混播草种在植被混凝土上群落构建技术应用研究
2021-11-12胡蝶费永俊张洋
胡蝶,费永俊, 张洋
长江大学园艺园林学院,湖北 荆州 434025
种群的空间格局分析是研究种群特征、种群间相互作用以及种群与环境关系的重要手段,一直是生态学中研究的热点之一[1]。一般植物群在群落中的分布有3种类型,即随机分布、集群分布和均匀分布[2]。种群的分布特征是种群关系和环境条件的综合影响所决定的[3]。在一定意义上,种群的格局是对环境适应和选择的结果,因而种群空间分布格局通常反映一定环境因子对个体行为﹑生存和生长的影响。
在草坪学中,根据草坪草生长发育对环境温度的要求,将其分为冷季型和暖季型2大类[4,5]。在冷、暖季草坪过渡的中间地区,冷、暖季型草坪草都能生长,但不论是种植冷季型还是暖季型都不理想,暖季型草有绿期短、不易越冬的问题,冷季型草则有夏枯、休眠甚至死亡的现象。因此,在该地区如何将冷、暖地型草种合理搭配进行混合混种,互相弥补生长季节的差异以解决夏枯、越冬问题,使草坪四季如春,成为我国未来草坪业发展技术研究的一个热点与难点。
植被混凝土作为近几年发展起来的新型环保建筑材料,既具备普通混凝土的特性,又满足一定的生态需求,其最大的特点就是具有透水、透气等植生性有利于植物生长[6],并且具有较强的抗冲刷能力与护坡性能[7]。因此对于植被混凝土的研究也多集中在“性能与应用”[8,9]、“ ‘植被混凝土’的外界抗性与配置”[10,11]、“边坡绿化与植被修复”[12,13]等方面。而针对植被混凝土草坪建植中冷暖草种的混播组合筛选是一个新的研究方向,迄今鲜有报道。植被混凝土调配材料含有水泥使植物基质呈碱性环境,且混播草种的不同生物学特性导致种间互作等,直接影响植被混凝土草坪建植质量。因此,要实现高质量的砼护坡混播草坪,首先要研究混播草种的配方,其次是了解各混播草种在植被混凝土上的适应性表现,如种群的空间分布格局以及群落内部草种间的竞争共存等。下面,笔者对普通未脱壳狗牙根(Cynodondactylon)、日本结缕草(ZoysiajaponicaSteud)、公园早熟禾(Poaannua‘Park’)、正义高羊茅(Festucaelata‘Justice’)4种草种在植被混凝土上进行混播试验, 并研究各种群的空间分布格局,目的在于揭示该混播组合中各个种群的分布规律,探讨其分布与环境的关系以及种间的竞争与共存关系,对不同混播比例的植被混凝土草坪进行质量评价,研究在植被混凝土中这4种冷、暖草种混播的可行性,同时找出一种适于江汉平原建植的、能够提高草坪草在混凝土环境下整体抗逆性和有利于边坡植被修复的冷暖草种搭配方案,从而为荆江大堤的生态固防提供便捷的技术支持。
1 材料与方法
1.1 试验地自然条件
试验地点选在位于我国长江中下游地区的湖北省荆州市长江大学园艺园林学院设施园艺基地。试验地点所在地位于东经111°15′~144°05′,北纬29°26′~31°37′,地处湖北省中南部、江汉平原腹地,海拔35m。毗邻长江,地下水位高。该地域土壤由近代河流冲积物形成,以水稻土、潮土、黄棕壤为主体,土层深厚肥沃,pH 7.15~8.15,呈石灰性反应。该地区属北亚热带季风气候区,光能充足,年日照时间1400~1600h。无霜期长,全年无霜期242~263d。气候温和,全年平均气温为15.9~16.6℃,年有效积温(≥10℃)为5000~5350℃。雨量充沛,且雨热同季,年降水量1100~1300mm。该地区4~10月份降水量占全年80%,太阳辐射量占全年75%,有效积温(≥10℃)占全年80%。试验所在地区的气候条件适合许多草坪草种的生长[14]。
1.2 试验材料及来源
供试草种为4种护坡绿化草种:普通未脱壳狗牙根、日本结缕草、公园早熟禾、正义高羊茅,由上海春茵种业有限公司提供。植被混凝土由黄沙壤土、水泥等材料混合而成,沙壤土∶水泥∶其他(化肥、添加剂等)的质量比为77∶9∶14。
1.3 试验方法
1.3.1 草种的选择
2018年12月2日取各供试草种的若干品种在长江大学园艺园林学院设施园艺基地播种,浇足水后覆以地膜,待其出苗后揭去。于2019年2月22日、3月17日和3月25日分3次测定各品种的适应性指标,包括密度、色泽、质地、成坪性、枯萎度、枯萎率等。用“九分制”评分法[15]评定各品种的综合表现,进而筛选出普通未脱壳狗牙根、日本结缕草、公园早熟禾和正义高羊茅4种适宜荆州地区生存环境的最优草种。
1.3.2 植被混凝土的制作与播种方法
2019年3月28日制作植被混凝土并播种。方法如下:将按植被混凝土配方调制好的基质加水调和成泥浆状,然后用40cm×40cm×10cm的木质方格作模,先将调好的泥浆状基质倒入其中约9cm高度后,然后将按比例混匀的草种和倒入剩下1cm模具中的泥浆搅匀,倒入模具中,自然流平。重复一次待干燥取出木质模具,覆以遮阳网,每日浇水3~4次,待全部出苗后揭去遮阳网。
1.3.3 草种混播比例
各冷季型和暖季型草种的混播组合见表1。
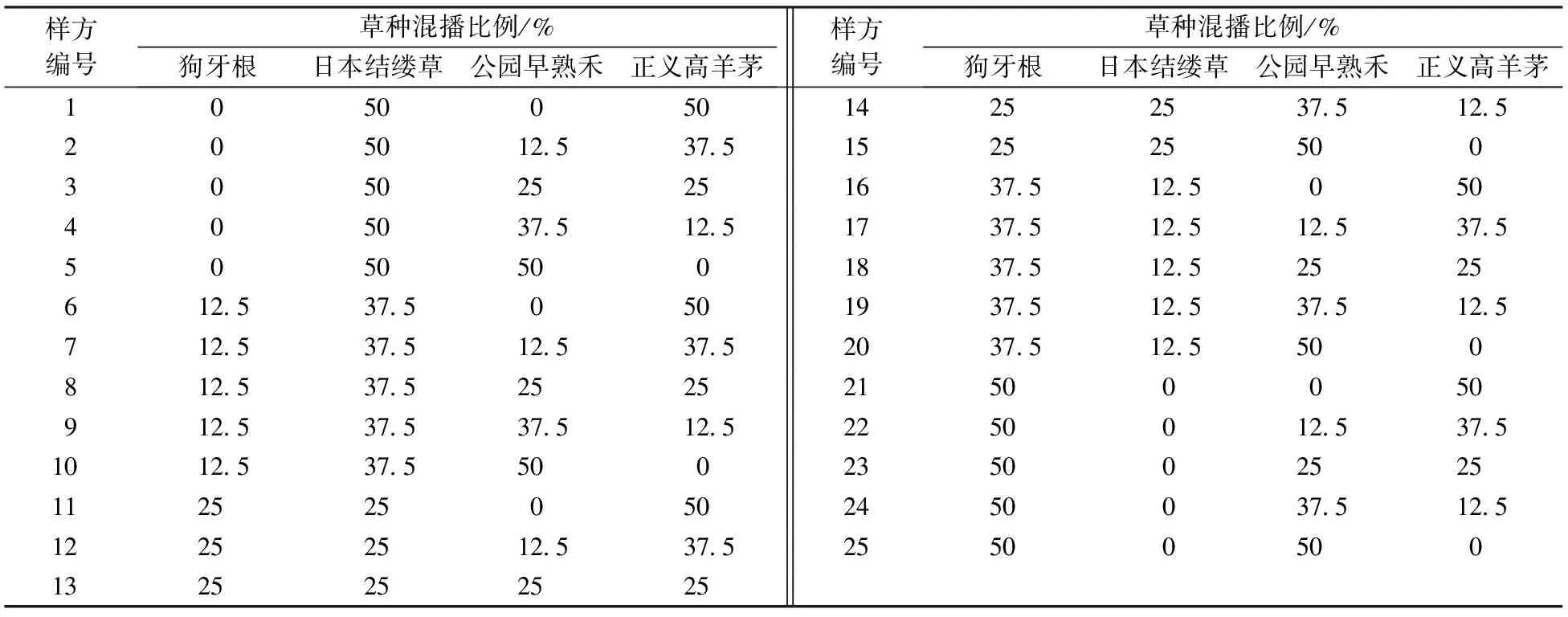
表1 草种混合比例表
1.3.4 取样方法
以每一砼块为一个区,以各个区左下角为原点沿混凝土边沿设置20cm×20cm的样方,并以每个样方的左下角为坐标原点记录数据。在每个样方内用目测法估测各种群的相对多度、相对盖度[16]。
2 结果与分析
2.1 各样方上混播草种的分布点图
利用Excel表格将采集到的各样方内各草种的坐标制表,由此绘制各样方内草种分布的点阵图(见图1)。在绘图时将所测数据按0.05的比例缩放尺寸绘制。由图1可以清晰、直观地看出各草种在植被混凝土上的分布格局。
2.2 各草种播种比例与其相对多度的相关性
2.2.1 狗牙根播种比例与其相对多度的相关性
狗牙根的相对多度随播种比例的增加而增大。在冷季型草高羊茅播种量的相对优势区,如样方7、9、12、14中狗牙根的相对多度随播种量的增加而增大的趋势较为明显;而在高羊茅和早熟禾各自播种量的绝对优势区,如样方6、10、11、15中,狗牙根的相对多度随播种量的增加而增大的趋势较为平缓。尤其是在高羊茅的绝对播种区,如样方6、11,由于该样方中的混播的冷季型草种只有高羊茅,狗牙根的生长受到抑制。在样方8、13、18、23中,由于高羊茅和早熟禾的播种比例相同,同季型草种间的竞争较为激烈,狗牙根则因此获得了生存发展的样方空间,其相对多度增长也最快(见图2)。
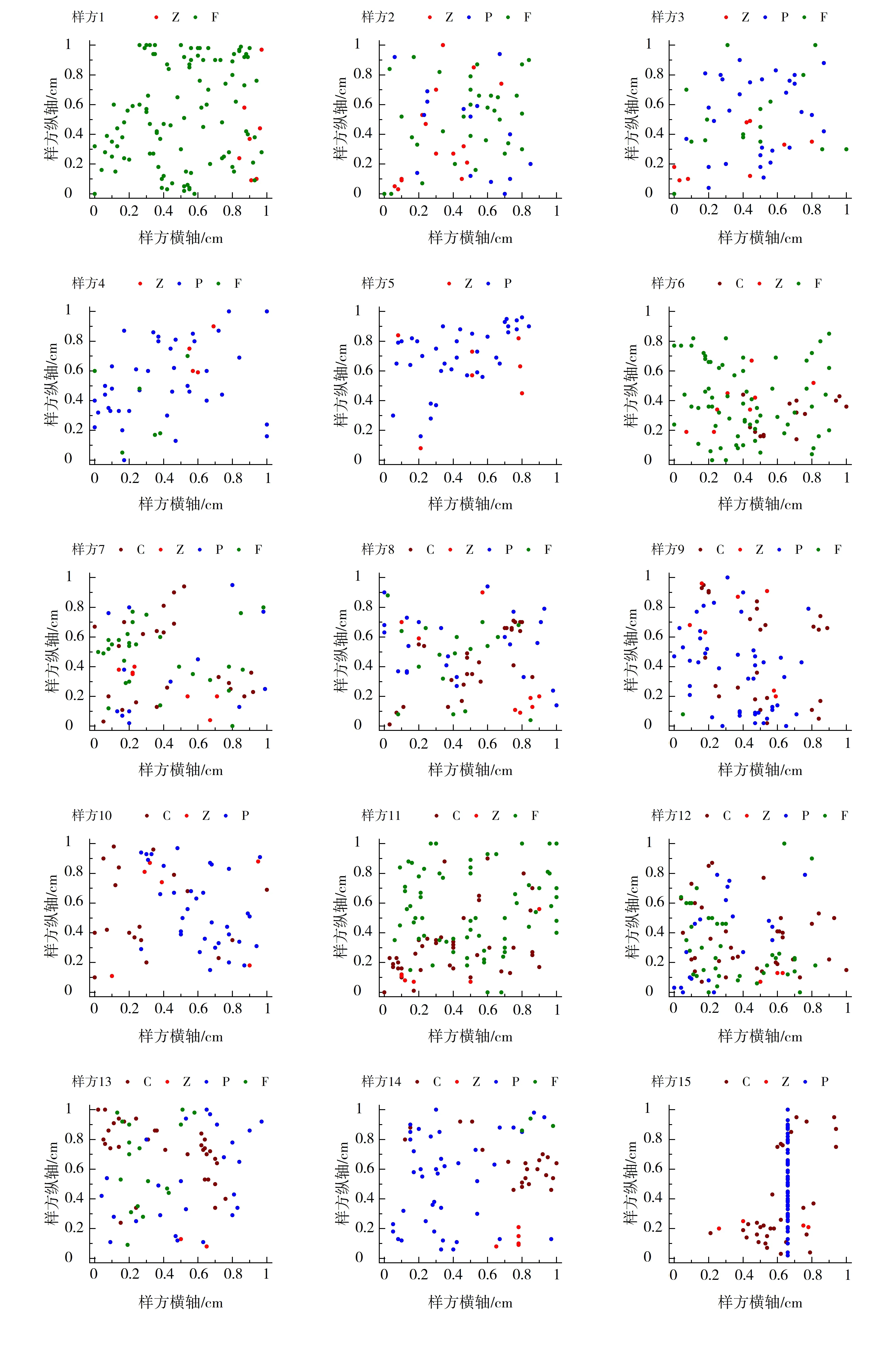
注:图例中大写字母C、Z、P、F 分别表示普通未脱壳狗牙根、日本结缕草、公园早熟禾和正义高羊茅。
图1 各样方内草种分布点阵图
Fig.1 Dot plot of distribution of grass species in each sample plot
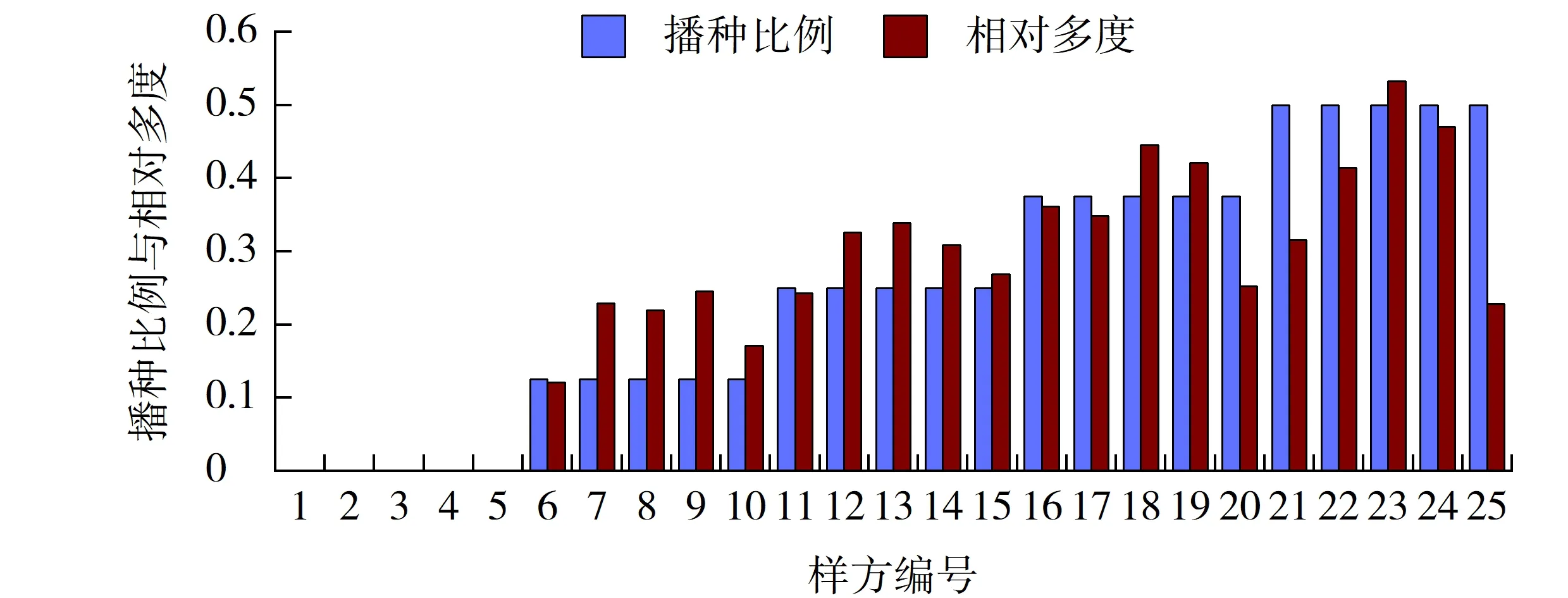
图2 狗牙根播种比例与其相对多度的相关性Fig.2 Correlation between sowing proportion and its relative abundance of Cynodon dactylon
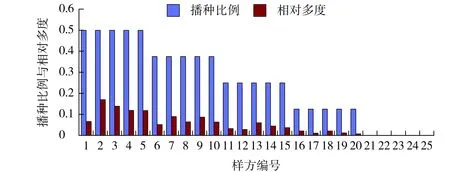
图3 结缕草播种比例与其相对多度的相关性Fig.3 Correlation between sowing proportion and its relative abundance of Zoysia japonica
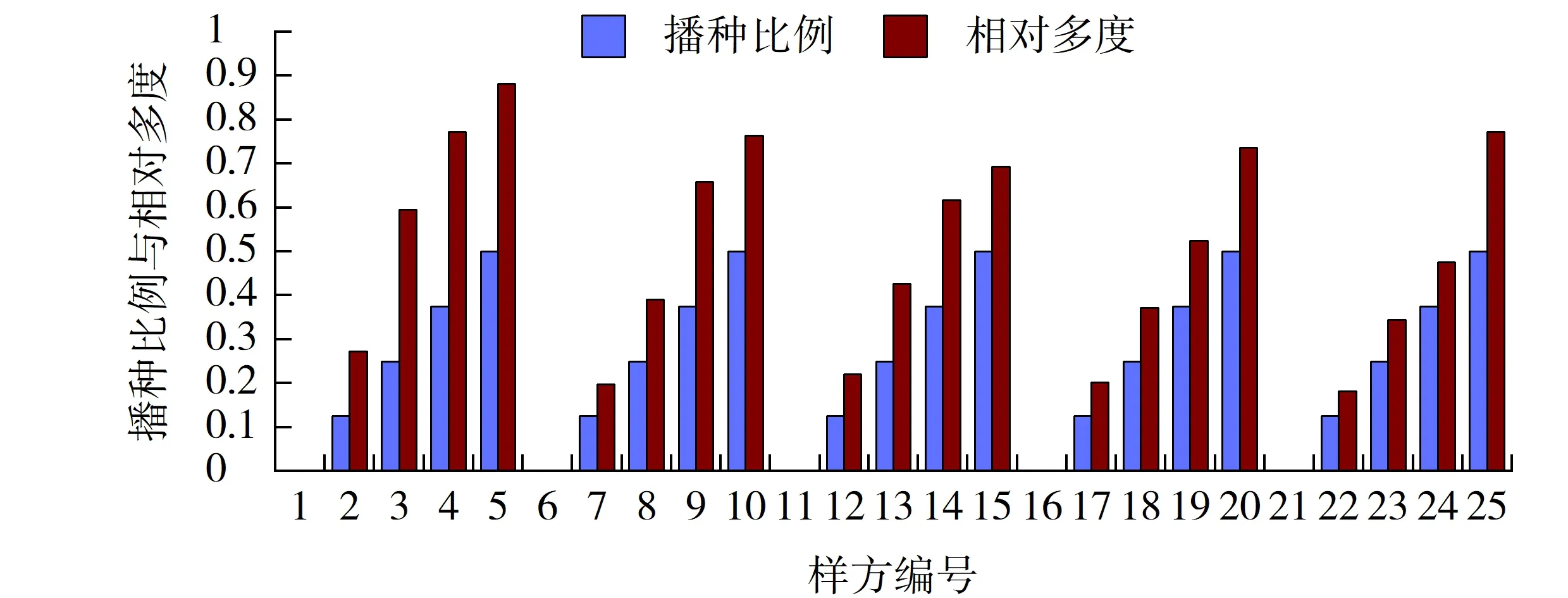
图4 早熟禾播种比例与其相对多度的相关性Fig.4 Correlation between sowing proportion and its relative abundance of Poa pratensis
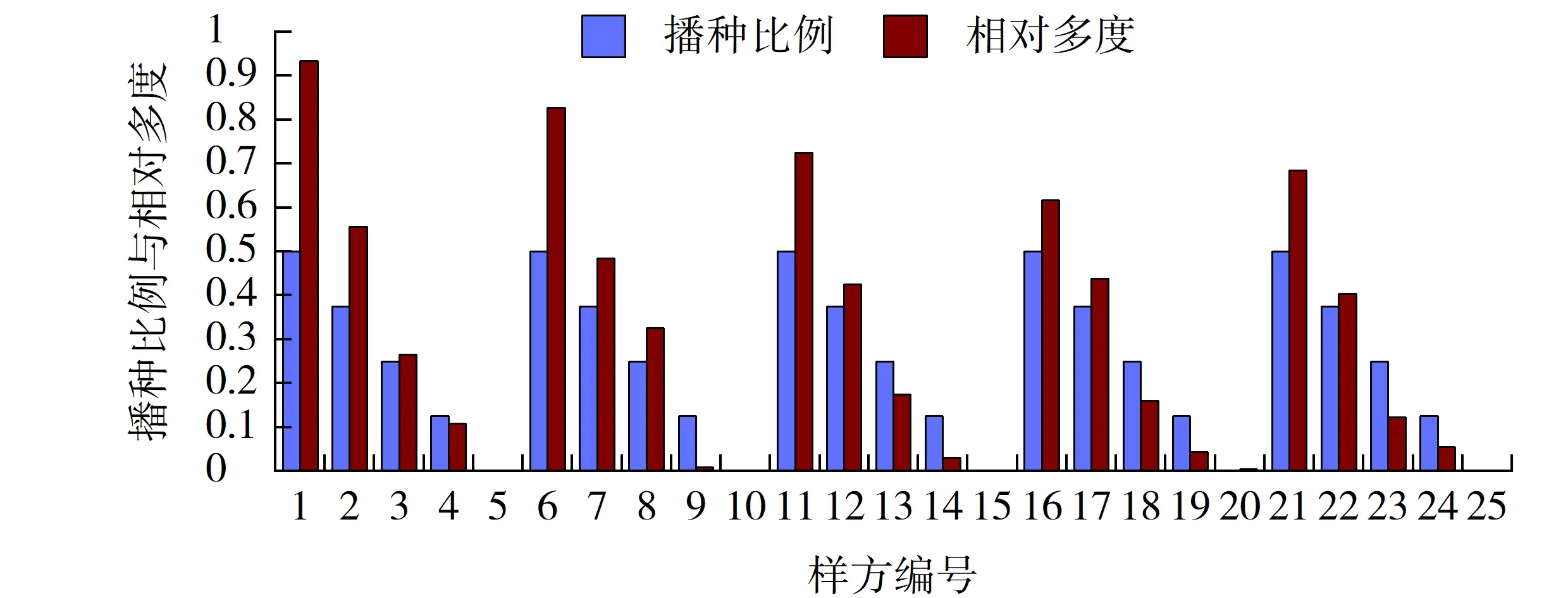
图5 高羊茅播种比例与其相对多度的相关性Fig.5 Correlation between sowing proportion and its relative abundance of Festuca arundinacea
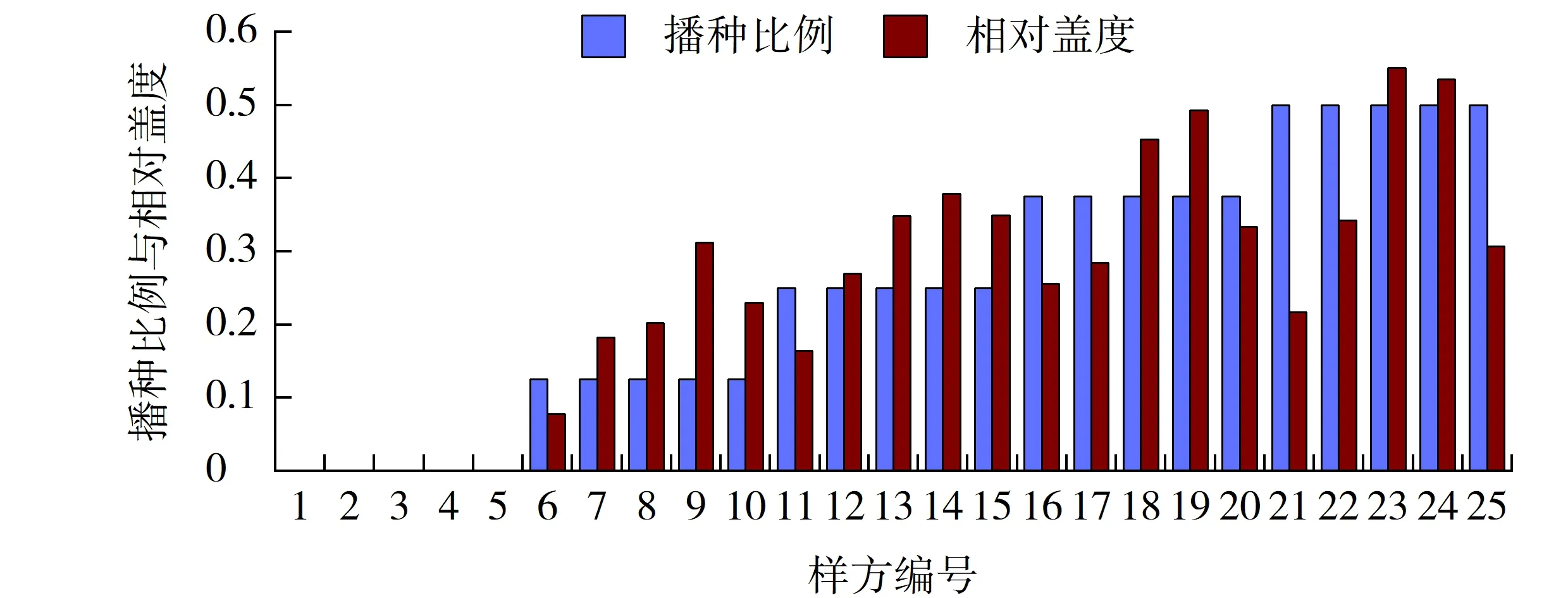
图6 狗牙根播种比例与其相对盖度的相关性Fig.6 Correlation between sowing proportion and its relative coverage of Cynodon dactylon
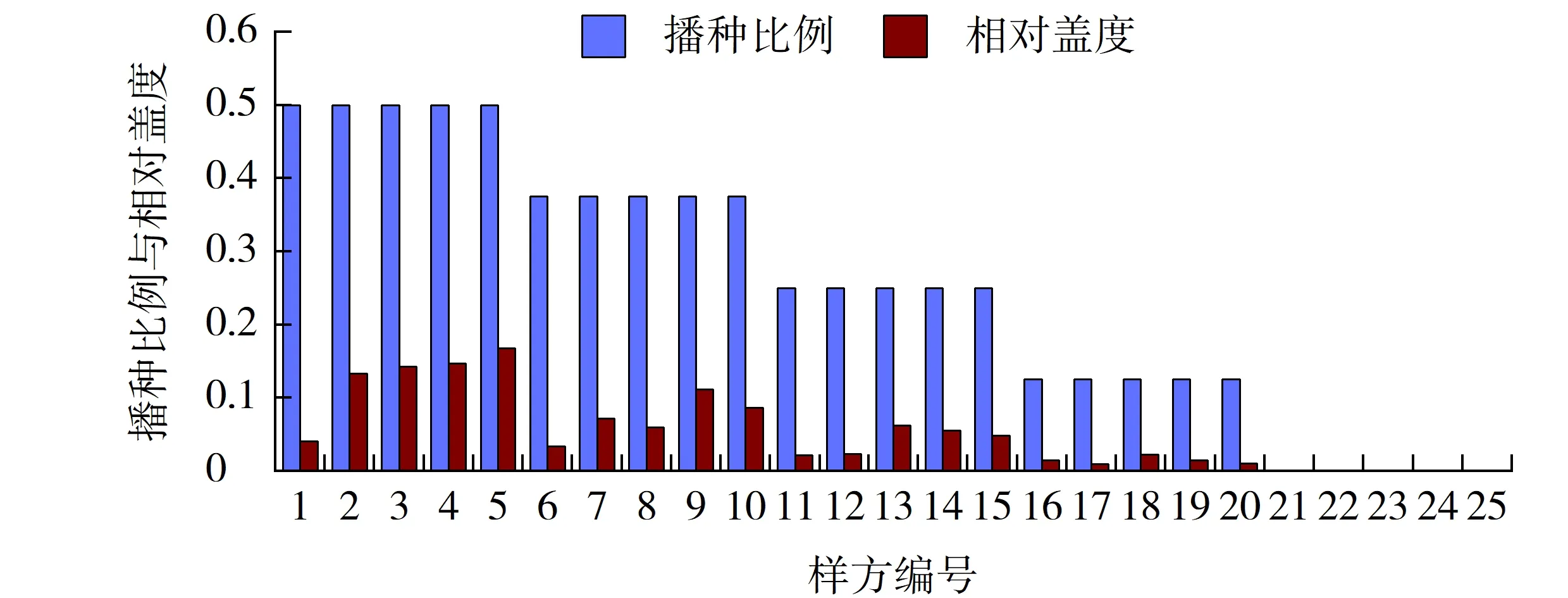
图7 结缕草播种比例与其相对盖度的相关性Fig.7 Correlation between sowing proportion and its relative coverage of Zoysia japonica
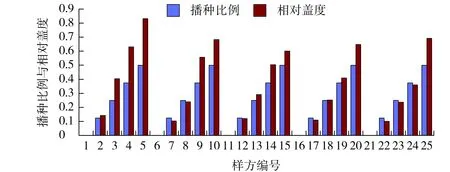
图8 早熟禾播种比例与其相对盖度的相关性Fig.8 Correlation between sowing proportion and its relative coverage of Poa pratensis
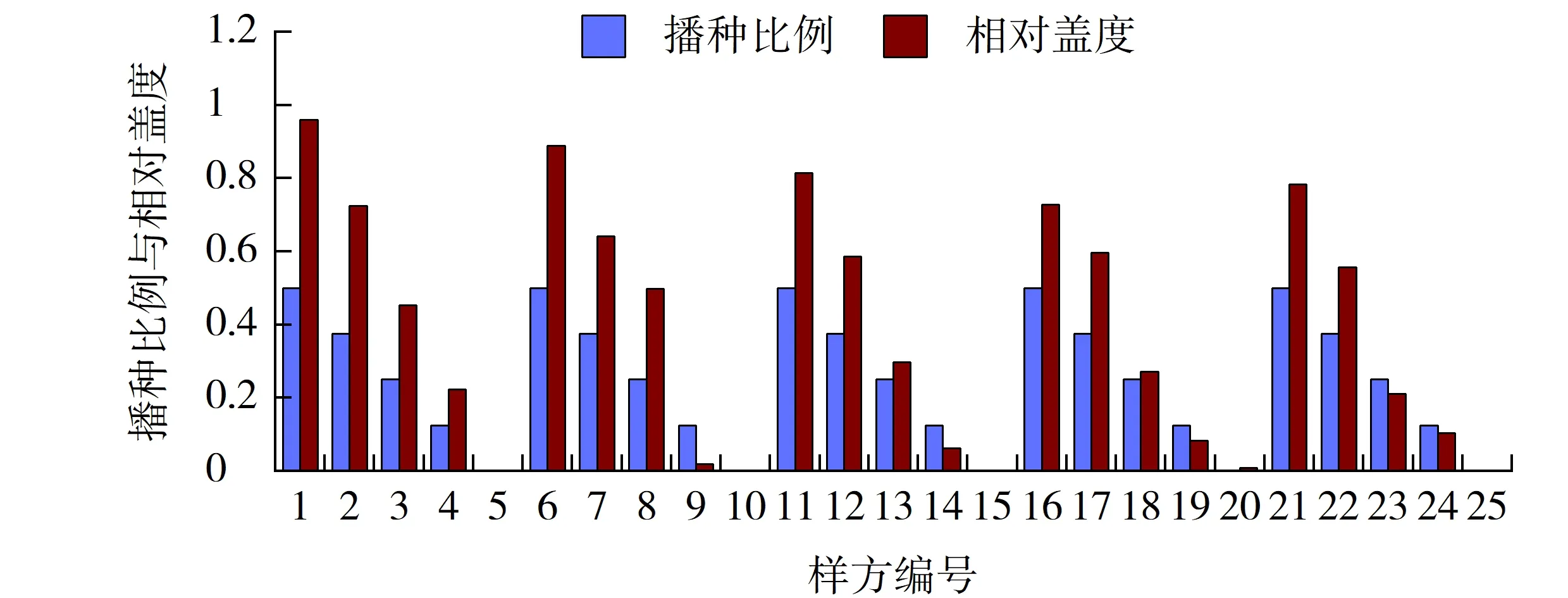
图9 高羊茅播种比例与其相对盖度的相关性Fig.9 Correlation between sowing proportion and its relative coverage of Festuca arundinacea
2.2.2 结缕草播种比例与其相对多度的相关性
日本结缕草播种比例与其相对多度相关性的最大特点是其相对多度远小于其播种比例。一方面,结缕草需要承受同季型草种狗牙根的生存竞争压力,而狗牙根作为过渡区的乡土种,其适应性所产生的竞争优势要优于结缕草;另一方面,结缕草还需要承受来自高羊茅和早熟禾对自然资源利用方面的压力。高羊茅的粗生性和早熟禾生长的致密性都抑制了结缕草的分布空间。此外,结缕草发芽和成坪较慢是此草种的重要特性之一,表现为早期的发芽障碍(见图3)。
2.2.3 早熟禾播种比例与其相对多度的相关性
各样方上的早熟禾的相对多度均要高于其播种比例。在结缕草播种量的绝对优势区,如1~5样方中,在结缕草的调解下早熟禾在与高羊茅的竞争中占据了优势,成为该样方中的绝对优势种。随着狗牙根播种比例的增加,这种优势逐渐减弱,其增加趋势也变得缓慢(见图4)。
2.2.4 高羊茅播种比例与其相对多度的相关性
高羊茅的相对多度随着播种比例变化而变化的规律较为复杂,在样方1~5中,混播的暖季型草种只有结缕草,高羊茅的相对多度随播种比例的降低而减小的幅度较大;其后,随着结缕草播种比例的降低其减小的趋势也渐趋平缓。但与早熟禾不同的是,并非所有的样方上高羊茅的相对多度都要高于其播种比例。在样方4、9、14、19、24中,其相对多度均要小于播种比例,说明高羊茅在早熟禾播种量的相对优势区,与早熟禾的竞争处于劣势(见图5)。
2.3 各草种播种比例与其相对盖度的相关性
至采集相对盖度数据时,高羊茅的分蘖数为2~3,狗牙根为1~2,早熟禾未分蘖。
2.3.1 狗牙根播种比例与其相对盖度的相关性
狗牙根的相对盖度在样方6、11、16、21中低于其播种比例,而在样方10、15中却要高于其混播比例。上述两组样方分别为高羊茅和早熟禾的播种绝对优势区。在样方7、8、12、13中,由于强分蘖性的高羊茅挤占了早熟禾的生存空间,故狗牙根的相对盖度要高于其播种比例,但在暖季型草种只剩下狗牙根时,早熟禾的生长致密性抢占了更多的生存空间,在相对多度上占据了绝对优势,所以狗牙根的相对盖度相应减小(见图6)。
2.3.2 结缕草播种比例与其相对盖度的相关性
日本结缕草的相对盖度均在20%以下,且与狗牙根相同,在高羊茅的优势区相对盖度较小,而在早熟禾的优势区相对盖度较大,这与其在跟狗牙根竞争中的劣势、相对多度过小以及体形较小有关(见图7)。
2.3.3 早熟禾播种比例与其相对盖度的相关性
早熟禾在有结缕草的样方中,其相对盖度较大,随着狗牙根的播种比例加大,其相对盖度相应减小。如在样方4、9、14 中,由于结缕草与狗牙根的播种比例分别为4∶0、3∶1、2∶2,早熟禾利用相对多度的优势使得其相对盖度高于其播种比例。而在样方19和24中,其相对盖度比播种比例的优势微弱,甚至更小(见图8)。
2.3.4 高羊茅播种比例与其相对盖度的相关性
在有高羊茅分布的植被混凝土上,除样方4、9、14、19、24外,其相对盖度均要高于其播种比例,一方面源于其相对多度优势,另一方面则因为其强分蘖性和簇生性。在上述5个样方中则由于高羊茅相比于早熟禾的播种比例过小致使其相对盖度比播种比例略小(见图9)。
3 讨论与结论
草种的空间分布格局,决定于种群的生物学特性和环境条件以及相互作用,是由种群特性、种内关系和环境条件的综合作用所决定的[17]。在某种意义上,它与环境条件的相关成因果关系,或者说种群的空间分布格局是对环境适应的结果,因而种群格局通常反映着一定环境因子对个体行为生存和生长的影响。
蒙古冰草种群在不同生境下,随尺度增大由集聚分布逐渐过渡到随机分布和均匀分布[1];南荻(Triarrhenalutarioriparia)随着生长由均匀分布变为聚集分布,且不同高程、不同土壤下种群分布格局具有明显差异[18];空灵山不同生境下的油松(Pinustabulaeformis)均呈集聚分布[19];黄背草(Themedajaponica)种群在河南各区域空间分布均为聚集分布[20]。以上报道均说明生境与种群构成对于种群分布格局型态有一定影响,这与本研究中由于高羊茅的簇生性和早熟禾生长的致密性,使得暖季草种在其零星分布区呈现聚集分布态势的结果相符。在阳光下,光合强度高的植株与光合强度低的植株争夺阳光和水分,强壮的个体形成了优势个体,发育缓慢的个体出现衰弱或死亡。在该研究中,冷暖混播的结果表明,4种草在植被混泥土上均为聚集分布,且在高羊茅草种播种比例较大的植被混凝土上,暖季型草种均不同程度地受到来自高羊茅的生存压力的作用,这与孙傲[21]针对混播草种研究中提到的在含有高羊茅的混播配方中高羊茅为优势种的结果相符。
刘杰等[22]对3种草坪草的测评中结缕草的坪用价值较高,这与本研究中结缕草的不适性有着较大差异。可能的原因是混播草种的种间竞争与植被混凝土环境劣于自然生境,表现为早期萌芽障碍,而过渡区的土壤与气候环境较为特殊,结缕草面临越冬问题。植被混凝土的碱性环境使各草种的发芽情况和长势均劣于自然生境,由此可推断,混播草种种群的这种分布格局还与种子在碱性条件下的发芽率等有关。
依据各草种的生物学特性,结合该研究中混播草种在植被混凝土碱性环境中的表现以及最终所需建植草坪的功能,所选择的草种配方并不是唯一的。但根据本研究目的,即主要应用场景和价值是植被恢复和边坡绿化,综合冷暖季草种在不同混播配比样方中相对多度与相对盖度的分析最终得出样方17(狗牙根37.5%+结缕草12.5%+早熟禾12.5%+高羊茅37.5%)混播效果最为理想,其次是样方22(狗牙根50%+早熟禾12.5%+高羊茅37.5%)。由于其中结缕草不适应混泥土混播环境,将样方17中的结缕草去掉,得到最适宜的混播配比为高羊茅∶狗牙根∶早熟禾=4∶3.5∶2.5。
根据生态学理论,遵循种群的分布规律,选择合适的植被混凝土配方,设计适当的草种混播组合和播种量,将会提高草坪的整体质量。由于生物因子主导了草种的分布格局,因此建议加强对混播草种配方、种群、群落以及物种多样性相关的多学科综合的生态学和系统生态学的研究。
