GSNOR的原核表达及亚硝基化修饰对其活性的影响
2021-11-07张林林
张林林, 韩 毅
(合肥工业大学 食品与生物工程学院,安徽 合肥 230601)
0 引 言
S-亚硝基谷胱甘肽还原酶(GSNOR)的生物学研究起源于动物和大肠杆菌,从细菌到人类所有物种中都存在,且保守性高。其最初被认定为是一个依赖谷胱甘肽的甲醛脱氢酶,后来发现其催化反应过程包含 S-亚硝基谷胱甘肽羟基的氧化、谷胱甘肽与甲醛成形成 S-甲酰基谷胱甘肽[1]。因此,重新命名为 S-亚硝基谷胱甘肽还原酶,其在植物生长发育过程中扮演着重要的角色。研究已发现,在番茄中,抑制GSNOR的表达提高了内源的NO和S-亚硝基化水平,导致了发芽率提高、根和下胚轴生长被抑制、根毛数量减少,顶端优势被破坏,侧枝生长被促进、光合作用降低、果实结实率和产量低等不良影响[2];在拟南芥中发现GSNOR基因的突变出现了生长缺陷、植物疾病反应受损、热敏感性等问题[3-4]。因此,研究植物体内的GSNOR活性调节机制是十分必要的。
NO作为一种信号分子来调节植物中的许多细胞功能和信号传导,其在植物生长发育、代谢和胁迫响应等的调节功能已被广泛研究。在动物中,NO也以信号分子的形式介导神经调节、细胞凋亡等途径[5-6]。蛋白质的翻译后修饰是NO依赖信号通路的重要特征[7],NO可以通过蛋白翻译后修饰来影响酶或蛋白质的活性和功能。例如蛋白质的不可逆硝化和可逆亚硝基化等, NO也可以以离子形式结合到过渡金属上,形成金属-亚硝酰基络合物,称为金属亚硝化作用。其中作为亚硝基基团(—NO)与半胱氨酸(Cys)硫醇的共价连接的蛋白质S-亚硝化是NO生物学活性的关键机制[8-10]。S-亚硝基谷胱甘肽(GSNO)是NO的主要储存库和转运形式,比NO自由基稳定,可将其NO部分转移到蛋白质上,其在植物体内的水平由其合成或降解2个途径决定。NO及其衍生物活性氮在有氧条件下均能与细胞内的抗氧化分子谷胱甘肽反应形成亚硝基谷胱甘肽(GSNO)。非酶促途径和酶促途径是GSNO的2种降解途径[11],非酶途径为紫外线、高温、碱等的破坏,而酶促途径的关键酶为GSNOR,它不可逆降解GSNO,这一作用对于维持亚硝基化的稳态十分重要。
研究发现,拟南芥GSNOR包含9个高度保守的非锌配位半胱氨酸残基,可发生S-亚硝化、谷胱甘肽化或可逆氧化等翻译后修饰,其中Cys10、Cys271、Cys370 共3个位点被认为在GSNOR活性的调节中具有保守作用[12]。本文运用原核表达的方法将GSNOR体外表达并分离出,探索NO能否亚硝基化GSNOR,以及能否通过这种修饰影响GSNOR的活性。
1 材料与方法
1.1 材料
1.1.1 实验材料
拟南芥(哥伦比亚)、PET28a(+)质粒、大肠杆菌DH5α克隆菌株、大肠杆菌BL21表达菌株。
1.1.2 实验试剂
反转录酶、高保真DNA聚合酶、限制性内切酶、Taq酶、T4DNA连接酶、镍柱、咪唑、质粒提取试剂盒、普通DNA产物纯化试剂盒、琼脂粉、酵母提取物、牛肉膏。
1.2 仪器
高温高压灭菌锅、恒温培养箱、超净工作台、真空泵等恒温培养震荡器、紫外分光光度计、冷柜等。
1.3 方法
1.3.1 目的基因克隆
从tair网站中搜索目的基因的cDNA序列(GSONR的登录号为AT5G43940),根据cDNA序列设计出引物GSNOR-F和GSNOR-R。
提取野生型拟南芥总RNA,将mRNA反转得到cDNA,以cDNA为模板扩增GSNOR基因的全编码序列(coding sequence,CDS)。聚合酶链式反应(polymerase chain reaction,PCR)的反应条件为98 ℃、3 min;98 ℃、10 s;54 ℃、15 s;72 ℃、80 s;72 ℃、10 min。PCR产物和PET28a(+)载体的连接产物转化大肠杆菌,确定阳性菌落后,提取质粒并测序。测序结果符合已知基因序列,将质粒转化大肠杆菌表达感受态BL21,确认阳性菌落,保存甘油菌备用。
1.3.2 蛋白的诱导
将转化大肠杆菌 BL21的阳性菌接种在4个装有250 mL LB+250 μL kana的培养液中,在220 r/min、37 ℃条件下摇至菌的OD600为0.6,在每个锥形瓶中加1 mol/L的IPTG 50 μL(终浓度为0.2 mmol/L),22 ℃条件下诱导16~24 h。SDS-PAGE电泳确认是否诱导成功。
1.3.3 蛋白的纯化
收集菌体沉淀并用溶液Ⅰ(20 mmol/L Tris-HCl、300 mmol/L NaCl,pH值为7.6)重悬,细胞裂解30 min,离心收集上清并抽滤,通过镍柱纯化蛋白,所有上柱的液体都需抽滤。用溶液Ⅰ、30 mL溶液Ⅱ(20 mmol/L Tris-HCl、300 mmol/L NaCl 、50 mmol/L咪唑, pH值为7.6)洗脱杂蛋白,用溶液Ⅲ(20 mmol/L Tris-HCl、300 mmol/L NaCl 、300 mmol/L咪唑,pH值为7.6)洗脱目的蛋白。利用透析法将咪唑除去,将蛋白保存在-80 ℃备用。
1.3.4 蛋白质量浓度和酶活的检测
以牛血清蛋白为标准蛋白制作标准曲线,将目的蛋白稀释合适的倍数测A595,计算蛋白质量浓度。底物GSNO在利用NADH的条件下能被GSNOR降解,测定NADH在A340的吸光值可计算出GSNOR的酶活[13]。根据实际的蛋白质量浓度来决定GSNOR用量。测定buffer(20 mmol/L Tris、0.5 mmol/L EDTA,pH值为8.0)、GSNOR蛋白、20 mmol/L GSNO的总体积为l mL,底物GSNO终浓度为400 μmol/L,检测2 min。
1.3.5 亚硝基化水平检测
本文利用生物素转换技术(biotin switch technique,BST)[14]来检测GNSOR能否被NO亚硝基化。先通过MMTS封闭巯基(—SH),然后用抗坏血酸将SNO还原为—SH,再用生物素Biotin标记—SH。处理的样品经SDS-PAGE电泳后,蛋白转到聚偏二氟乙烯膜上与生物素亲和抗体Biotin-HRP孵育。显影后可观察到GSNOR蛋白亚硝基化水平。
1.3.6 DEA/NO和DTT处理GSNOR蛋白
1 mmol/L的NO供体2-(N,N-二乙基氨基)-二氮烯-2-氧二乙铵盐(DEA/NO)和GSNOR蛋白孵育30 min后,一组加20 mmol/L 二硫苏糖醇(DTT)孵育30 min,另一组加缓冲液放置30 min,分别测定未用DEA/NO处理、DEA/NO处理但未加DTT还原及DEA/NO处理后加DTT还原这3组处理后的GSNOR蛋白的酶活。
2 结果与分析
2.1 GSNOR的PCR扩增结果
本文以拟南芥野生型的cDNA为模板,以GSNOR-F和GSNOR-R为引物进行扩增,核酸琼脂糖电泳检测结果如图1所示,由图1可知,扩增产物有一条特异性带,与拟南芥GSNOR基因的大小相符(GSNOR大小为1 176 bp)。
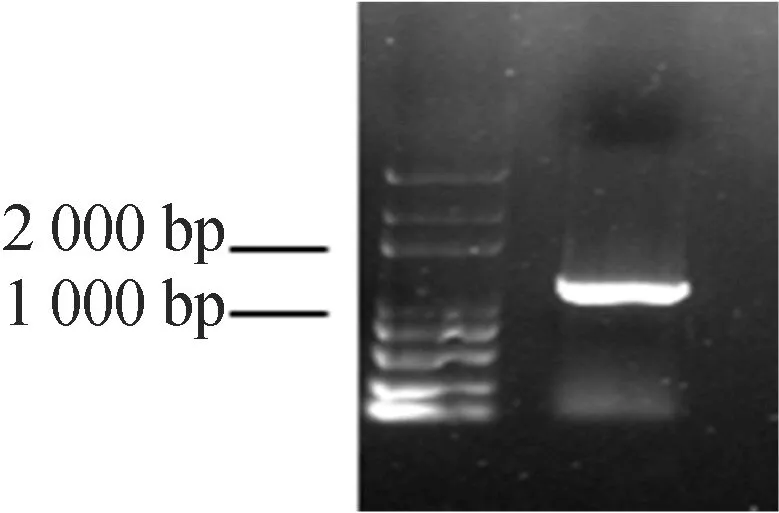
图1 PCR扩增GSNOR基因片段琼脂糖电泳图
2.2 PET28a(+)-GSNOR原核表达载体的鉴定
提取阳性菌落质粒进行测序,测序结果显示质粒中连接的目的基因与GSNOR基因片段完全相符,未发生任何突变,将送去测序的菌落进行扩摇,提取质粒并转入表达菌株BL21,挑取单克隆进行菌落鉴定。以GSNOR-F和GSNOR-R为引物,确认GSNOR与载体是否成功连接。菌落PET28a(+)-GSNOR/BL21的PCR电泳图如图2所示,图2中,条带在1 000 bp稍多的位置,与实际大小1 176 bp相符,因此可以选取含目的基因的单克隆作为诱导菌种。
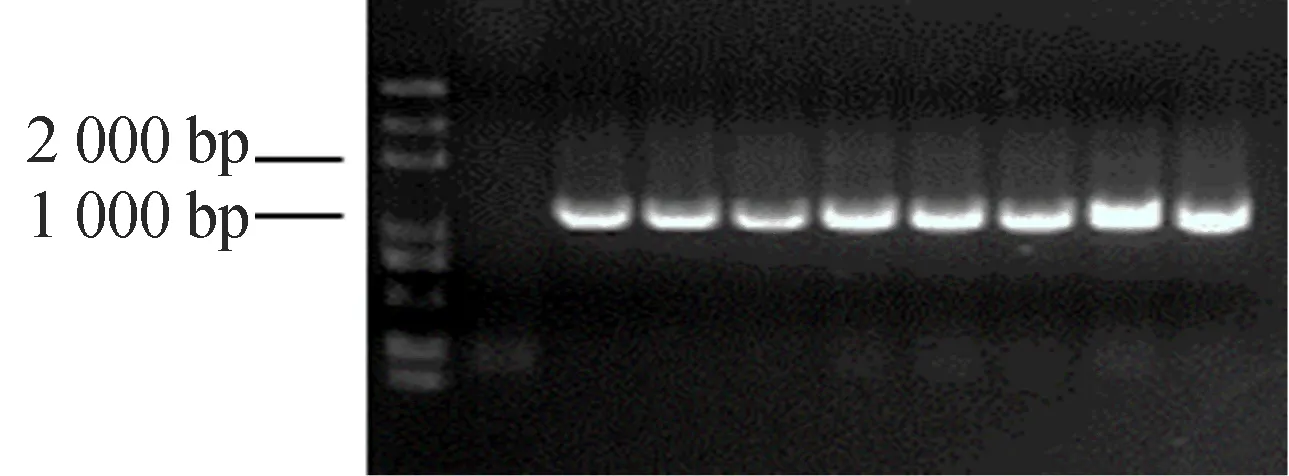
图2 菌落PET28a(+)-GSNOR/BL21 PCR电泳图
2.3 GSNOR蛋白的诱导结果
在22 ℃条件下,0.2 mmol/L IPTG诱导12~16 h,SDS-PAGE电泳检测诱导产物如图3所示,图3中,M代表Marker;1泳道代表未诱导时大肠杆菌中的蛋白分布;2、3泳道代表诱导后的蛋白分布。由图3可知,在接近42.7 kDa(GSNOR为42 kDa,6个组氨酸为0.9 kDa)处,诱导后的2、3泳道与对照1泳道相比明显增多,表明GSNOR蛋白被成功诱导。
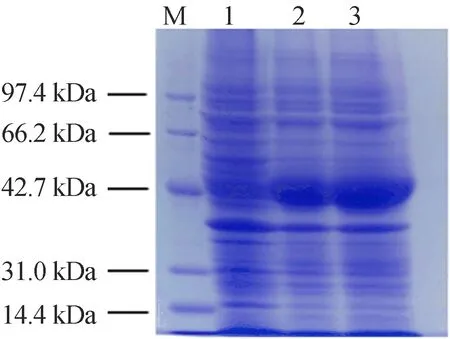
图3 GSNOR蛋白诱导表达SDS-PAGE电泳图
2.4 GSNOR蛋白的纯化结果
诱导的粗蛋白利用镍柱纯化,在蛋白上样后,带有His标签的融合蛋白特异性结合到柱子里,其他的杂蛋白随之流出。镍与咪唑发生竞争性结合,目的蛋白则被洗脱下来,即GSONR与杂蛋白成功分离,SDS-PAGE电泳检测不同时段的洗脱液,电泳图如图4所示。
图4中,M代表Marker;1、2、3泳道代表杂蛋白的洗脱结果;4、5、6泳道代表目的蛋白的洗脱结果,4泳道的洗脱液含大量的目的蛋白,利用透析袋将该洗脱液的咪唑脱去后,将目的蛋白置于-80 ℃保存。
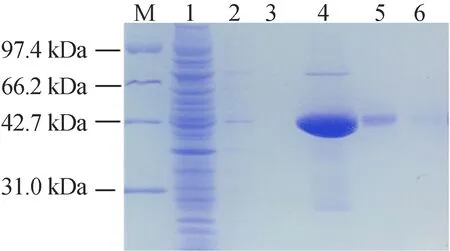
图4 GSNOR蛋白纯化SDS-PAGE电泳图
2.5 GSNOR蛋白的质量浓度及酶活检测结果
以牛血清蛋白为标准蛋白制作标准曲线,据此标准曲线算出蛋白质量浓度为0.77 μg/μL。以GSNO为底物,NADH为还原力测得酶活约为140 U/mg。蛋白质量浓度、纯度及活力足以支持下一步实验。
2.6 亚硝基化水的检测结果
为确定NO对GSNOR的修饰是否是亚硝基化修饰,本文利用BST技术检测NO处理后的GSNOR的亚硝基化程度和DTT还原NO处理的GSNOR蛋白的亚硝基化程度,结果如图5所示。由图5可知,未用NO处理时GSNOR蛋白几乎无亚硝基化,而NO处理后未用DTT还原的GSNOR蛋白发生了明显的亚硝基化,加DTT还原后,亚硝基化程度明显削弱。表明GSNOR在NO处理后经过了S-亚硝基化。

图5 GSNOR的亚硝基化水平分析
2.7 GSNOR蛋白处理后的酶活检测结果
DEA/NO和DTT处理 GSNOR后的酶活分析如图6所示,由图6可知,1 mmol/L NO供体DEA/NO处理GSNOR后会导致其酶活降低,且还原剂DTT会使GSNOR酶活恢复。说明NO使得GSNOR发生了亚硝基化修饰,且这种修饰导致了GSNOR活性下降,还原剂DTT可以还原这种修饰,使酶活恢复。

图6 DEA/NO和DTT处理 GSNOR后的酶活分析
3 讨论与展望
蛋白质是生命的物质基础,是构成细胞的基本有机物,研究蛋白质的功能与结构是重中之重,获得大量纯化的蛋白质是研究的基础。许多蛋白翻译后修饰发现是通过体外表达目的蛋白,然后与猜测的物质直接反应从而确定两者是否有关联,所得到的结果可为体内的研究提供思路。目前有四大蛋白表达系统,分别为哺乳动物蛋白表达、原核表达系统、酵母表达系统和昆虫表达系统。原核表达系统中的大肠杆菌表达系统是最常见的,这是由于大肠杆菌遗传背景清楚、表达水平高、成本低、操作简单。原核蛋白表达是将外源目的基因,通过构建表达载体并导入特定原核生物或细胞内表达(如大肠杆菌)。表达载体带有特定标签蛋白,例如His标签蛋白,由6个寡聚的组氨酸组成,标签是目的蛋白纯化的关键。文中表达出的GSNOR蛋白酶活约为140 U/mg,质量浓度约为0.77 μg/μL。文献[15]的GSNOR蛋白的酶活仅有100 U/mg[15],此方法可高效表达出GSNOR蛋白,说明原核表达载体及相关条件适合GSNOR蛋白的表达。蛋白被修饰后,常用质谱法鉴定修饰位点,样品所需体积小,同时对蛋白含量也有较高的要求,高效表达出高质量浓度的蛋白可解决这一问题。
NO的主要作用是对目标蛋白的半胱氨酸残基进行亚硝基化的修饰。大多数蛋白质都含有半胱氨酸,但这种氨基酸残基与NO的亲和力可能有很大的不同。在文中,BST结果显示,DEA/NO处理后,GSNOR蛋白发生了亚硝基化修饰,DTT显著削弱亚硝基化水平,且GSNOR酶活被NO供体DEA/NO可逆抑制。此外,GSNOR的酶活也可被GSNO抑制,实验过程中发现还原剂DTT可降解GSNO,因此选用了DEA/NO作为NO供体。两者皆可修饰GSNOR,使得GSNOR酶活下降,说明GSNOR的某些半胱氨酸残基被NO修饰且影响了GSNOR的活性。GSNOR的这种修饰可能代表一种信号,对细胞内的GSNO水平起着调控作用。GSNOR可降解GSNO,而NO及其衍生物又能修饰GSNOR,由此推测,NO可能通过修饰GSNOR的半胱氨酸影响GSNOR的活性,进而影响了GSNO的水平。
4 结 论
本文建立了体外高效表达GSNOR的方法,获得了质量浓度约0.77 μg/μL的GSNOR蛋白,并探索亚硝基化修饰对GSNOR酶活的影响,为NO的信号转导机制及GSNOR的功能和结构的研究提供了思路和基础。
