三种不同类型亚高山森林凋落物输入对土壤腐殖化的影响
2021-11-18卫芯宇倪祥银陈子豪杨玉莲吴福忠
卫芯宇,倪祥银,谌 亚,陈子豪,杨玉莲,吴福忠,*
1 福建师范大学地理科学学院, 福州 350007
2 四川农业大学生态林业研究所, 林业生态工程重点实验室, 成都 611130
腐殖质作为土壤有机质的主要组成部分[1-2],不仅是土壤养分的重要来源,也有助于维持土壤结构和功能[3]。凋落物作为养分和碳的载体,是连接植物和土壤的“纽带”[4]。然而,凋落物输入土壤后,难降解物质的增加能为土壤腐殖质的合成提供原料,促进土壤腐殖质的合成[5-6],也可能通过增加易降解组分激发土壤微生物和酶的活性,引起“激发效应”,导致原有土壤腐殖质的降解[7-8],这说明凋落物与土壤腐殖质的关系尚不明确。腐殖质的颜色能很好的反映其腐殖化程度[9],腐殖质的降解会导致其颜色变浅[10-11],而随着颜色加深,腐殖质的结构会变得更加复杂且稳定[12]。因此,Kumada等人首先提出使用色调系数(ΔlogK)来表征腐殖质的复杂程度[13],而Kononova和Alexandrova则使用光密度值(E4/E6)作为腐殖化的指标[14]。ΔlogK、E4/E6和A600/C值都与腐殖物质芳烃共轭结构复杂性、分子量和缩合度相关[15- 16]。随着ΔlogK和E4/E6值的增加,芳构化程度降低,分子量和缩合度降低,这表明腐殖化程度较低,腐殖物质对土壤有机质的贡献越小[17- 18],而A600/C值则表现出相反的规律。
川西亚高山森林多分布在青藏高原东缘,海拔2000—4000 m,在调节区域气候、保持水土、保育生物多样性等方面具有重要的作用[19]。亚高山地区冬季漫长且伴随明显的季节性冻融循环[20],这些特殊的气候环境不仅不利于土壤腐殖质的累积[21],甚至加速已形成腐殖质的降解[9,22],且低温环境也会对凋落物的分解产生影响[23- 24],从而影响该地区土壤的腐殖化过程。同时,不同森林类型的凋落物具有不同的基质质量,会对土壤腐殖质产生不同的影响[25- 26]。更为重要的是,气候变暖所导致的冬季缩短以及更加频繁的冻融循环会对土壤的腐殖化产生直接或间接的影响[27- 28]。基于以上研究,本文假设凋落物的输入可能在一定程度上提高土壤的腐殖化度,但受到季节和林型的影响。本研究在前期工作的基础上,选择川西亚高山地区具有代表性的针叶林、针阔混交林以及阔叶林为研究对象,通过控制凋落物的输入,研究该地区凋落物和气候变化对土壤腐殖化程度的影响,以期为该地区森林生态系统的发展提供依据。
1 试验地概况
试验地位于四川省平武县王朗国家级自然保护区(32°49′—33°02′ N,103°55′—104°10′ E),海拔2300—4980 m。该地区年平均温度2.5—2.9℃,最高温26℃(7月),最低温-18℃(1月)。冬季时间长,通常从第1年10月延伸到第2年4月[29]。年平均降水量约为826 mm,主要集中在5月到8月。根据中国土壤系统分类,该地区土壤类型为暗棕壤。本研究在该地区海拔2540—2600 m处选择具有代表性的针叶林、针阔混交林以及阔叶林为研究对象。样地详细情况见表1。

表1 针叶林、针阔混交林和阔叶林的海拔,坡向,坡度,优势树种和代表性灌木Table 1 Altitude, aspect, slope, dominant arboreal species and representative shrubs of the coniferous forest, mixed forest and broadleaved forest in the study site
2 研究方法
2.1 试验设计
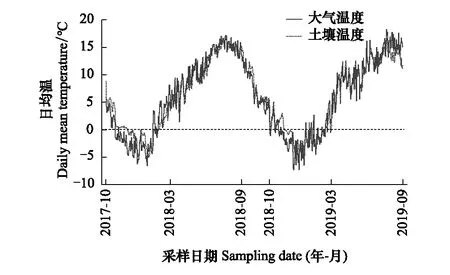
图1 2017年10月至2019年9月青藏高原东部亚高山森林的大气和土壤日平均温度Fig.1 Daily mean temperature of air and soil in the subalpine forest on the eastern Tibetan Plateau from October, 2017 to September, 2019
2017年5月,在针叶林、针阔混交林以及阔叶林内设置原位培养试验,每个森林内选取1个大小为100 m×100 m的样地,每个样地内随机设置3个样方,每个样方包含2种处理(保留凋落物和去除凋落物)。为了确保样方的同质性,所有样方均设置在相似的坡向和坡度(表1)。在每个样方中设置6个长70 cm、宽51 cm、高43 cm的培养框,共计54个(每个样方2种处理×3个重复×3个样方×3种林型)。挖掘深度约30 cm的土壤,在保证其理化性质不改变的情况下,去除其中的凋落物、石头、根系等杂质,混合均匀后放入相应位置的培养框中。在培养框底部钻小孔,避免培养框内积水,同时在底部铺设尼龙网,确保水流出的同时不会带走土壤。将每个样方中的3个培养框设置为凋落物输入框,允许凋落物的自然输入;在3个培养框上方50 cm处设置尼龙网,拦截凋落物的输入,使凋落物无法进入培养框内,将其设置为凋落物去除框。在每个样方内随机选择1个培养框并在深度约15 cm处放置纽扣温度计(iButton DS1923-F5, Maxim/Dallas Semiconductor, Sunnyvale, USA),用以记录土壤温度。每个样方内随机悬挂1个温度记录仪(LITE5032P-RH, Fourtec-Fourier Technologies, Israel),悬挂高度距离地面约2 m,以记录样地内大气温度(图1)。
2.2 样品采集与处理
从2017年10月到2019年9月,分别在每年的10月、3月和9月采集土壤和凋落物样品,共6次。根据长期监测的温度数据[20],将两次采样之间的时期命名为冬季(2017年10月—2018年3月,2018年10月—2019年3月)、生长季前期(2018年3月—2018年9月,2019年3月—2019年9月)、生长季后期(2018年9月—2018年10月)。
每次采样时,首先收集凋落物去除框上方尼龙网内拦截的凋落物,并将来自同一样方的凋落物混合。然后多点随机收集每个培养框内0—20 cm(区域森林土壤有机层的平均深度)的土壤样品并混合均匀。将收集的所有凋落物和土壤样品带回实验室,去除杂质后在室温下风干2周。将凋落物样品分类为针叶,阔叶,枝,花、果实和其他(包括无法识别的植物残留物和动物粪便)(图2)。将土壤样品研磨并过0.25 mm筛用于测定土壤腐殖物质光密度值。

图2 各关键时期3种森林不同类型凋落物产量Fig.2 Average production of different litter components in the three forests in the different sampling periods on the eastern Tibetan Plateau
2.3 指标测定与计算方法
ΔlogK、E4/E6和A600/C的测定参考Chen等[30]的方法。称取风干样品0.500 g于150 mL锥形瓶,加入100 mL 0.1 mol/L NaOH和Na4P2O7·10 H2O混合提取液,加塞振荡10 min,沸水浴1 h,待冷却后过滤,再过0.45 μm滤膜,滤液即为浸提液[31]。取浸提液10 mL,加0.05 mol/L NaHCO3调节pH至7—8,用紫外可见光谱分析仪(TU- 1901, Puxi, Beijing, China)测定400 nm、465 nm、600 nm和665 nm处的吸光值。
腐殖质的光密度与可见波长存在密切的关系。因此,色调系数(ΔlogK)、光密度值(E4/E6)以及A600/C值等光学指标近年来被广泛用于表征腐殖质分子结构的复杂程度与腐殖化程度[15]。ΔlogK值越大,E4/E6比值越高,A600/C值越低,表明腐殖质的光密度愈小,芳香核原子团越少,芳香缩合度低,说明其分子结构越简单,腐殖化程度越低。
ΔlogK=log (A400/A600)
E4/E6=A465/A665
式中,A400、A600、A465、A665分别为400、600、465、665 nm处吸光值。A600/C中的C表示每1 mL提取液中含有的有机碳。
2.4 数据处理
采用重复测量方差分析(Multi-way ANOVA)检验采样时间、森林类型和凋落物处理对土壤腐殖质ΔlogK、E4/E6和A600/C值的影响。用最小显著差异法(Least significant difference, LSD)检验同一时期不同林型土壤腐殖化程度的差异以及不同时期同一林型土壤腐殖化程度的差异。显著性水平设为P=0.05。以上数据分析在SPSS 20.0(IBM SPSS Statistics Inc., Chicago, IL, USA)中进行,在Origin Pro9.0(OriginLab, Northampton, MA)中绘图。
本研究同步检测了培养过程中的土壤基质质量和气候因素共10个变量,为了解这些变量对土壤腐殖化程度的影响,采用偏最小二乘法(partial least squares, PLS)计算各变量的PLS系数(对因变量的作用方向和强度)和重要值(显著性水平;n=108)。以上分析在SIMCA 14.0(Umetrics, Umeå, Sweden)中进行。
3 结果分析
3.1 凋落物输入对土壤腐殖物质ΔlogK值的影响
培养试验表明,亚高山3种森林凋落物去除对土壤腐殖质ΔlogK值并无显著影响(P>0.05),但采样时期和森林类型之间存在显著差异(P<0.01,表2)。阔叶林土壤ΔlogK值显著大于混交林和针叶林土壤,混交林土壤ΔlogK值略高于针叶林土壤,但仅在试验初期有显著差异(图3)。尽管3种林型土壤ΔlogK值在不同凋落物处理下均表现出先逐渐增加而后下降的动态特征,但凋落物去除显著改变了不同林型土壤ΔlogK值的变化趋势。保留凋落物的阔叶林土壤ΔlogK值保持持续增加的动态特征直至第2个冬季结束,且冬季增加趋势更为明显,而去除凋落物的阔叶林土壤ΔlogK值在第1个冬季表现出明显的增加趋势,在第2个冬季表现出明显下降的特征。相对于阔叶林,混交林土壤ΔlogK值在冬季对凋落物去除表现出明显的响应差异,针叶林土壤ΔlogK值在整个研究期间对凋落物去除的响应均不明显(图3)。

图3 培养期间针叶林、混交林和阔叶林中土壤腐殖质的ΔlogK值Fig.3 ΔlogK values in the coniferous forest, mixed forest, and broadleaved forest数值为平均值±标准误, n=3;小写字母表示相同处理下同一时期不同林型之间的差异显著性,大写字母表示相同处理下同一林型不同时期之间的差异显著性

表2 不同采样时期、凋落物处理、森林类型对土壤腐殖质ΔlogK、E4/E6和A600/C值的重复测量方差分析Table 2 Repeated measures ANOVA results for the effects of time, litter, type and their interactions on the value of ΔlogK, E4/E6 and A600/C
3.2 凋落物输入对土壤腐殖物质E4/E6值的影响
与ΔlogK值相似,试验结果表明,亚高山3种林型下凋落物去除对土壤E4/E6值并无显著影响(P>0.05),但采样时期和森林类型显著影响E4/E6值(P<0.01,表2)。阔叶林土壤E4/E6值显著大于混交林和针叶林土壤,而混交林和针叶林土壤E4/E6值无显著差异(图4)。尽管3种林型土壤E4/E6值在不同凋落物处理下均表现出先逐渐增加而后下降的动态特征,但去除凋落物改变了不同林型土壤E4/E6值的变化趋势(图4)。阔叶林土壤E4/E6值保持持续增加的动态变化直至第2个冬季结束,且冬季的增加趋势更为明显,但阔叶林土壤E4/E6值对凋落物去除的响应不明显。针叶林土壤E4/E6值在第1年变化较小,第2年冬季明显增加,生长季下降,且对凋落物去除的响应不明显。混交林土壤E4/E6值对凋落物去除的响应主要表现在冬季,冬季去除凋落物后混交林土壤E4/E6值下降趋势大于保留凋落物土壤。
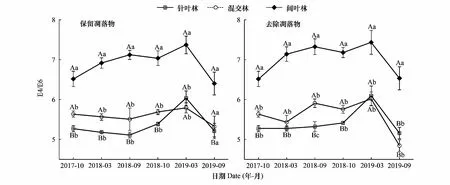
图4 培养期间针叶林、混交林和阔叶林中土壤腐殖质的E4/E6值Fig.4 E4/E6 values in the coniferous forest, mixed forest, and broadleaved forest数值为平均值±标准误, n=3;小写字母表示相同处理下同一时期不同林型之间的差异显著性,大写字母表示相同处理下同一林型不同时期之间的差异显著性
3.3 凋落物输入对土壤腐殖物质A600/C值的影响
培养试验结果表明,亚高山3种林型下凋落物去除对土壤A600/C值并无显著影响(P>0.05),但采样时期和森林类型显著影响A600/C值(P<0.01,表2)。与ΔlogK和E4/E6值不同,阔叶林土壤A600/C值始终显著低于针叶林和混交林,而混交林土壤A600/C值仅在第1年生长季显著低于针叶林(图5)。3种森林土壤A600/C值均表现出逐渐增加的动态趋势,且针叶林和混交林土壤A600/C值在生长季前期的增加趋势较阔叶林更加显著。针叶林在整个研究期间对凋落物去除的响应不明显。保留凋落物的混交林土壤A600/C值在第1年冬季表现为下降,而去除凋落物土壤A600/C值上升。保留凋落物的阔叶林土壤A600/C值在第2年冬季下降,而去除凋落物土壤A600/C值上升(图5)。

图5 培养期间针叶林、混交林和阔叶林中土壤腐殖质的A600/C值Fig.5 A600/C values in the coniferous forest, mixed forest, and broadleaved forest数值为平均值±标准误, n=3;小写字母表示相同处理下同一时期不同林型之间的差异显著性,大写字母表示相同处理下同一林型不同时期之间的差异显著性
4 讨论
腐殖物质的光密度值在一定程度上反映了其分子的复杂程度,即ΔlogK、E4/E6值越低,说明腐殖质中芳香核原子团越多,分子结构越复杂[32- 33],从而间接表明土壤的腐殖化程度越高[15]。本研究结果表明,亚高山地区3种森林土壤的ΔlogK和E4/E6值大小依次为:阔叶林>混交林>针叶林,A600/C值大小依次为针叶林>混交林>阔叶林。3种森林土壤的腐殖化程度随季节发生变化,冬季土壤腐殖质的ΔlogK和E4/E6值升高,而生长季节降低,A600/C值则在冬季下降,生长季节升高。试验表明,保留凋落物的阔叶林和混交林土壤腐殖质ΔlogK值在冬季呈下降趋势,而去除凋落物的阔叶林和混交林土壤ΔlogK值在冬季呈上升趋势。这些结果表明,亚高山针叶林土壤的腐殖化程度最高,而阔叶林土壤最低。土壤腐殖化程度主要受到温度和森林类型的调控,温度升高会导致土壤腐殖化程度增加,而冬季凋落物输入会导致土壤腐殖化程度降低。
地上植被变化所引起的凋落物、根系及其分泌物的改变会显著影响土壤的物理化学性质[34];而土壤作为生态系统中诸多生态过程的载体,不同土壤养分状况又会作用于群落内的许多生态过程。本研究发现,川西亚高山地区3种林分土壤的腐殖化程度依次为:针叶林>混交林>阔叶林,这与凋落物的性质和数量有关。与阔叶凋落物相比,针叶凋落物中含有大量难分解的纤维素和木质素,因此针叶凋落物的分解较慢[35],容易累积大量的难分解物质在土壤中聚合形成腐殖质,使得土壤腐殖物质的分子结构更复杂,腐殖化程度更高。同时,由于针叶凋落物分解过程中会产生酸性物质[36-37],不利于土壤微生物活性,所以针叶林土壤腐殖质的降解也是相对较慢的。而混交林土壤腐殖化程度高于阔叶林,可能是因为混交林的凋落物种类更加的丰富,且凋落物量也更大(图2),较阔叶林更有利于土壤腐殖质的合成,所以混交林土壤腐殖化程度高于阔叶林。
亚高山森林由于全年温度较低,所以凋落物的分解也缓慢[38]。培养前期由于凋落物对微生物的养分供应不足,微生物更多的从土壤中汲取养分,导致土壤腐殖质发生降解,所以3种森林土壤的腐殖化程度在培养前期一直降低(图3—5)。3种森林土壤的腐殖化程度在生长季节开始升高,这可能与温度升高存在相关性,且偏最小二乘法分析的结果也印证了这一结果(图6)。土壤和大气正积温与ΔlogK、E4/E6值呈负相关关系,与A600/C值呈正相关关系,这说明热量资源丰富会提高土壤的腐殖化程度。一方面,低温限制微生物活性,导致微生物途径的土壤腐殖质合成受阻[28],且冬季积雪所导致的低氧甚至厌氧环境不仅不利于土壤腐殖质的稳定性[11,39- 40],甚至破坏土壤腐殖质的结构[9],导致土壤腐殖化程度降低。而生长季节,随着温度的升高,微生物活性增强[41- 42],土壤中的腐殖质被不断加工,所以土壤腐殖化程度增加。另一方面,生长季节凋落物量相对更多,且种类丰富,为土壤腐殖质的合成提供原料。且生长季凋落物多以叶凋落物为主(图2),相较枝凋落物而言含有更多的有效成分和营养物质,能刺激微生物活动,并进一步促进土壤的腐殖化进程[43]。

图6 环境因子和土壤基质质量对土壤腐殖化度的偏最小二乘法结果Fig.6 Results of partial least squares (PLS) testing for the effect of environmental factors and soil qualities on the humification of soil humic substances during incubation periods偏最小二乘法系数大于0表示正作用,小于0表示负作用;变量重要值大于1表示作用显著(P<0.05);C, 碳 carbon; N, 氮 nitrogen; P, 磷 phosphorus; NFTC, 冻融循环次数 number of freeze-thaw cycles; MAT, 大气均温 mean air temperature; MST, 土壤均温 mean soil temperature; APAT, 大气正积温 air positive accumulation temperature; ANAT, 大气负积温 air negative accumulation temperature; SPAT, 土壤正积温 soil positive accumulation temperature; SNAT, 土壤负积温 soil negative accumulation temperature. 数值为各采样时期数据(n=108)
整体而言,凋落物输入在生长季节对3种森林土壤的腐殖化程度没有显著影响,但在冬季改变了混交林和阔叶林土壤的腐殖化程度。导致这一结果的原因可能是亚高山地区全年温度较低,凋落物分解缓慢。凋落物覆盖在土壤表面具有一定的保温作用[44],所以在低温环境下保留凋落物土壤中的微生物活性更高[45],这使得由微生物驱动的土壤腐殖物质的合成过程受气候条件的影响更小。同时凋落物输入会积累更多的难降解物质到土壤表面,为土壤腐殖质的合成提供原料[5],但新形成的土壤腐殖质结构相对简单,所以其腐殖化程度更低[46]。去除凋落物处理下,由于冬季的低温和频繁的冻融循环,土壤中不稳定、结构简单的腐殖质被破坏和矿化[47],而结构复杂且稳定的腐殖质被保留下来,所以去除凋落物土壤的腐殖化程度更高。去除凋落物对针叶林土壤腐殖化程度的影响不显著,这可能与针叶林土壤本身较高的腐殖化程度,以及针叶凋落物分解缓慢且易产生酸性物质有关。
5 结论
亚高山森林土壤的腐殖化程度与温度存在显著的正相关关系,表现为冬季土壤腐殖化程度降低,而生长季节升高的动态规律。针叶林土壤的腐殖化程度最高,混交林次之,阔叶林最低。冬季凋落物输入显著降低了阔叶林和混交林土壤的腐殖化程度,而在生长季节无显著影响,且凋落物的输入对针叶林土壤腐殖化程度并无显著影响。总之,气候变暖情景下冬季温度的升高可能导致亚高山森林土壤的腐殖化程度升高,且受到森林类型的调控。这些结果为了解亚高山森林土壤腐殖质的形成与动态及其对气候变化的响应提供了一定的科学依据。
