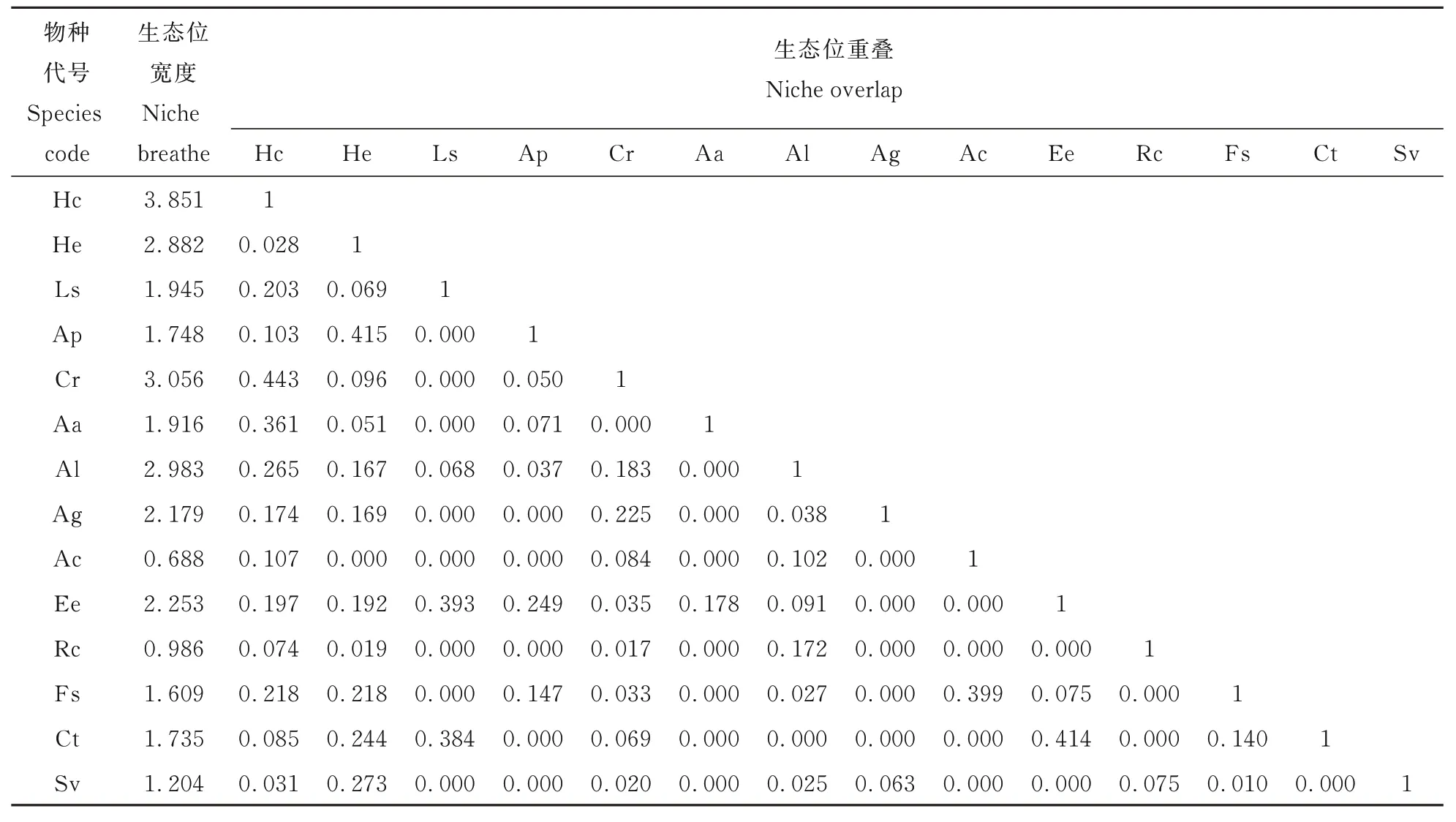
表4 14 个优势种的生态位宽度和重叠Table 4 Niche breathes and overlaps of 14 dominant species
2. 6 生态位相似性
从表5 可知,黎城野生萱草群落生态位相似性(Cik)介于0~0. 354 之间,Cik最大的赖草和乳浆大戟(0. 354),Cik>0. 3 以上的种对(3 对)占3. 30%,介于0. 2<Cik≤0. 3 的种对(6 对)占6. 59%,0. 1<Cik≤0. 2 的种对(14 对)占15. 38%,0. 01<Cik≤0. 1 的种对(30 对)占32. 97%,Cik为0 的种对(38对)占41. 76%。生态位宽度和重叠值最大的优势种黄花菜和细叶苔草的生态位相似性达0. 337,而生态位宽度较窄的皱叶酸模和茵陈蒿的生态位相似性和重叠值为0,说明生态位相似性与生态位宽度和重叠有一定关联。黄花菜和北萱草的生态位相似性较小(0. 047),说明二者对环境资源的需求不同[28],该结果与生态种组划分结果及生态位宽度和重叠结果一致。

表5 14 个优势种的生态位相似性Table 5 Niche similarity of 14 dominant species
3 讨论
3. 1 种间关联性与相关性检验结果的比较
从表6 可知,黎城野生萱草群落14 个优势种的种间关联性检验(χ2检验)与相关性检验(Pear⁃son 相关系数和Spearman 秩相关系数检验)结果一致,且Spearman 秩相关系数检验中呈显著关联的种对数(16 对)比χ2检验(9 对)、Pearson 相关系数检验(10 对)的多,说明Spearman 秩相关系数检验灵敏度高,可更准确反映种间关系,这与简敏菲对九连山常绿阔叶林乔木优势种群的种间关联性分析的结果一致[27]。3 种检验的正负种对间关联比均小于1,且80% 以上优势种对间的关联程度未达到显著水平,说明黎城野生萱草群落整体关联性不紧密,群落结构不稳定(表2,图1),易受外界干扰而发生波动。
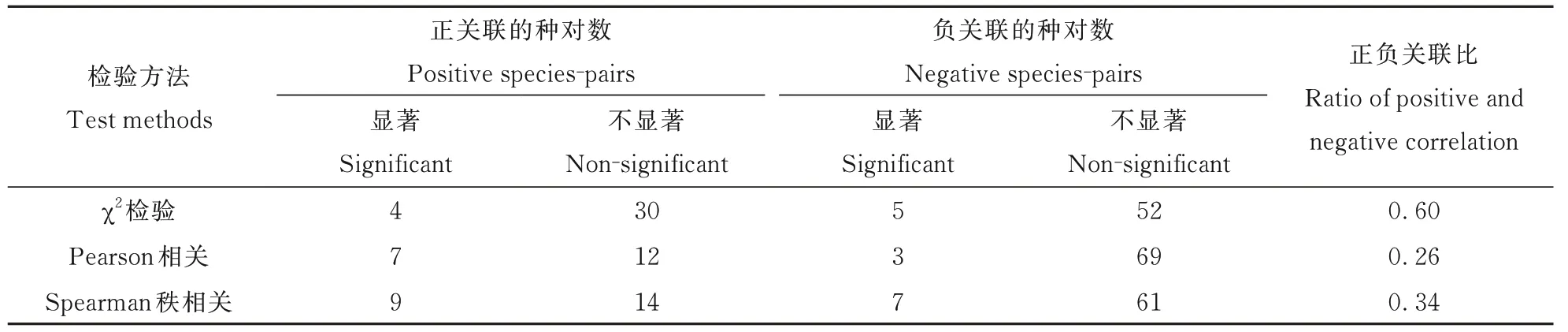
表6 14 个优势种的χ2检验、Pearson 相关检验和Spearman 秩相关检验比较Table 6 The χ2⁃test,Pearson’s correlation coefficients and Spearman’s rank correlation coefficients of 14 dominant species
3. 2 生态种组与种间关系和生态位的相关性
张金屯研究表明,同一生态种组内植物种在环境适应、资源利用等方面相对一致,而组间则表现出明显的差异性[1]。本研究中,Spearman 秩相关系数聚类将黎城野生萱草群落14 个优势种划分为以建群种黄花菜、北萱草为主的2 个生态种组,从种间关系看,同一生态种组内优势种间多呈正相关,而组间呈显著负相关。特别是建群种黄花菜与北萱草种对间χ2检验、Pearson 相关和Spear⁃man 秩相关系数呈极显著负相关(P<0. 01),说明二者的生物学特性不同,彼此对环境有不同的反应,或具有相互分离的生态位。这验证了王孝安等“物种间负相关,是因为它们对环境具有不同的生态适应性和相互分离的生态位所致”的研究结果[31]。
从生态位宽度和重叠值看,建群种黄花菜生态位宽度值大于北萱草,说明黄花菜的生态适应性和资源竞争能力比北萱草强。同时,黄花菜和北萱草的生态位重叠较低,说明二者的生物学属性与生态习性不同,分属于不同的生态种组,这与生态种组划分的结果一致。
3. 3 种间关系与生态位相似性和生态位重叠的回归分析
从种间关系与生态位重叠和生态位相似性值的回归分析看,黎城野生萱草群落14 个优势种的Pearson 相关系数、Spearman 秩相关系数与和生态位相似性生态位重叠值之间均呈极显著正相关(P<0. 001),说明种间正相关越强,其生态位相似性和生态位重叠程度越大;反之,种间负相关越强,其生态位相似性和生态位重叠程度越小(图4)。该结论已在许多有关种间关系与生态位的研究中得到证实[6,32]。本研究中,建群种黄花菜与北萱草种的χ2检验、Pearson 相关检验和Spearman 秩相关检验呈极显著负相关(P<0. 01),二者的生态位重叠较低(0. 028);而赖草与鹅耳枥幼苗呈极显著正相关(P<0. 01),二者的生态位重叠较大(0. 414),与郭志华等对庐山常绿阔叶、落叶阔叶林混交林乔木种群种间联结性研究结论一致[33]。
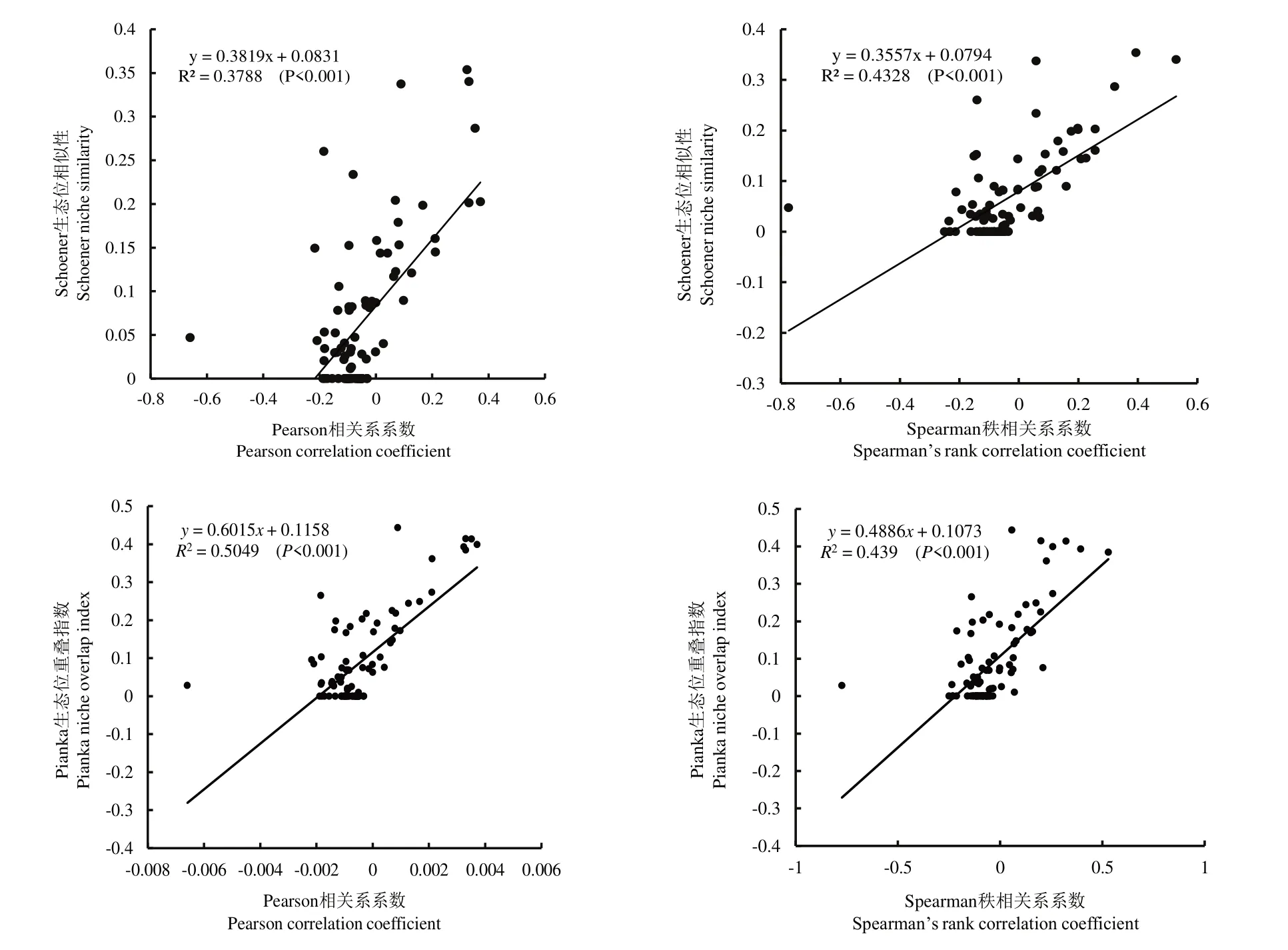
图4 14 个优势种的相关系数与生态位重叠指数的回归分析Fig.4 Regression analysis between correlation coefficient and niche overlap index of 14 dominant species
3. 4 种间关系与生境及物种的生物学属性相关性
陈旭研究表明,“种对间的负相关成因比较复杂,由生境差异导致的负相关,种对间的生态位重叠程度较低,而由资源竞争导致的负相关,种对间的生态位重叠程度较高”[34]。这一结论已在刘润红等关于桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结种间关系与生态位的研究中得到证实[6,32]。据此,综合本研究结果“黄花菜与北萱草种对间3 种相关系数呈极显著负相关(P<0. 01),二者占据不同的生态位,且生态位重叠较低(0. 028)和生态位相似性较小(0. 047)”,可推断出黄花菜与北萱草种对间负相关是由于由生境差异导致的。这一结论与本研究在黎城进行野生萱草群落植被调查的实际相符。
任毅等关于萱草属种质资源多样性的研究表明,萱草属分为昼、夜开花2 种类型,其中夜间开花的黄花菜是进化类群,而白天开花的北萱草进化程度不高[18]。贾贺燕等关于昼夜开花萱草杂交后代花朵开闭时间的研究表明,白天开花萱草主要以蝶类、蜂类为虫媒,而傍晚开花萱草主要以蛾类为虫媒,二者的主要传粉生物及其相关生物学特性不同[35],该结论与本研究中野生萱草群落生态种组划分为2 组及导致其生态种组分化的主要原因与黄花菜和北萱草的生物学特性和生态习性不同有关的结果一致。
4 结论
(1)山西黎城野生萱草群落14 个优势种总体呈显著负关联,二者主要分布于海拔1 070~1 530 m的中低山和山麓地带,受旅游、放牧等人为干扰影响较大,群落结构不稳定,应加强野生萱草种质资源的科学保护。
(2)91 个种对的χ2检验、Pearson 相关和Spear⁃man 秩相关系数检验正负关联比均小于1,多数种间关系不紧密。但建群种黄花菜与北萱草种之间呈极显著负相关,二者生态位宽度较大是生态适应性优良的野生萱草种质资源。
(3)14 个优势种可划分为以夜晚开花的黄花菜和白天开花的北萱草为建群种的2 个生态种组。黄花菜生态位宽度值大于北萱草,但二者的生态位重叠值和生态位相似性较低,表明二者生态习性和生境不同,黎城野生黄花菜的生态适应能力比北萱草强。在未来引种种植中,二者适宜单种。
