沙鞭不同居群染色体数目及核型分析
2021-11-09王亚男刘玉萍刘雪利刘瑞娟
王亚男,刘玉萍,刘雪利,苏 旭,3*,刘瑞娟
(1 青海师范大学 生命科学学院,西宁 810008;2 青海师范大学 青海省青藏高原药用动植物资源重点实验室,西宁 810008;3 高原科学与可持续发展研究院,西宁 810016;4 中国科学院西北高原生物研究所,青海省作物分子育种重点实验室,西宁 810008)
染色体作为细胞遗传信息的载体,其数目、结构和形态受外界环境影响较小[1],在植物生长发育和繁殖过程中具有较高的稳定性。染色体核型分析主要是对分裂中期染色体的数目、形态和结构等重要性状进行观察,计算和判别染色体的长度、着丝点位置、长短臂比例及随体有无等特征[2],进而推断不同物种或同一物种不同居群间的亲缘关系[3]。现今,染色体核型分析已成为物种分类修订、系统进化关系研究的重要手段[3]。先前诸多学者[4-7]对大量禾本科植物的染色体核型进行了报道。阎贵兴等[4]通过对33种禾本科饲用植物染色体核型的研究,发现染色体基数均为7,染色体类型有中部着丝粒和近中部着丝粒染色体,多数禾本科植物的核型对称性较高;张同林等[5]基于小麦族5个近缘属植物染色体核型的分析,推断了它们的系统发育关系,认为赖草属植物的进化程度最低,鹅观草属植物的进化程度最高,披碱草属植物的演化水平居于两者之间;郭玉堂等[6]通过对芨芨草(Achnatherumsplendens)染色体核型的探讨,结果发现其染色体数目为2n = 48,核型类别为2B;李伟伟等[7]利用染色体压片技术对沙鞭(Psammochloavillosa)的染色体数目进行了观察,表明该物种的染色体数目为2n = 40。
沙鞭是禾本科(Poaceae)沙鞭属(Psammochloa)一种多年生根茎类草本植物,主要分布于中国内蒙古、陕西、宁夏、甘肃及青海等沙地和荒漠化生境中[8]。沙鞭生长于海拔900 ~ 2 900 m的流动沙丘和固定沙丘上[8],根状茎发达,具有较强的耐旱、耐寒、耐碱及抗风沙功能[9],是内蒙古及其毗邻沙化地区的重要优势植物;同时,该物种营养价值较高,是畜牧业的重要牧草,具有重要的经济价值和生态价值。迄今,已有学者对沙鞭的外部形态特征[10]、叶表皮微形态特征[11]、种子生物学特性[12]、等位酶变异和克隆多样性[13]、地理分布和种质资源[14]、群体遗传多样性[15]等方面进行了研究。然而,从居群水平上对沙鞭染色体数目、核型及亲缘关系的研究目前尚未见报道。因此,本研究采用常规染色体制片技术,通过对沙鞭不同居群染色体数目和核型的综合分析,系统探讨它们的核型差异及系统亲缘关系,旨在为今后沙鞭群体遗传学、基因组学和进化生物学研究提供细胞学方面的参考依据。
1 材料和方法
1.1 实验材料
实验材料为沙鞭不同居群的成熟种子,采自于内蒙古阿拉善盟、鄂尔多斯和锡林郭勒盟,凭证标本保存于中国科学院西北高原生物研究所青藏高原生物标本馆(HNWP),详细采样信息见表1。
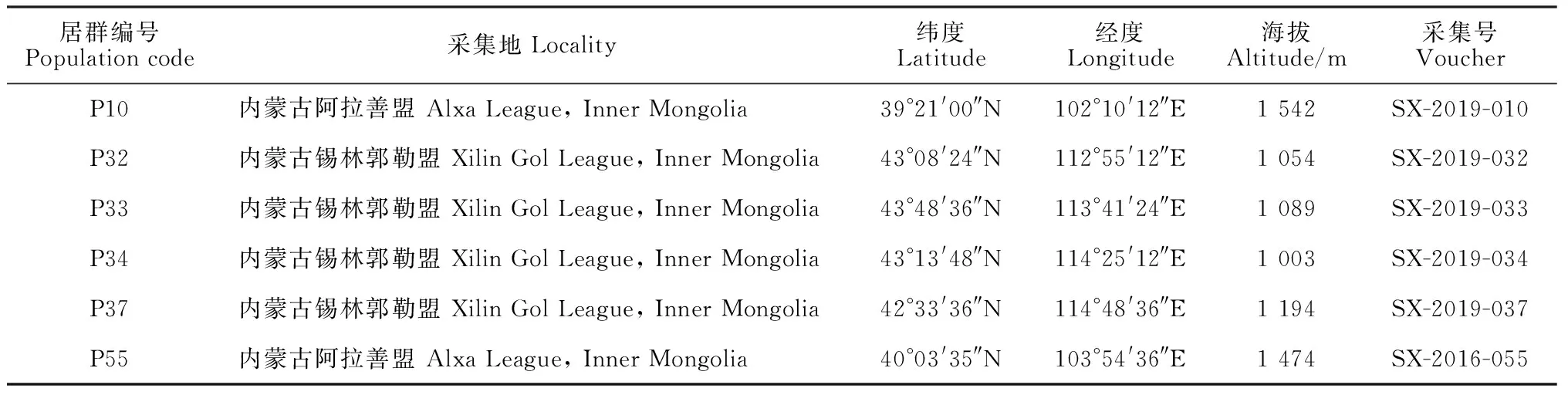
表1 实验材料及来源
1.2 方 法
1.2.1 种子萌发沙鞭每个参试居群挑选10个个体的成熟饱满种子,-20 ℃冰箱内处理24 h;然后将种子均匀放置于铺有蒸馏水润湿的双层滤纸培养皿中[16],22 ℃恒温培养箱中培养,待根长出0.5~1 cm时取样进行预处理。
1.2.2 预处理将根尖放于预先用蒸馏水喷湿的2.0 mL离心管中,并使离心管内外保持湿润,然后置于密封罐中N2O处理130 min,压强设置为0.75 MPa。
1.2.3 固定与解离从密封罐中取出离心管,迅速加入卡诺氏固定液(冰乙酸∶无水乙醇=1∶3,现配现用),随后轻轻翻滚离心管,使固定液充分覆盖于根尖表面,4 ℃冰箱中固定10 min以上,45%乙酸解离5 min。
1.2.4 染色、压片与观察将适当长度的根尖分生组织置于载玻片中央,滴2滴45%乙酸,盖上盖玻片用镊子钝端敲击,并在酒精灯上微烤,大拇指轻按盖玻片后置于相差显微镜(Olympus BX43)下观察;挑选分散良好的制片标记,然后放入-80 ℃冰箱中处理2 h以上,取出后荧光染液染色,用荧光显微镜(Olympus BX53)对染色体形态和数目进行观察,并挑选分散良好、形态清晰的视野拍照[17]。
1.2.5 核型数据分析每个沙鞭参试居群统计30个完整的染色体中期分裂图像,参照邓爱辉等[18]方法确定染色体数目;利用Adobe Photoshop 2021软件测量染色体长臂和短臂的长度[19];常规方法统计染色体相对长度(RL)、长度比(CLR)、臂比(AR)等参数;核型分析依据李懋学[20]方法;相对长度系数(I.R.L)依据Kuo等[21]方法;核型不对称系数(As.K%)依据Arano[22]方法;核型分类依据Stebbins[23]方法。
1.2.6 核型进化趋势和聚类分析核型进化趋势分析参照邢世岩等[24]方法,利用Microsoft Excel 2010软件绘制沙鞭核型不对称性程度散点图,即核型二维进化图;聚类分析依据李晓莉等[25]方法,利用SPSS 26软件统计沙鞭染色体臂比平均值(MAR)和方差、相对长度方差、染色体长度比(CLR)和正中部着丝粒(M)、中部着丝粒(m)、近中部着丝粒(sm)或近端部着丝粒(st)染色体比例等参数,并对其进行聚类分析。
2 结果与分析
2.1 沙鞭不同居群染色体数目和倍性
通过对沙鞭不同居群细胞中期分裂相的观察和计数,发现沙鞭居群的染色体均为二倍体,数目为2n = 2x = 46,同时未见异常的染色体数目和形态(图1),表明沙鞭染色体的数目较为稳定。
2.2 沙鞭不同居群染色体形态特征和类型
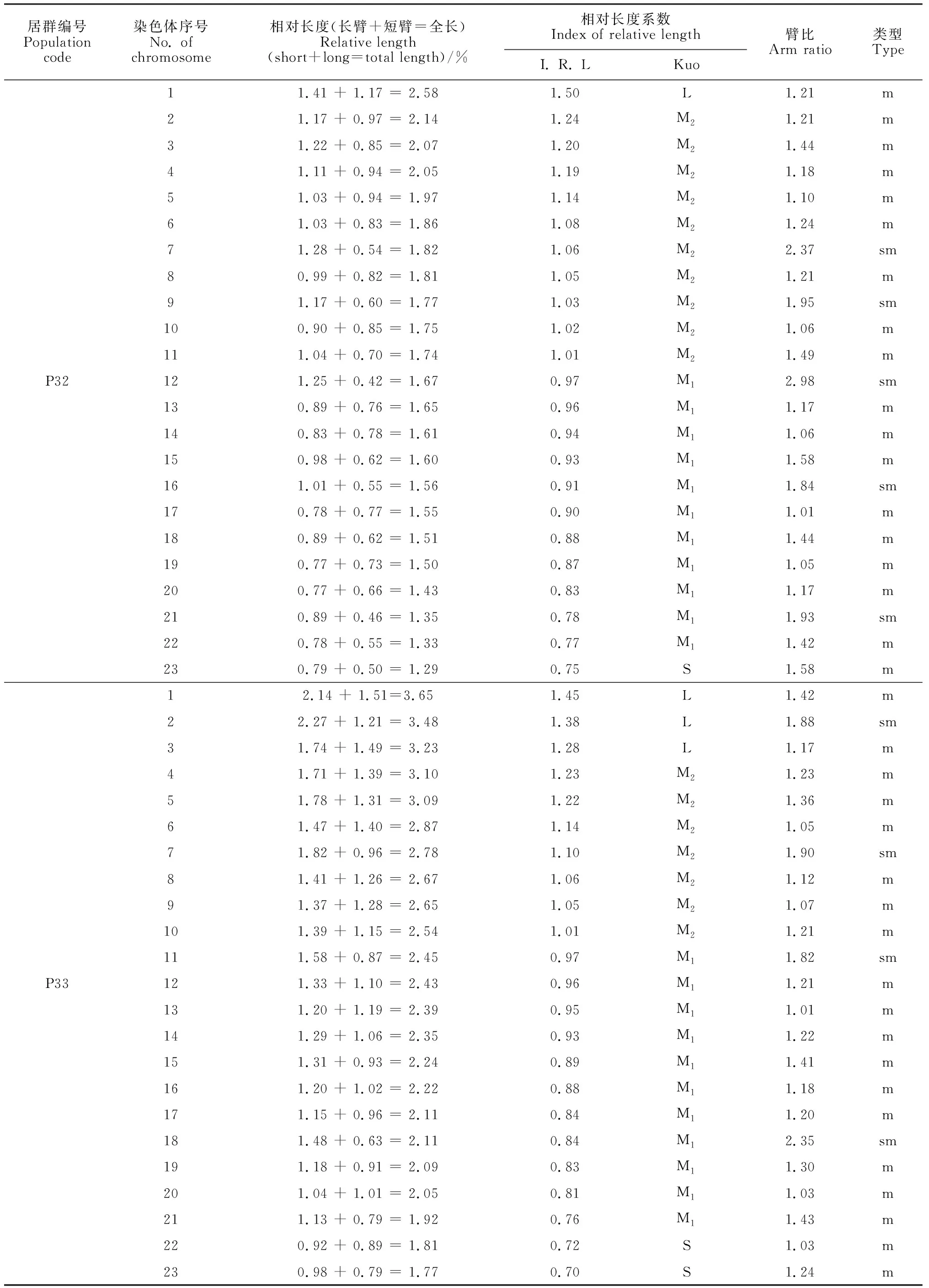
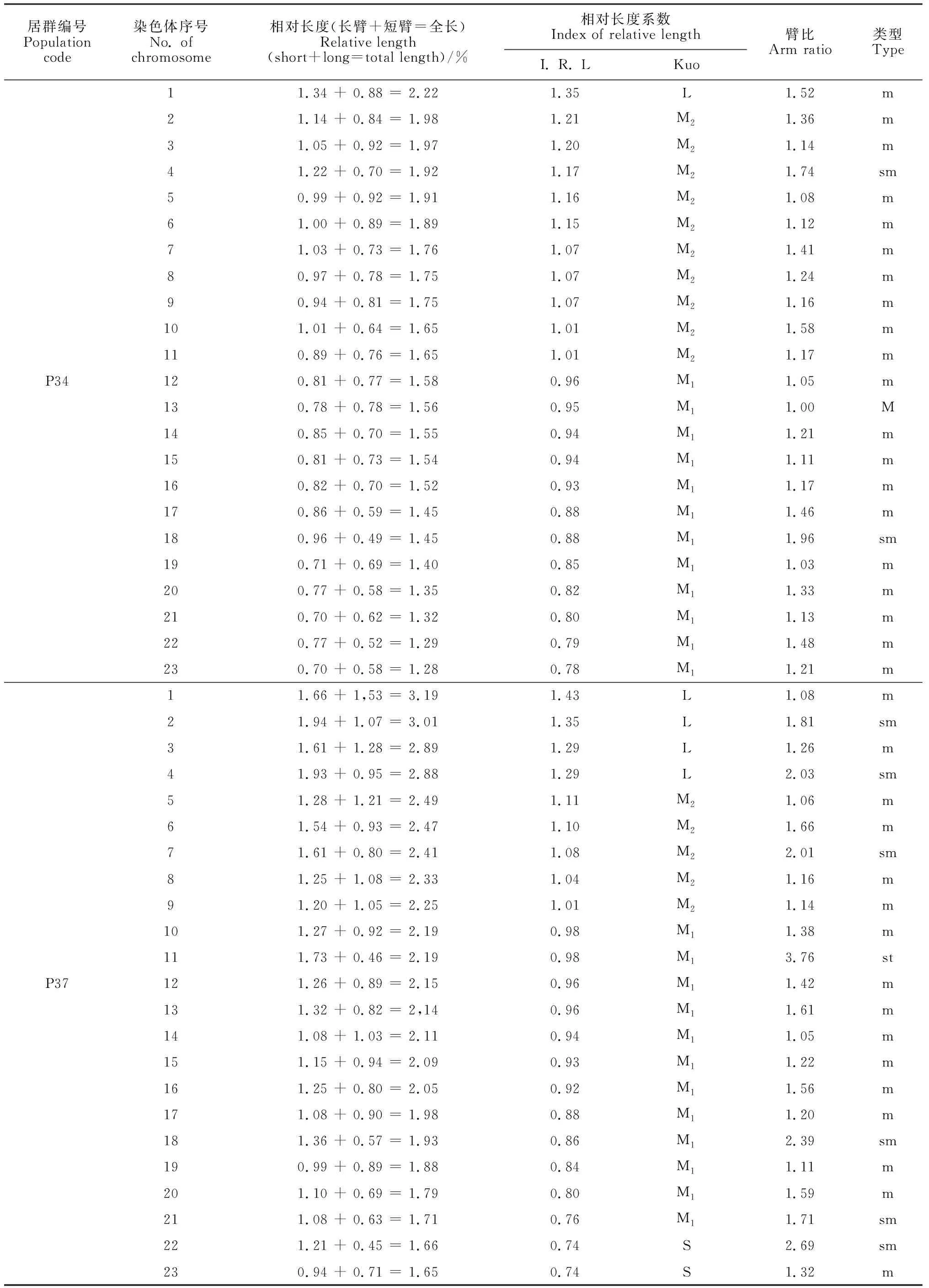
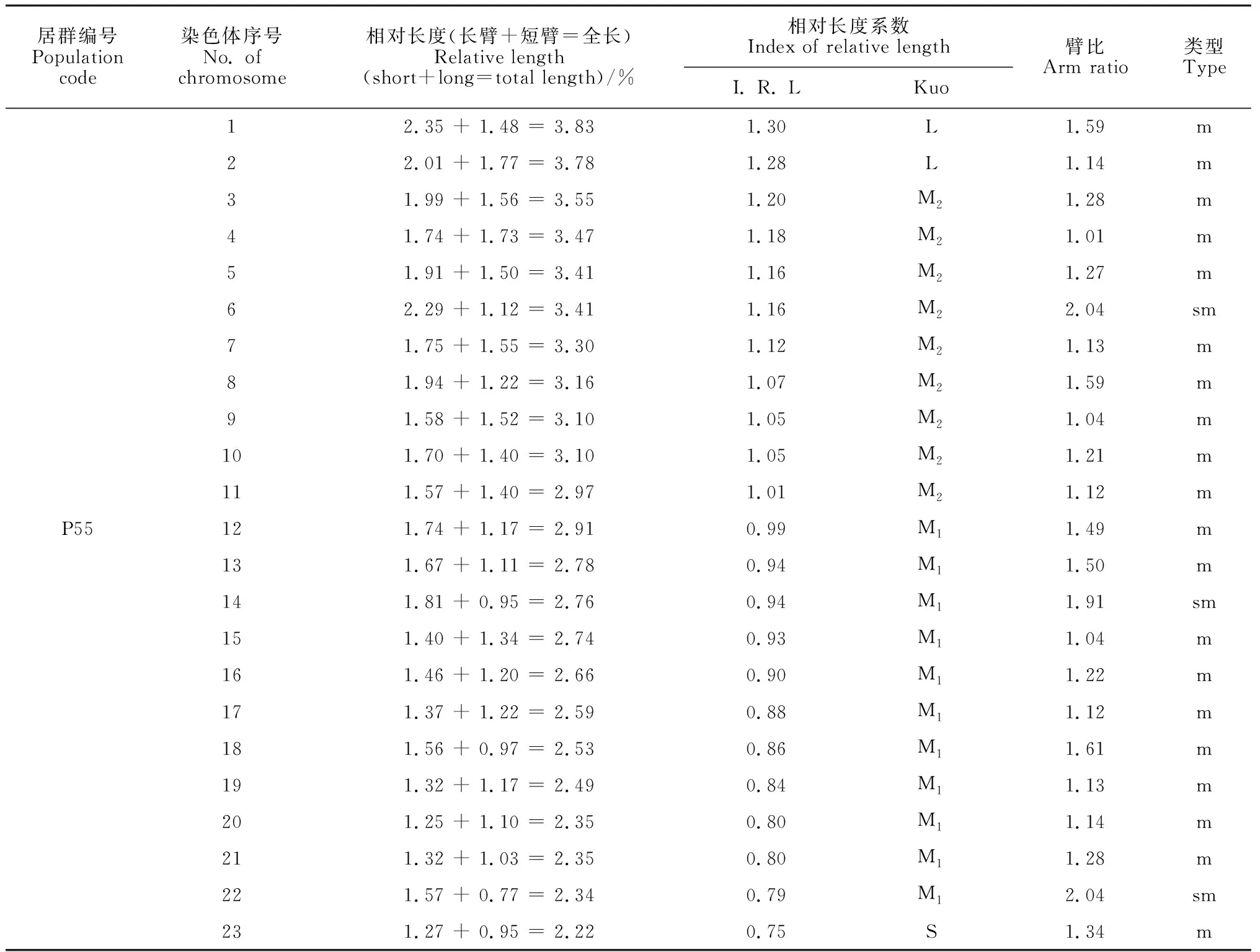
选取沙鞭分散良好的染色体有丝分裂中期图,结合表2中染色体参数和位置,将46条染色体进行同源染色体配对,共23对染色体。臂比统计结果显示,沙鞭染色体有M、m、sm和st 4种类型,其中34居群有M、m和sm 3种类型, 37居群有m、sm和st 3种类型,其余居群只有m和sm 2种类型(表2);并且,本研究还发现沙鞭不同居群中各类型染色体所占比例相差较大(表2)。基于沙鞭染色体相对长度系数及Kuo[21]分类标准,本研究中沙鞭不同居群染色体可划分为长染色体L型(≥1.26)、中长染色体M2型(1.01 ~ 1.25)、中短染色体M1型(0.76 ~ 1.00)和短染色体S型(< 0.76)4种类型;除沙鞭34居群无S型染色体外,其他所有居群均具以上4种类型染色体(表2)。
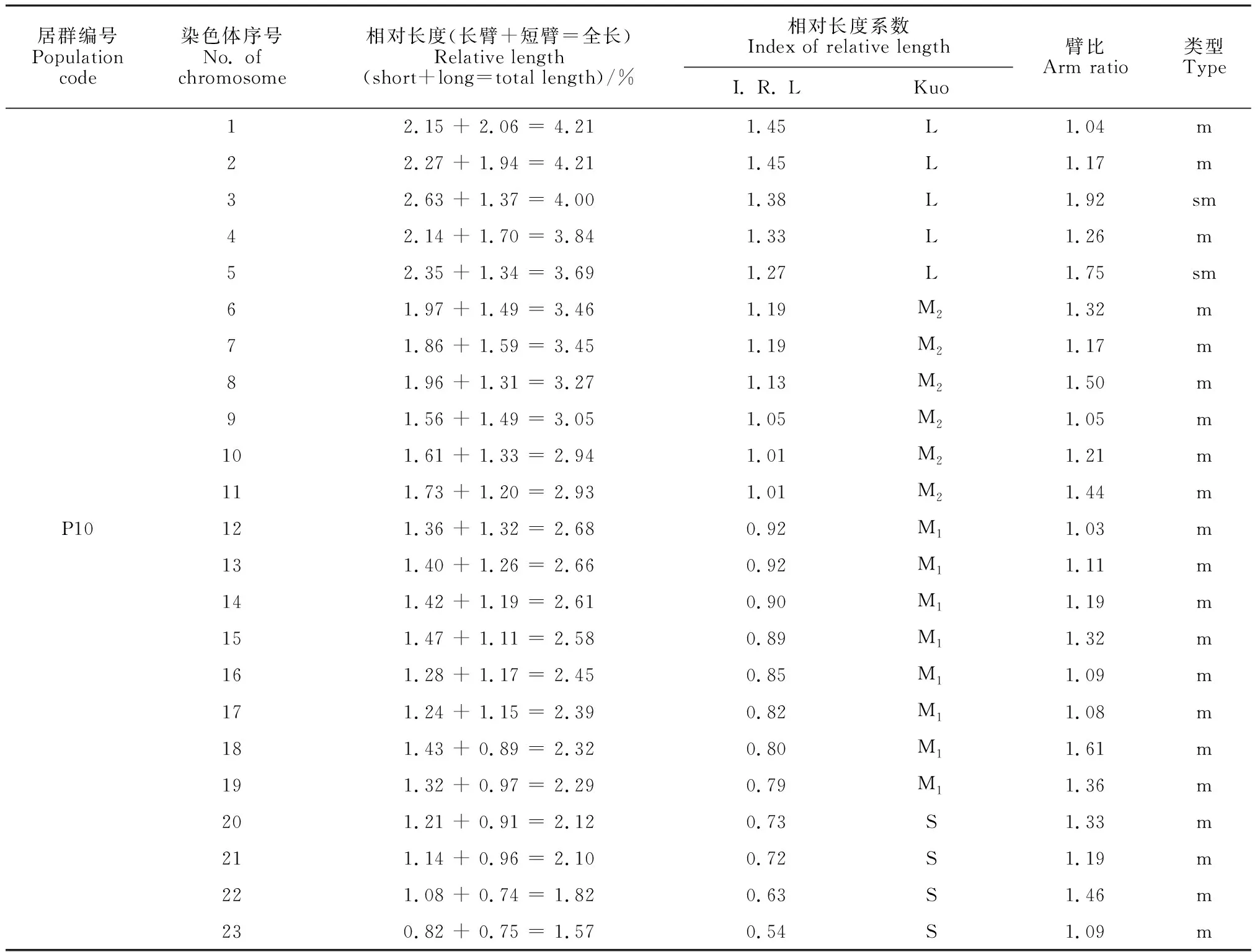
表2 沙鞭不同居群染色体参数

A.P10; B. P32; C. P33; D. P34; E. P37; F. P55图1 沙鞭有丝分裂中期染色体图Fig.1 Mitotic chromosomes in metaphase of P. villosa
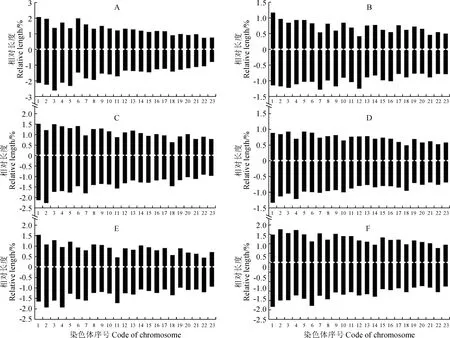
A. P10; B. P32; C. P33; D. P34; E. P37; F. P55图2 沙鞭不同居群的染色体核型模式图Fig.2 Chromosome idiograms in different populations of P. villosa
2.3 沙鞭不同居群染色体核型分析
沙鞭不同居群染色体核型模式图和主要核型参数见图2和表3。本研究结果表明,沙鞭不同居群在染色体核型公式、核型类型、染色体长度比、平均臂比及核型不对称系数等指标均存在明显差异。就核型类型来看,沙鞭居群染色体核型类型有1A、2A、1B和2B型,其中37和55居群的染色体核型类型为2A型, 32和33居群的染色体核型类型属于2B型,而34和10居群的染色体核型类型分别隶属于1A型和1B型(表3);就沙鞭染色体长度比而言, 10居群的长度比最大(2.68),而34和55居群的长度比最小(1.73)(表3);同样,在染色体平均臂比上, 37居群的平均臂比最大(1.62),10和34居群的平均臂比最小(1.29)(表3);就核型不对称系数来说,沙鞭34居群的核型不对称系数最小(55.96%),表明核型对称性最佳,而37居群的核型不对称系数最大(59.95%),说明核型对称性最差(表3)。
2.4 沙鞭不同居群染色体核型进化趋势分析
通过对沙鞭居群染色体长度比和平均臂比统计分析,本研究发现沙鞭染色体核型呈现“双向进化趋势”,即有些居群沿染色体平均臂比方向进化较快,如37居群;而另一些居群则沿染色体长度比方向进化较快,如10居群(图3,A)。就沙鞭居群总体进化趋势而言,二维图右上方的居群通常具有较高的进化程度,相反左下方的居群往往进化程度较低,因而我们认为34居群进化程度较低,37居群进化程度相对较高(图3,A)。同样,基于沙鞭居群染色体平均臂比和核型不对称系数的研究,我们发现沙鞭居群总体沿着二维图右上角方向进化,其中37居群的染色体平均臂比和核型不对称系数最大,表明进化程度较高;然而34居群的染色体平均臂比和核型不对称系数最小,说明进化程度较低(图3,B),这与上述研究结果一致。因此,本研究认为沙鞭34居群起源较早,进化程度较低,而37居群起源较晚,进化程度较高(图3)。
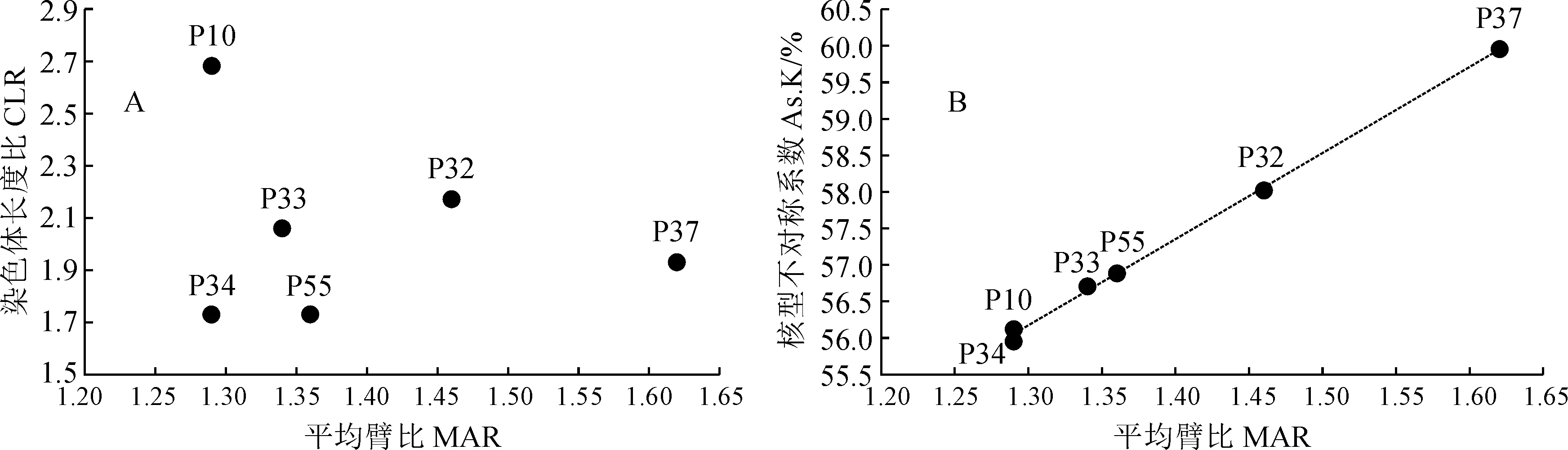
图3 沙鞭不同居群染色体核型进化趋势Fig.3 Evolution trend of chromosome karyotypes in different populations of P. villosa
续表2 Continued Table 2
续表2 Continued Table 2
续表2 Continued Table 2
2.5 沙鞭不同居群的染色体核型聚类分析
对沙鞭不同居群进行聚类分析,结果(图4)表明当遗传距离为1.0时,沙鞭6个居群聚为3类,其中32和33居群聚为一类, 10、34和55居群聚为一类,而37居群单独聚为一类;当遗传距离为9.0时,沙鞭6个居群聚为2类, 37居群仍单独聚为一类,而剩余所有居群聚为一类。研究表明在亲缘关系上,37居群与其他居群的相似度较小。
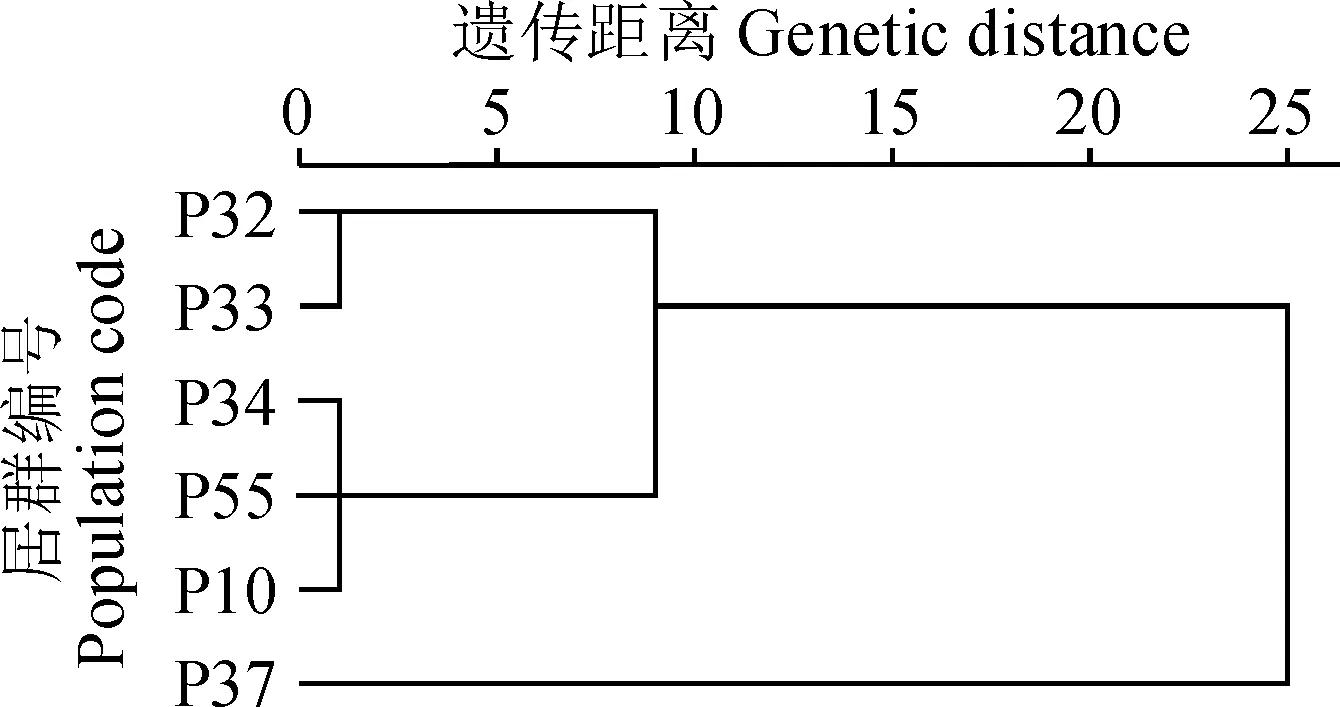
图4 沙鞭不同居群染色体核型系统聚类图Fig.4 The clustering analysis of chromosome karyotypes in different populations of P. villosa
3 讨 论
染色体作为生物细胞核中的重要组成成分,其数目、形态、结构和核型通常较为稳定,可作为物种分类和系统进化关系研究的重要细胞学依据[26-27]。本研究表明,沙鞭不同居群的染色体均为二倍体,染色体长度介于1 ~ 4.21 μm,隶属于小型染色体,这与李伟伟等[7]的研究结果一致;沙鞭所有参试居群的染色体数目均为46条,这与李伟伟等[7]先前报道的沙鞭染色体数目为40条显然不同,我们认为这可能是由于压片过程中染色体重叠所导致。同时,通过对沙鞭不同居群染色体参数的统计分析,我们还发现沙鞭不同居群之间的染色体形态存在差异,染色体长度比和平均臂比等不同。张建波等[28]对川西北高原12个垂穗披碱草(Elymusnutans)居群的染色体核型研究,认为同种植物染色体形态随着生境不同而发生变化,即存在染色体形态多样性。染色体形态是研究属下或种下物种或居群亲缘关系和演化趋势的主要核型特征[29],本研究参试沙鞭居群具有相同的染色体数目,但形态不同,可以推测它们属于同一物种的不同生态居群。同种植物在长期的自然选择和人工选择的作用下,为适应当地的自然生态环境有可能逐步演化成不同的生态居群[30]。因此,我们认为沙鞭不同居群染色体形态的差异也是由生长环境不同导致的,这进一步验证了先前诸多研究的合理性[18,28]。
研究表明,植物染色体核型的进化趋势由对称向不对称方向进化,因此系统进化上比较古老或原始的植物往往具有较对称的染色体核型,而不对称的染色体核型通常存在于较进化的植物类群中,即核型的对称性越高,其进化程度越原始[29,31]。通过对沙鞭6个居群染色体核型不对称系数和核型类型的研究,我们发现沙鞭居群核型不对称系数介于50% ~ 60%,核型类型为1A、2A、1B或2B型,属于对称性较好和较原始的核型类型,大小依次为37 > 32 > 55 > 33 > 10 > 34居群,表明它们在系统进化中的地位不同,同时这与先前报道的许多沙鞭近缘种如芨芨草、大针茅(Stipagrandis)等[32]的核型类型较为一致。此外,基于沙鞭居群核型进化趋势研究结果,我们同样发现沙鞭居群37的进化程度相对较高, 34居群的进化程度相对较低,表明37居群在沙鞭所有参试居群中具有最强的抗旱性[32],因此沙鞭37居群在今后沙鞭遗传育种、优良种质资源筛选以及品种推广应用等方面具有重要利用价值。
遗传物质的相似性不仅可以决定物种间的亲缘关系,而且可以推测物种在进化过程中彼此分化的历程。遗传物质在细胞水平上的表征就是核型。将核型数据进行数理统计并聚类分析,以此估算遗传距离,能够多向的、立体的、多维的考察物种间的相似性,从而判断物种间的亲缘关系[30]。此外,植物染色体核型聚类分析还能够直观反映不同居群间的遗传分化和亲缘关系[33]。本研究结果显示,当遗传距离为9时,沙鞭不同产地的6个居群聚为两大类, 其中一大类有10、32、33、34和37居群构成,表明这5个居群间具有相对较近的亲缘关系,彼此间相似度和进化程度较为类似;另一大类仅有 37居群组成,其与其他供试居群间亲缘关系较远。总之,本研究从居群水平上首次对沙鞭染色体核型进行了较为系统的报道,旨在为今后沙鞭的系统进化、优良种质资源筛选和新品种选育等研究提供细胞学依据。
致谢:感谢中国科学院西北高原生物研究所刘博师兄、青海师范大学梁瑞芳和吕婷师姐在沙鞭染色体压片,以及青海师范大学刘峰师兄在染色体核型模式图制作上给予的指导和帮助。
