黄淮稻区早熟水稻品种(品系)穗颈瘟抗性分析
2021-11-14邢运高刘艳迟铭陈庭木孙志广徐波杨波李景芳李健方兆伟卢百关王宝祥徐大勇
邢运高 刘艳 迟铭 陈庭木 孙志广 徐波 杨波 李景芳 李健 方兆伟 卢百关 王宝祥 徐大勇


摘要: 為明确黄淮稻区早熟水稻品种(品系)的稻瘟病抗性,对145份黄淮稻区早熟水稻品种(品系)进行连续2年的接种鉴定,并利用 Pi-ta 、 Pi-b 、 Pi-km 、 Pi-54 、 Pi-5 和 Pi-gm 6个抗病基因的分子标记进行抗稻瘟基因型检测。抗性基因在检测品种(品系)中的分布结果表明,抗性基因 Pi-b 分布比例最高,检出率为57.9%,其次是抗性基因 Pi-km、Pi-ta、Pi-54 和 Pi-5 ,检出率分别为51.0%、42.1%、32.4%及32.4%,抗性基因 Pi-gm 分布比例最低,仅有4.1%。2019-2020年连续2年的人工接种鉴定结果显示,74.5%和68.3%的品种(品系)表现为感病,说明黄淮稻区早熟品种(品系)的稻瘟病抗性较差,抗病材料较少。携带抗性基因 Pi-ta 、 Pi-b 、 Pi-km 、 Pi-54 、 Pi-5 和 Pi-gm 的品种(品系),2019年抗性比例分别为42.6%、21.4%、27.7%、34.3%、27.7%和100.0%,2020年抗性比例分别为49.2%、26.2%、25.5%、32.9%、34.0%和100.0%。6个基因中, Pi-gm 抗性比例最高,达到100.0%, Pi-ta 次之,抗性比例达45.9%,而 Pi-b 最差,抗性比例仅为23.8%。黄淮稻区早熟水稻品种(品系)抗稻瘟病能力较差,需要通过聚合多个抗性基因来提高抗性,特别需要加强 Pi gm 等新的抗稻瘟病基因的应用。
关键词: 水稻; 早熟品种(品系); 稻瘟病; 抗性基因
中图分类号: S435.111.4 +1 文献标识码: A 文章编号: 1000-4440(2021)05-1089-11
Resistance analysis of early-maturing rice varieties (strains) to neck blast in Huang-Huai rice region
XING Yun-gao, LIU Yan, CHI Ming, CHEN Ting-mu, SUN Zhi-guang, XU Bo, YANG Bo, LI Jing-fang, LI Jian, FANG Zhao-wei, LU Bai-guan, WANG Bao-xiang, XU Da-yong
(Lianyungang Academy of Agricultural Sciences/Jiangsu Collaborative Innovation Center for Modern Crop Production, Lianyungang 222006, China)
Abstract: To clarify the resistance of early-maturing rice varieties (strains) to rice blast in Huang-Huai rice region, 145 early-maturing rice varieties (strains) in Huang-Huai rice region were identified through artificial inoculation in two consecutive years. The rice blast resistant genotypes were analyzed by molecular markers of six resistance genes such as Pi-ta 、Pi-b、Pi-km、Pi-54、Pi-5 and Pi-gm . Among the tested varieties (strains), resistance gene Pi-b had the highest distribution proportion, with a detection rate of 57.9%, followed by resistance genes Pi-km , Pi-ta , Pi-54 and Pi-5 , their detection rates were 51.0%, 42.1%, 32.4% and 32.4% respectively, and resistance gene Pi-gm had the lowest distribution proportion, which was only 4.1%. The identification results of artificial inoculation in two consecutive years from 2019 to 2020 showed that, 74.5% and 68.3% of the varieties (strains) were sensitive to rice blast in 2019 and 2020, respectively. It was indicated that the resistance of early-maturing varieties (strains) in Huang-Huai rice region was poor and the resistant materials were few. The resistance proportions of the varieties (strains) carrying resistance genes Pi-ta、Pi-b、Pi-km、Pi-54、Pi-5 and Pi-gm were 42.6%, 21.4%, 27.7%, 34.3%, 27.7% and 100.0% respectively in 2019, and were 49.2%, 26.2%, 25.5%, 32.9%, 34.0% and 100.0% in 2020. Among the six genes, Pi-gm had the highest resistance proportion, which reached 100.0%, followed by Pi-ta , with resistance proportion of 45.9%. Pi-b had the lowest resistance proportion, with a resistance proportion of only 23.8%. The resistance of the early-maturing rice varieties (strains) to rice blast in Huang-Huai rice region is poor, and is necessary to aggregate multiple resistance genes to improve the resistance of rice varieties (strains), especially to strengthen the application of new blast resistance genes.
Key words: rice; early-maturing varieties (strains); rice blast; resistant gene
黄淮稻区水稻面积约 1.60× 10 6 hm 2 ,占南方粳稻面积的一半左右,是中国重要的粳稻产区 [1] 。稻瘟病一直是危害水稻生產的重要病害,严重威胁中国的水稻生产安全。黄淮稻区地处中国南北过渡带,年际间气候不稳定,特别在粳稻抽穗灌浆期间经常遭遇连续的低温、阴雨等不良天气。适温高湿的环境利于稻瘟病,特别是穗颈瘟的暴发,严重影响水稻的生产安全。此外,稻瘟病病菌小种遗传复杂,种群演替迅速,克服寄主抗病性的小种能快速形成优势小种,因此新品种推广数年后面临抗性丧失的风险 [2] 。研究结果表明,稻瘟病抗性基因的导入和聚合是解决该问题的最有效措施之一 [3-6] 。
随着农村优质劳力的进一步转移、劳动力成本的提升以及规模化经营的发展,能省去育秧、移栽等环节的直播稻越来越多地受到农户的青睐,成为现代稻作的发展方向之一 [7] 。江苏省的直播稻已由苏南和沿江地区发展到苏中、苏北地区,而且面积还在逐年扩大 [8] 。多年试验及大田生产情况表明,早熟水稻品种对穗颈瘟抗性较差,因此对早熟水稻品种进行抗稻瘟基因型分子检测并综合评价抗稻瘟病基因效应,对培育直播稻新品种具有重要的意义。
当前,一些抗性基因如 Pi-ta、Pi-b、Pi-km、Pi-54、Pi-5、Pi-gm、Pi9、Pi2 等已经在生产上广泛运用并表现出良好的抗瘟性。 Pi-ta 与 Pi-b 在江苏省粳稻中分布频率较高,对穗颈瘟表现出较好的抗性 [9] ,是江苏省粳稻育种主要利用的2个基因。 Pi-kh 是一个具有较高利用价值的基因,对不同地区的生理小种具有广谱抗性 [10] 。 Pi-km 为在江苏粳稻中分布较广的抗性基因,位于11号染色体上,由相邻的2个NBS-LRR类基因组成 [11-12] 。陈峰等 [13] 认为 Pi-ta、Pi-b、Pi-kh、Pi-km 在黄淮粳稻中广泛应用。张善磊等 [14] 认为 Pi-ta 对穗颈瘟的抗性效果最好,其次是 Pi-5 、 Pi-km 、 Pi-b 。宋兆强等 [3] 的研究结果表明, Pi-ta+Pi-b 基因组合的抗性能力有减弱趋势。 Pi-gm 是从谷梅4号中鉴定到的一个广谱抗稻瘟病基因。王小秋等 [15] 认为携带 Pi-gm 基因的品系对穗颈瘟的抗性较好。以上研究结果表明, Pi-ta+Pi-b 基因组合模式虽然在江苏省粳稻抗稻瘟病方面发挥重要的作用,但是其抗性效应正在减弱。前人对水稻资源材料进行了抗稻瘟病基因检测 [13] ,但对黄淮稻区早熟品种的抗性分析还未有报道。本研究利用黄淮稻区145份早熟水稻种质资源,连续2年进行稻瘟病抗病性鉴定,并利用抗病基因 Pi-5 、 Pi-ta 、 Pi-b 、 Pi-km 、 Pi-54 和 Pi-gm 分子标记对其进行基因型检测,综合评价黄淮稻区早熟品种的稻瘟病抗性。
1 材料与方法
1.1 试验材料
为了研究适合黄淮稻区直播的水稻品种的稻瘟病抗病性,选用的水稻材料为145份黄淮稻区早熟品种(品系),这些材料在连云港地区于5月20日播种,8月20日之前抽穗,抽穗时间比江苏省中熟中粳早熟组区域试验对照品种苏秀867早3 d。所有品种(品系)同一时间播插,每个品种种植4行,每行10株。田间管理同一般大田管理。
1.2 稻瘟病抗性鉴定方法
2年稻瘟病抗性鉴定菌株均引自江苏省农业科学院植物保护研究所。2019年接种鉴定的菌株是ZB3、ZC15、ZD5、ZE3、ZF1、ZG1,共6个。2020年接种鉴定的菌株是ZB7、ZC11、ZD7、ZF1、ZG1,共5个。
抗性鉴定参照刘永锋等 [16] 的方法,在水稻孕穗初期采用混合菌株进行接种,用注射器吸取1 ml混合菌液在幼穗中上部缓缓注入稻苞内,每个品种(品系)接种5株,每株接种1个穗子。按照接种结果,将稻瘟病抗性等级分为5 级: 0级(免疫),1级(抗病),2级(中抗),3级(感病),4级(高感)。
1.3 稻瘟病抗性基因检测分子标记与基因型检测
参照前人的方法 [17-22] ,利用与 Pi-ta 、 Pi-b 、 Pi-km 、 Pi-54 、 Pi-5 和 Pi-gm 基因紧密连锁的分子标记检测早熟品种(表1)。在分蘖盛期采集水稻叶片,利用十六烷基三甲基溴化铵(CTAB)法提取水稻DNA,参照曾生元等 [17] 的方法对早熟品种(品系)进行基因型鉴定。
1.4 数据处理
利用Excel 2016 对数据进行处理和作图。采用SPSS 16.0对品种(品系)携带的抗病基因数和穗颈瘟抗性级别进行相关分析。
2 结果与分析
2.1 黄淮稻区早熟品种(品系)稻瘟病抗性水平
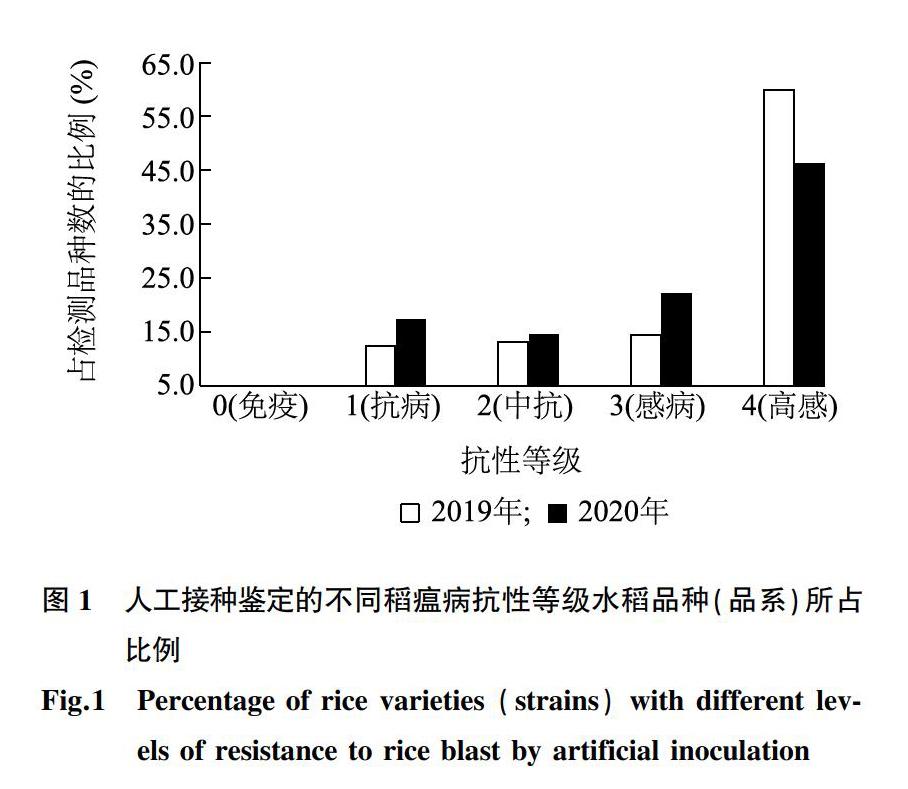
为了明确早熟水稻品种(品系)的稻瘟病抗性情况,对145个黄淮稻区早熟品种(品系)资源进行了连续2年稻瘟病接种鉴定。 145个早熟水稻品种(品系)中,2019年接种鉴定的免疫、抗病、中抗、感病、高感品种(品系)分别有0份、18份、19份、21份、87份,所占比例分别为0%、12.4%、13.1%、14.5%、60.0%,抗性水平在高感水平分布的比例较高;2020年免疫、抗病、中抗、感病、高感品种(品系)分别有0份、25份、21份、32份、67份,所占比例分别为0%、17.2%、14.5%、22.1%、46.2%,在高感、感水平分布的比例较高(图1)。2年都未发现0 级(免疫)的材料,连续2年的接种结果表明,有70%左右的品种(品系)表现为感病,说明黄淮稻区早熟品种(品系)的稻瘟病抗性较差,抗病材料较少。
2.2 黄淮稻区早熟品种(品系)携带的抗稻瘟病基因类型
利用 Pi-ta 、 Pi-b 、 Pi-km 、 Pi-54 、 Pi-5 和 Pi-gm 特异分子标记检测了145份黄淮稻区早熟品种(品系)(表2),检测出的抗稻瘟病基因数量为 0~ 5个,检出率为93.8%。有 61个品种(品系)含有 Pi-ta ,有84个品种(品系)含有 Pi-b ,有47个品种(品系)含有 Pi-54 ,有74个品种(品系)含有 Pi-km ,有47个品种(品系)含有 Pi-5 ,只有6个品种(品系)检测出 Pi-gm ,6个抗性基因都没检测到的材料有9 份。抗性基因 Pi-b 检出率为57.9%,分布比例最高,表明该基因在黄淮稻区粳稻品种得到较为广泛的应用;其次是 Pi-km、Pi-ta、Pi-54 和 Pi-5 ,检出率分别为51.0%、42.1%、32.4%及32.4%;抗性基因 Pi-gm 分布比例最低,仅有4.1%(图2)。
2.3 含有抗稻瘟病基因的抗性效应分析
根据稻瘟病抗性基因分布和连续2年接种结果,进一步分析 Pi-ta 、 Pi-b 、 Pi-km 、 Pi-54 、 Pi-5 和 Pi-gm 抗性基因對水稻品种(品系)抗性比例的贡献。在 145 份测试品种(品系)中(表2), 2019年含有 Pi-ta 、 Pi-b 、 Pi-km 、 Pi-54 、 Pi-5 和 Pi-gm 的水稻品种(品系),接种鉴定表现为抗病品种(品系)的比例分别为42.6%、21.4%、27.7%、34.3%、27.7%和100.0%;2020年含有 Pi-ta 、 Pi-b 、 Pi-km 、 Pi-54 、 Pi-5 和 Pi-gm 的水稻品种(品系),接种鉴定表现为抗病品种(品系)的比例分别为49.2%、26.2%、25.5%、32.9%、34.0%和100.0%。2年结果表明,6个基因中, Pi-gm 抗性比例最高,达到100.0%, Pi-ta 次之,抗性比例达45.9%,而 Pi-b 最差,抗性比例仅为23.8%。上述结果表明, Pi-gm 对黄淮稻区早熟品种(品系)资源稻瘟病的抗瘟性效应最大, Pi-ta 次之,而 Pi-b 最差。对检测出 Pi-gm 基因材料的来源进行分析可知,创新1号、落育6、回粳12、东北086、GM-1和GM-2等6个来源于东北三省的资源中检测出 Pi-gm 基因,而在其他品种(品系)中未检测出该基因。携带 Pi-ta 基因品种(品系)的抗病比例达到40.0%以上,并且品种(品系)中检出率也达到40.0%以上,说明 Pi-ta 基因对黄淮稻区早熟品种(品系)的稻瘟病抗性发挥着重要作用。携带 Pi-b 基因品种(品系)的抗病比例约为20.0%,但检出率达到57.9%。分析仅携带 Pi-b 基因的17份材料可知,2年接种鉴定的抗性比例分别为0和17.6%,抗病性差。
2.4 水稻品种(品系)携带的抗病基因数量与穗颈瘟抗性相关分析
对145个水稻品种(品系)携带的抗瘟基因数量进行分析,由表2、表3可见,未检测出抗性基因的品种(品系)有9份,占总检测品种(品系)数的比例为6.2%,2年接种鉴定的抗性比例分别为0和11.1%,说明这部分材料的抗性较差。检测出润农Y-4等44个品种(品系)只携带1个抗性基因,占总检测品种(品系)的比例为30.3%。抗性基因 Pi-5 和 Pi-gm 未被单独检测到。检测出JD79等40个品种(品系)携带2个抗性基因,占总检测品种(品系)数的比例为27.6%,检测出基因型为 Pi-ta+Pi-5、Pi-b+Pi-54 和 Pi-b+Pi-gm 的品种(品系)各1个,连续2年接种抗性比例均为100.0%。检测出隆粳341054等23个品种(品系)携带3个抗性基因,占总检测品种(品系)数的比例为15.9%,其中4个品种(品系)基因型为 Pi-ta+Pi-54+Pi-km ,连续2年接种抗性比例分别为75.0%和50.0%,表现出较好的抗性。检测出新科稻31等19个品种(品系)携带4个抗性基因,占总检测品种(品系)数的比例为13.1%,其中基因型为 Pi-ta+ Pi-b+Pi-54+Pi-km、Pi-ta+Pi-b+Pi-54+Pi-5、Pi-ta+Pi-b+Pi-km+Pi-5 的品种(品系),2年接种鉴定的平均抗性比例均在60.0%及以上。回粳12的基因型为 Pi-b+Pi-54+Pi-km+Pi-gm ,GM-1的基因型为 Pi-b+Pi-km+Pi-5+Pi-gm ,这2个品种(品系)连续2年接种的抗性比例均为100.0%,表现出较强的抗病性。检测出连粳19116等10个品种(品系)携带5个抗性基因,占总检测品种(品系)数的比例为6.9%。其中连粳19116等9个品种(品系)的基因型为 Pi-ta+Pi-b+Pi-54+Pi-km+Pi-5 ,落育6的基因型为 Pi-b+ Pi-54+Pi-km+ Pi-5+ Pi-gm ,2年接种鉴定的抗性比例均为100.0%。
3 讨 论
2008年起,中国水稻新品种审定实行稻瘟病抗性的“一票否决”制,因为稻瘟病抗性不达标,每年国家区试中约40%的品种(品系)被淘汰 [23] 。本研究对145份黄淮稻区早熟品种进行了6个抗稻瘟病基因分子检测并连续2年进行穗颈瘟抗性鉴定。145份品种(品系)接种鉴定结果显示,穗颈瘟发病率2年分别达74.5%和68.3%,说明黄淮稻区早熟品种(品系)的稻瘟病抗性较差。大多数品种2年间接种鉴定结果差异较小,少部分品种(品系)由于菌种不同、接种时间以及环境等因素导致发病级别在年际间表现出较大差异。例如15JD278 在2019年接种鉴定为4级(高感),而在2020年接种鉴定为1级(抗病)。2020年在抽穗期间温度较高,不利于稻瘟病的诱发,整体抗瘟性好于2019年。
研究发现,携带 Pi-gm 基因的品种(品系)分布比例最低,仅有4.1%,但是6个携带 Pi-gm 基因的品种(品系)连续2年的抗性比例都达到了100.0%,表现出较强的抗病能力。李刚等 [24] 仅在谷梅 4号中检测出 Pi-gm 基因,该基因在年际间表现为高抗。潘争艳等 [25] 分析了260份辽宁省粳稻品种稻瘟病抗性基因,发现 Pi-gm 基因检出率为8.85%,携带 Pi-gm 基因单基因系的抗性比例为76.30%。进一步分析检测出 Pi-gm 基因的6个材料的来源,发现都来源于东北地区,其他地方品种资源中未检测出该基因,说明黄淮稻区早熟品种不含有 Pi-gm 基因。以上结果表明, Pi-gm 基因可能是改良黄淮稻区粳稻品种(品系)抗病能力的一个重要基因,但分布频率较低。检测到17个仅携带 Pi-b 基因的品种(品系)连续2年的抗病比例分别为0、17.6%,而 Pi-b 基因在黄淮稻区早熟品种(品系)中检出率最高,2年试验结果表明该基因的抗性比例非常低,分别为21.4%、26.2%,说明该基因对黄淮稻区早熟品种(品系)抗瘟水平的贡献非常小。张善磊等 [14] 的研究结果表明,只携带 Pi-b 基因的品种(品系)感病率高达 86.84%。刘艳等 [6] 对黄淮稻区水稻品种的穗颈瘟抗性进行分析,结果表明 Pi-b 检出率达到65%,但在黄淮海稻区的抗性水平较低。以上结果说明 Pi-b 抗性效应正在丧失。
辽粳401和天域125-3-3仅含有 Pi-ta 基因,连粳GP46仅含有 Pi-54 基因,这3个品种2年的接种鉴定结果都达到了1级(抗病)。检测到11个仅携带 Pi-ta 基因品种(品系)抗病比例为27.3%,而携带 Pi-ta + 其他抗性基因组合的品种(品系)2年的抗病比例分别为46.0%、52.0%,其中检测到3个品种(品系)抗性基因组合为 Pi-ta+Pi-b+Pi-km ,连续2年的抗病比例为100.0%,表现出稳定的抗病性,说明 Pi-ta 基因对黄淮稻区早熟品种稻瘟病的抗性发挥着重要作用, Pi-ta+Pi-b+Pi-km 基因组合可能是改良黄淮稻区早熟品种(品系)抗瘟性较好的基因组合模式。检测到7个仅携带 Pi-km 基因的品种连续2年的抗病比例为0,说明该基因可能已失去抗病能力。检测到9个仅携带 Pi-54 基因品种(品系)抗病比例仅为22.2%。本研究仅对6个基因进行检测,连粳GP46中是否含有其他基因还需进一步检测。 江苏省品种(品系)中 Pi-9、 Pi-zt 2个广谱抗病基因分布频率较高,下一步准备对 Pi-2、Pi-9、Pi-zt 等具有广谱抗性的基因进行检测。
研究结果表明,将稻瘟病抗性基因聚合到同一个品种中可以提升品种的抗病能力,但并不是聚合基因越多越好,而是应该选择抗性好、抗性互补的基因进行聚合。品种资源基因型分析结果显示,抗瘟基因 Pi-b 、 Pi-km、Pi-ta、Pi-54 和 Pi-5 的检出率分别为57.9%、51.0%、42.1%、32.4%及32.4%,抗性基因 Pi-gm 分布比例最低,仅有4.1%。笔者的前期研究结果表明 Pi-54 、 Pi-b 在黄淮稻区水稻品种(品系)中的检出率较高,分别达 84%、65% [6] 。本研究结果表明,黄淮稻区早熟品种(品系)的抗性基因比例偏低,这可能是早熟品种(品系)抗性较差的原因之一。本研究中有6个品种(品系)的基因型是 Pi-ta+Pi-b+Pi-km+Pi-5 ,连续2年的抗性比例都是66.7%,有9个品种(品系)的基因型是 Pi-ta+ Pi-b+Pi-54+Pi-km+Pi-5 ,2019年与2020年抗性比例分别是44.4%、55.6%,说明 Pi-54 与品种(品系)的抗病性可能存在负相关。这与王军等 [5] 认为 Pi-54 与穗颈瘟抗性呈负相关的结果一致。本研究的结果表明,4个基因聚合的平均病级最低,为2.3,但含有 Pi-ta+ Pi-b + Pi-km + Pi-54 + Pi-5 这5个抗病基因的品种(品系)2年间均出现4级(高感)的品种(品系),说明该基因组合抗病能力不足。总之,随着抗病基因数的增加,穗颈瘟接种鉴定级别有降低的趋势,但并非基因聚合越多,品种(品系)的抗性越强。
由于基因的多效性和连锁累赘效应 [26] ,在导入优良目标基因的同时,常常连带产生对生育期及产量等性状不利的影响,即存在一定负效应。向小娇等 [27] 研究结果表明,京作1号在导入基因 Pi-21 后,株型和综合性状相对较差,单株产量显著降低。Masao等 [28] 认为,稻瘟病抗性基因 Pi-zt 与晚熟基因紧密连锁。不同学者对 Pi-gm 的抗谱测定结果表明,该基因具有广谱抗性 [15,25] ,但对该基因负向效应的结论不尽相同。 Pi-gm 基因位于水稻第6染色体上,该条染色体短臂上有多个控制生育期、产量、品质的主效基因,例如遲抽穗基因 Hd1 就与 Pi-gm 连锁。田红刚等 [29] 的研究结果还表明, Pi-gm 基因的导入导致株高增加,穗粒数增加,结实率下降,有5个导入系产量显著提髙。张礼霞等 [30] 研究结果表明,导入 Pi-gm 基因的水稻品系生育期延迟,千粒质量增加,株高增加,有效穗数、每穗总粒数以及产量减少。向小娇 [27] 等研究结果表明, Pi-gm 基因能提升水稻品种的稻瘟病抗性,除了结实率显著下降外,产量及其他相关性状影响较小。Deng等 [31] 的研究结果表明, Pi-gm 对水稻的产量性状也存在影响,该基因能通过提升结实率、降低千粒质量平衡水稻产量结构,确保产量。笔者分析了千粒质量较高的连粳JD2、连粳JD3和千粒质量较低的连粳JD8、连粳15号与GM-1组配后代的千粒质量,结果表明千粒质量较高的连粳JD2、连粳JD3的后代千粒质量平均为28.1 g,而千粒质量较低的连粳JD8、连粳15号的后代千粒质量平均为23.5 g,育种实践表明,后代千粒质量的大小与选用的亲本有关,含有 Pi-gm 基因亲本配组后代可以选育出千粒质量不低于亲本的株系。
参考文献:
[1] 王宝祥,卢百关,潘启明,等.黄淮稻区水稻直播研究进展及育种策略探讨[J].北方农业学报,2017,45(5):1-5.
[2] 沈乐融,齐中强,杜 艳,等. 江苏省稻瘟病菌致病力分化及无毒基因组成分析[J]. 江苏农业学报,2019,35(1):42-47.
[3] 宋兆强,刘 艳,王宝祥,等.稻瘟病抗性基因 Pi-ta、Pi-b、Pi-54 和 Pi-km 的育种利用价值评价[J]. 江苏农业学报, 2017,33(5): 968-974.
[4] 王宝祥,刘 艳,邢运高,等.抗稻瘟病基因 Pi-ta、Pi-b、Pi-54 和 Pi-km 在黄淮稻区水稻品种资源中的抗性分布[J].植物遗传资源学报,2019,20(6):1465-1471.
[5] 王 军,宫丹妮,杨 杰,等. 江苏省粳稻品种抗稻瘟病基因型与穗颈瘟抗性分析[J]. 江苏农业学报,2016,32(2): 250-256.
[6] 刘 艳,王宝祥,杨 波,等. 黄淮海稻区水稻品种的穗颈瘟抗性[J]. 中国水稻科学,2019,33(4):377-382.
[7] 张 璐,何瑞银,沈有柏,等.江苏省主要水稻种植模式技术对比分析[J].中国农机化学报,2017,38(7):106-111,116.
[8] 陈长红,汪洪洋,孙克存,等. 苏北麦茬直播水稻栽培技术[J]. 现代农业科技,2007(13):151-154.
[9] 王 军,杨 杰,杨金欢,等. Pi-ta、Pi-b 基因在江苏粳稻穗颈瘟抗性育种中的价值分析[J]. 华北农学报,2012,27(6): 141-145.
[10] 王 军,杨 杰,朱金燕,等.稻瘟病抗病基因 Pi-k h 功能标记的开发及江苏粳稻品种中 Pi-k h 的变异[J]. 中国水稻科学,2014,28(2): 141-147.
[11] SHARMA T R, MADHAV M S, SINGH B K, et al.High- resolution mapping, cloning and molecular characterization of the Pi-kh gene of rice, which confers resistance to Magnaporthe grisea [J].Mol Genet Genomics, 2005, 274(6):569-578.
[12] ASHIKAWA I, HAYASHI N, YAMANE H, et al. Two adjacent nucleotide-binding site- leucine- rich repeat class genes are required to confer Pi-km specific rice blast resistance[J].Genetics,2008, 180(4):2267-2276.
[13] 陈 峰,徐建第,姜明松,等.黄淮区粳稻抗稻瘟病基因 Pi-ta、Pi-b、Pi-54、Pi-km 的分子检测[J].生物技术进展,2018,8(1):46-54.
[14] 张善磊,孙旭超,陈 涛,等. Pi-ta、Pi-5、Pi-km 和 Pi-b 基因在粳稻品种(系)中的分布及对穗颈瘟的抗性[J].江苏农业学报,2018,34(5):961-971.
[15] 王小秋,杜海波,陈夕军,等.江苏近年育成粳稻新品种/系的稻瘟病抗性基因及穗颈瘟抗性分析[J].中国水稻科学,2020,34(5):413-424.
[16] 刘永锋,陆 凡,陈志谊,等. 江苏省水稻新品种(系)对稻瘟病的抗性评价[J]. 中国水稻科学,2002,16(1): 96-98.
[17] 曾生元,李 闯,杜灿灿,等. Pi-gm 特异性选择标记的开发及其在粳稻穗颈瘟抗性育种中的利用[J].中国水稻科学,2018,32(5):453-461.
[18] 高利军,高汉亮,颜 群,等. 4个抗稻瘟病基因分子标记的建立及在水稻亲本中的分布[J]. 杂交水稻,2010,25(增刊1): 294-298.
[19] 王伟舵,刘永锋.中国稻瘟病菌遗传多样性研究进展[J].江苏农业科学,2016,44(6):196-198.
[20] 朱亚军,孙 强,王金明,等. 粳稻品种吉粳 809 的稻瘟病抗性基因分析[J].作物学报,2016,42(11):1638-1646.
[21] WU Y Y , XIAO N, Y U L, et al. Combination patterns of major R genes determine the level of resistance to the M. oryzae in rice ( Oryza sativa L.) [J]. PLoS One, 2014, 10(6): e0126130.
[22] 杨永义,张 华,宣 宁,等.利用分子标记辅助选择培育抗稻瘟病粳稻新品系[J].山东农业科学,2016,48(11):18-20,25.
[23] 王 洁,曾 波,雷财林,等.北方国家水稻区域试验近15年参试品种分析[J]. 作物杂志, 2018(1):71-76.
[24] 李 刚,袁彩勇,曹奎荣,等. 544 份水稻种质稻瘟病抗性鉴定及抗性基因的分布研究[J]. 中国农业大学学报,2018,23(5): 22-28.
[25] 潘争艳,邱福林,吕桂兰,等. 辽宁省粳稻品种稻瘟病抗性基因分析[J]. 中国水稻科学,2019,33(3):241-248.
[26] 刘敬贤,黄亚群,陈景堂,等. 基于高密度连锁图谱定位玉米株高QTL[J]. 江苏农业科学,2019,47(13):38-41.
[27] 向小娇,张 建,鄭天清,等.应用分子标记技术改良京作1号的稻瘟病抗性[J].植物遗传资源学报,2016,17(4):773-780.
[28] MASAO Y, HIROSHI F. Tight linkage of blast-resistance with late maturity observed in different indica varieties of rice [J]. Japanese Journal of Breeding, 1971,21(1):35-39.
[29] 田红刚,陈红旗,胡 江,等.抗稻瘟病基因 Pi-gm 导入对寒地粳稻抗病性和产量性状的影响[J].沈阳农业大学学报,2016,47(5):520-526.
[30] 张礼霞,王林友,范宏环,等.利用 Pi-gm 基因改良粳稻保持系的稻瘟病抗性研究[J].核农学报,2017,31(3):424-431.
[31] DENG Y W, ZHAI K R, XIE Z, et al. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance[J]. Science, 2017, 355(6328): 962-965.
(责任编辑:张震林)
