上海市草莓灰霉病菌对氟吡菌酰胺敏感性检测及抗性分子机制
2021-11-12高萍高士刚成玮等
高萍 高士刚 成玮等

中图分类号:S 481.4 文献标识码:A DOI:10.16688/j.zwbh.2020555
由灰葡萄孢Botrytis cinerea引起的灰霉病是一种难以防治的世界性的重要病害,可危害观赏植物、蔬菜、水果(例如草莓)等230余种作物。灰霉病菌可危害植物果实、叶片、茎叶和花等,该菌具有寄主范围广、产孢量大、遗传变异频繁、传播能力强等特点,所以非常难防治,且极易产生抗药性。草莓灰霉病主要发生在采果期,采后果实常迅速腐烂,造成非常严重的损失。
氟吡菌酰胺是由德国拜耳公司研发的一种琥珀酸脱氢酶抑制剂类(succinate dehydrogenase inhibi-tors,SDHIs)杀菌剂,2012年在我国获得登记。氟吡菌酰胺具有高效、广谱和持效久等特性,常用于防治霜霉病、灰霉病、白粉病和菌核病等。其通过抑制病原菌内呼吸链中琥珀酸脱氢酶活性,阻碍线粒体呼吸作用,从而抑制病原菌孢子萌发、菌丝体生长,进而控制病害发展。
上海市属亚热带海洋性季风气候,极适宜草莓灰霉病的发生,国内常用的酰胺类杀菌剂有啶酰菌胺,许多地区均已出现田间自然发生的灰霉病菌对啶酰菌胺產生抗性的报道,但关于草莓灰霉病菌对氟吡菌酰胺的抗性研究鲜有报道。因此,本研究通过氟吡菌酰胺对上海5个区的草莓灰霉病的抗药性检测,建立敏感性基线,研究抗性分子机制以及了解抗性分布,为未来氟吡菌酰胺的合理使用提供数据支持,为推动农药减量提供科学依据。
1材料与方法
1.1供试菌株
于2019年1月—3月,从上海市5个地区(崇明区、嘉定区、青浦区、奉贤区及浦东新区)的草莓生产基地,随机采集已感染发病的灰霉病病果,参考刘欣等的方法进行单孢分离、鉴定和保存,来源于同一病果的病菌保留1株,共获得90株单孢灰葡萄孢菌株。将所有菌株转接至PDA试管斜面,编号(地点缩写+序号),保存于4℃备用。
1.2药剂、试剂与培养基
98. 8%氟吡菌酰胺原药,南通泰禾化工股份有限公司提供(自制);丙酮,国药集团化学试剂有限公司;马铃薯葡萄糖琼脂培养基(PDA),青岛高科技工业园海博生物技术有限公司,用于菌株培养以及毒力测定;PCR EasyTaq Super Mix,北京全式金生物技术有限公司;琼脂糖,上海翊圣生物科技有限公司。
1.3菌丝生长抑制试验
采用菌丝生长速率法进行测定。氟吡菌酰胺原药用丙酮溶解,配成10000 μg/mL的母液,设置鉴别浓度分别为0、0. 25、0.5、1、2、4、8、16、32、64μg/mL及128μg/mL。将待测菌株接入PDA平板活化,于(25±1)℃黑暗培养72 h后,在菌落边缘打取直径为7mm的菌饼,接种到上述系列浓度的含药平板中央,以加入等量丙酮平板为对照,每处理重复3次,(25±1)℃黑暗倒置培养72 h后,用十字交叉法测量菌落直径。
抑制率=(对照菌落直径-处理菌落直径处理菌落直径)/(对照菌落直径-7)×100%;
抗性水平(RF)=菌株EC50/敏感基线EC50。
1.4琥珀酸脱氢酶基因序列测定及比对
菌株菌丝DNA的提取采用改良的CTAB法。根据灰葡萄孢琥珀酸脱氢酶基因(Sdh,A,Sdh,B,Sdh,C与Sdh, D)序列,采用刘欣等的4对引物:SdhA-F:ATTTGGAAACGCCCTTGGAC,SdhAR:CATTACCGAACAATCCCGCA;SdhBF:ACCTACTCGCCCTATCCAAT,SdhBR:AGACTTAGCAATAACCGCCC;SdhCF:GCCAGATTTCCTTAGTCAG,SdhCR:GCTGGACTCTGAATGTGAT;SdhDF:AGCCAATCAAATCCGTTCCG,SdhDR:CAAACTCCTCCCTGCCCTCT,对全部菌株的4条亚基编码基因进行PCR扩增。PCR 体系为20μL:包含2×PCR Easy犜犪狇Super Mix10μL、上下游引物各0.5μL(10 μmol/L)、DNA 模板20ng,最后用ddH2O补足。PCR程序:95℃预变性3min;95℃变性20s,55℃退火20s,72℃延伸30s,32个循环;72℃延伸5min。
取5μLPCR扩增产物于2%琼脂糖凝胶进行电泳检测,凝胶成像系统对胶片进行拍照记录,有条带PCR产物送生工生物工程(上海)公司进行测序。
1.5数据处理
利用Microsoft Excel软件计算药剂对菌丝生长的抑制率、有效抑制中浓度ECso值;利用SPSS22.0统计软件对试验数据进行分析,采用该软件描述统计模块进行Shapiro-Wilk法(W)法正态性检验,P>0. 05则符合正态分布,反之则为非正态分布;最后根据整个测定种群的ECso分布加以分析,绘制频率分布直方图及正态分布曲线。测序结果使用DNAMAN软件对敏感及抗性菌株的SDH亚基序列进行比对分析,并以NCBI网站(https:∥www. ncbi.nlm. nih. gov)数据库中相关序列作为参考。参考GenBank基因登记号:KR705916.1(BcSdhA)、KR866382.1(BcSdhB)、KR705923.1(BcSdhC)和KR705930.1(BcSdhD)。
2结果与分析
2.1草莓灰霉病菌对氟吡菌酰胺的敏感性基线
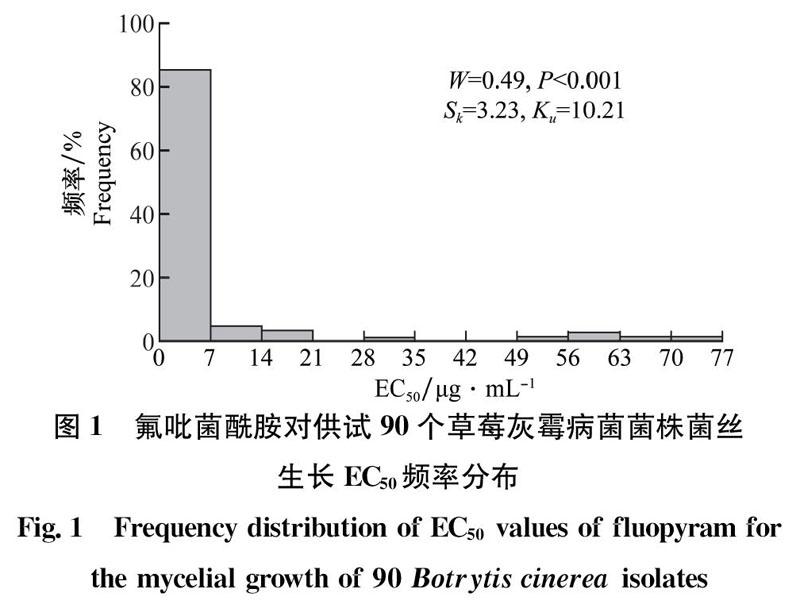
在上海市5个地区,共采集并成功分离到90个草莓灰霉病菌菌株。敏感性测定结果显示:氟吡菌酰胺对90个菌株的ECs。介于0.09~75.91μg/mL,表明上海不同地区之间草莓灰霉病菌对氟吡菌酰胺的敏感性差异较大。经Shapiro-Wilk正态性检验,结果显示:EC50频次分布W=0.49,P<0.001,呈非连续性分布(图1),表明不同菌株对氟吡菌酰胺的敏感性出现了分化,已出现敏感性降低的亚群体甚至抗性菌株。其中,22株菌株的EC50≥6.54μg/mL,呈非连续性分布(图1);其余68株菌株的ECo介于0.09~4. 80μg/mL,呈连续性正态分布,其W=0.97,P=0. 054>0.05(图2)。该68株菌株平均ECso为(1. 68±0.91)μg/mL,呈现一个连续单峰曲线,符合正态分布,故可作为敏感性基线参考值。
2.2菌株序列对比分析
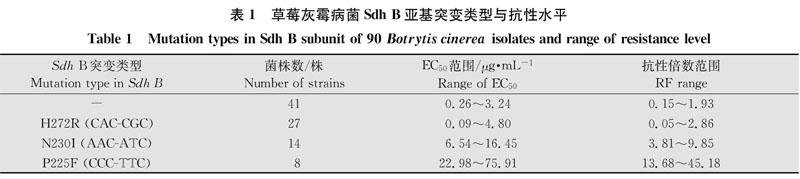
对全部90株菌株进行Sdh A,Sdh B,Sdh.C与Sdh D基因测序。结果显示:全部菌株的Sdh A、Sdh C与Sdh D 3个基因均未发生突变(结果未展示);Sdh B基因:41株菌株未发生突变,49株菌发生突。41株未突变的EG50范围为0. 26~3. 24μg/mL,27株272位组氨酸突变为精氨酸(CAC突变为CGC,H272R),菌株EC50范围为0.09~4.80μg/mL,与未突变菌株的EC50值存在交叉;14株230位天冬酰胺突变为异亮氨酸(AAC突变为ATC,N2301),菌株EC50范围为6. 54~16. 45μg/mL,8株225位脯氨酸突变为苯丙氨酸(CCC突变为TTC,P225F),菌株EC50范围为22. 98~75. 91μg/mL(表1)。
2.3上海地区草莓灰霉病菌对氟吡菌酰胺的抗性水平
参照表1的菌株突变类型、ECso和抗性水平范围,定义敏感性划分标准:RF<3为敏感(S);3≤RF<10为低抗菌株(LR);10≤RF<50为中抗菌株(MR);RF≥50为高抗菌株(HG)。根据上述划分标准,灰霉病菌Sdh B亚基未发生突变和272位突变的菌株属于敏感菌株;230位突变的菌株属于低抗菌株,225位突变的菌株属于中抗菌株(表1)。上海地区灰霉病菌菌株对氟吡菌酰胺的敏感性检测结果(表2)显示,全市抗性菌株频率24.44%,不同地区菌株的抗性频率差异较大,奉贤区抗性菌株频率最高,为77. 27%,浦东新区和嘉定区分别为25. 00%和4.76%,崇明区和青浦区未检测到抗性菌株,抗性频率均为0。不同地区菌株对氟吡菌酰胺的敏感性差异较大,奉贤区菌株出现最大ECo为75. 91μg/mL,而青浦区菌株出现最小EC50为0.09μg/mL,两者之间相差843倍(表2)。22个抗性菌株中低抗菌株占63. 63%、中抗占36. 36%,其中抗性最高的菌株抗性倍数达45. 18(表3)。
3结论与讨论
本研究采用菌丝生长速率法测定了2019年采自上海市5个草莓种植基地共计90株灰霉病菌株对氟吡菌酰胺的敏感性,其中的68株菌株EC50符合正态分布,所得曲线呈现一个连续单峰,均值为(1. 68±0.91)μg/mL,与张晓柯等针对江苏省草莓灰霉病菌建立的菌丝生长敏感基线(1. 94±1. 55)μg/mL接近,故可作为草莓灰霉病菌对氟吡菌酰胺的敏感基线参考值评估田间整体抗性情况。此外,结合菌株ECso和基因突变位点,对已报道的草莓灰霉病菌对氟吡菌酰胺抗药性标准(RF<2为敏感;2≤RF<10为低抗;10≤RF<50为中抗;RF≥50为高抗)进行了优化,新定义了更适用的草莓灰霉病菌对氟吡菌酰胺抗性划分标准(RF<3为敏感;3≤RF<10为低抗;10≤RF<50为中抗;RF≥50为高抗)。
研究表明,上海市5个区灰霉病菌对氟吡菌酰胺的敏感性已产生分化,全市灰霉病菌抗性菌株频率24. 44%,但是以低、中抗为主,而奉贤区抗性菌株频率已达较高水平,崇明、嘉定、浦东以及青浦草莓生产区草莓灰霉病对氟吡菌酰胺抗性频率较低,甚至为0,变异系数较小,说明这4个区对氟吡菌酰胺目前还处在敏感水平,未发现敏感性下降的群体。因此,在上海奉贤地区草莓灰霉病防治,应及时调整施药方案,减少氟吡菌酰胺的用藥频次,建议优先选用其他作用机制或类型的新型杀菌剂,如与咯菌腈等轮换使用,并密切监测病原菌对氟吡菌酰胺的敏感性变化动态,以避免防治失败。
本研究通过对供试菌株SDH基因的核苷酸序列的对比分析,发现Sdh A、Sdh C与Sdh D 3个基因均未发生突变,而Sdh B亚基272位密码子CAC突变为CGC,引起组氨酸(H)突变为精氨酸(R),但并不影响菌株的敏感性,这一结论与张晓柯等报道的抗性可能主要是由B亚基的272位突变引起不一致;230位密码子AAC突变为ATC,引起天冬酰胺(N)突变为异亮氨酸(I),菌株表现为低抗;225位密码子CCC突变为TTC,引起脯氨酸(P)突变为苯丙氨酸(F),菌株表现为中抗,这一结论与Amiri等的一致,Hu等和Fernandez等也报道草莓灰霉病菌株Sdh B的N230I和P225F突变导致其对氟吡菌酰胺产生抗性。
琥珀酸脱氢酶抑制剂类杀菌剂作用靶标为线粒体复合体Ⅱ(三羧酸循环过程中的关键复合体),通过完全或者部分占据底物泛醌的位点,抑制电子从琥珀酸传递到辅酶Q(泛醌),干扰呼吸链上复合体Ⅱ电子传递,阻断其能量代谢,最终导致病原菌死亡。结构包括3部分:酸片段(A)、胺片段(B)和酰胺键链接部分(C)(图3),C部分是固定基团,A和B部分为活性基团。啶酰菌胺和氟吡菌酰胺同为琥珀酸脱氢酶抑制剂类杀菌剂,具有不同的活性基团(图4)。许多研究发现灰霉菌Sdh B亚基的H272R突变可产生对啶酰菌胺的抗性,本研究发现H272R突变的灰霉菌株对氟吡菌酰胺仍旧敏感,两种杀菌剂对灰霉菌Sdh B亚基H272R突变表现出不同的生物活性。SDH杀菌剂的抑菌活性主要取决于它们与靶标位点的对接能力和亲和力,这种亲和力受到杀菌剂化学结构的高度影响。已有报道,氟吡菌酰胺的活性基团3-氯-5-三氟甲基2吡啶基被2-氯-4三氟甲基-2-苯基取代时,导致其抗子囊菌活性显著下降,而被正交苯基取代时,抗担子菌活性增强。
病原菌对SDHIs的抗性主要由琥珀酸脱氢酶复合体亚基上氨基酸突变所致,氨基酸突变,改变了药剂与药靶的亲和力,进而影响病菌的抗药性。Sdh B亚基230位点的天冬酰胺是亲水性的,异亮氨酸是高度疏水性,蛋白质疏水性的改变是否会影响琥珀酸脱氢酶的形成或氧化还原电位,从而有利于与其他氨基酸的对接形成Q位点(琥珀酸脱氢酶泛醌结合位点,quinone-binding sites)还有待进一步研究。同样,Sdh B亚基225位点的脯氨酸被体积较大的苯丙氨酸替代,使得225位点附近的空间构型发生改变,从而降低了杀菌剂结合Q位点的亲和性。本研究发现Sdh B亚基N2301和P225F突变与氟吡菌酰胺抗性程度高低具有相关性,为将来草莓灰霉病菌对氟吡菌酰胺的抗性检测技术的研究奠定了一定的理论基础。此外,由于本研究抗性菌株群体数量的局限性,上海市草莓灰霉病病菌抗氟吡菌酰胺的菌群是否还存在其他基因突变类型,以及氟吡菌酰胺与其他药剂交互抗性关联性等方面有待进一步研究。
