转基因作物对土壤微生物群落影响的研究进展
2021-11-06王婷杨阳李金萍杜坤
王婷 杨阳 李金萍 杜坤
(1. 扬州大学生物科学与技术学院,扬州 225009;2. 扬州大学生命科学基础实验教学中心,扬州 225009)
转基因(genetically modified,GM)作物是一类利用基因工程技术将外源的有益基因导入到作物基因组中并稳定遗传,以获得更加优良的农艺性状和经济价值的作物。自1996年第一例转基因作物商业化应用以来,各种转基因作物不断问世,且种植面积也不断扩大。目前,全球共有27种转基因作物的387个转化体被批准可用作粮食或饲料。全转基因作物的总种植面积从1996年的170万公顷增加到2018年的191.7万公顷(ISAAA,2019)。根际导入外源基因的类别,可以把转基因作物分为4类:(1)耐除草剂类,(2)抗虫类,(3)抗病类,(4)其他(品质相关、抗逆等)。
由于外源基因来源的多样化和植物发育的不可预测性,转基因作物商业化种植之前都必须通过系统、深入的环境安全性评价[1]。其中,土壤作为生态系统中物质循环和能量转化的重要场所,对外界环境的变化比较敏感,已被公认为是生态系统变化的预警指标之一[2]。本文综述近年来国内外学者关于转基因作物对土壤微生物群落影响方面的研究进展,并对转基因作物土壤安全评价中还应注意的问题进行了探讨和展望。
1 土壤微生物群落的研究方法
随着现代生物技术的发展,土壤微生物群落的主要研究方法有微生物平板培养法、Biolog微孔板法、生物标记物法和高通量测序技术4类。
土壤中的微生物可以通过经典的微生物平板培养方法直接检测出,但土壤微生物种类繁多,可培养的微生物只占所有土壤微生物的10%左右[3-4],这种方法只能反映出土壤中一部分微生物的变化情况。因此,微生物平板培养法现在大多被用来筛选一些具有某些特定功能的可培养微生物。
Biolog微孔板法是根据微生物对特定碳源利用方式的差异,来测算微生物群落代谢的功能多样性,即细菌群落代谢指纹[5]。Biolog微孔板有多种类型,目前以Biolog-ECO 1506应用最为广泛。它含有3组纵向排列的平行重复,每组含有31种碳源(分别是糖类、氨基酸类、酯类、醇类、胺类和酸类六大类)和1个空白对照。由于不同微生物对碳源的利用方式有一定差异,因此就可用于比较不同微生物群落之间的代谢差异。这种方法已被广泛的应用于评价转基因作物(如玉米、水稻、小麦和大豆)对根际土壤微生物群落的影响[6-9]。Biolog微孔板法虽然具有灵敏度高、培养周期短、检测简便等优点,但也存在一些局限性,如人工培养条件对实验结果的影响等[10]。
土壤微生物主要由细菌、真菌和放线菌等组成,不同微生物细胞膜中特定化合物(如醌和脂肪酸)的类型和含量存在一定差异,将这些化合物作为生物标记物,就可实现对微生物群落结构的定量分析,其中应用最多的就是磷脂脂肪酸(PLFA)图谱法[11]。常用的细菌特征PLFA有i15:0、a15:0、15:0、i16:0、16:1ω9、16:1ω7t、i17:0、a17:0、17:0、18:1ω7、cy19:0,真 菌 特 征PLFA有18:1ω9、18:2ω6、18:3ω3;革兰氏阳性菌的特征PLFA有16:0(10Me)、17:0(10Me)、18:0(10Me)、i15:0、a15:0、i16:0、i17:0、a17:0,革兰氏阴性菌的特征PLFA有16:1ω5、16:1ω7t、16:1ω9、cy17:0、18:1ω5、18:1ω7和cy19:0[12]。由于PLFA方法直接以土壤微生物群落中的脂肪酸为研究对象,解决了传统微生物培养方法中缺失不可培养微生物信息的问题[13]。但是,PLFA生物标记方法只适用于活的微生物,且不同种类的微生物可能会产生重叠的图谱,故此方法一般被用作为微生物群落研究的一种必要补充。
分子检测技术对微生物群落结构的变化十分敏感,可以检测细菌组成的微小差异,其在探索微生物群落多样性,尤其是鉴定微生物新物种方面发挥了一定的优势。常用的方法有荧光原位杂交法(FISH)、DNA微阵列法、单链构象多态性(SSCP)法、限制性片段长度多态性(RFLP)法、扩增核糖体DNA限制性分析(ARDRA)法、扩增片段长度多态性(AFLP)法、变性/温度梯度凝胶电泳(DGGE/TGGE)法等。其中,DGGE技术是被运用于土壤微生物群落研究中最多的方法。这些分子生物学方法具有灵敏度高、不用培养微生物等优点,可以更好的反应土壤微生物的真实状况。但是缺点也很明显,如DGGE中能够检测到的微生物群落通常是具有显著优势度的菌落,同一条带可能代表DNA分子遗传图谱中的多种微生物物种[14]。
随着分子生物学研究的快速发展,尤其是下一代测序(NGS)技术的广泛应用,极大的提高了土壤微生物学的研究广度和深度[15-16]。高通量测序技术主要包括 454 焦磷酸、ABI SOLiD、Illumina Hiseq 2000 及Illumina Miseq、Hecos Heliscope和PacBio Sequel测序平台。近年来,这些高通量测序已逐渐代替了DGGE技术,尤其是以单分子和长读长为主要特征的三代测序技术,逐渐成为微生物群落研究领域的主要分析方法[17]。如PacBio 16S扩增子测序技术,首先对细菌16S rRNA基因的扩增测序结果(raw reads)进行校正、过滤和去除嵌合体序列,得到有效数据(clean reads),然后将clean reads聚类为OTUs,并对其进行物种注释,获得分类学信息。再使用Qiime等软件计算Observed-OTUs,Chao1,Shannon,Simpson等Alpha多样性指数,计算Unifrac距离,构建PCoA和UPGMA聚类树图,并进行组间差异分析。最后运用Anosim、MRPP、ADONIS、t-test等统计分析方法对组间物种组成和群落结构进行差异显著性检验。
宏基因组测序是以特定生境中的整个微生物群落作为研究对象,直接提取环境样本DNA,通过采用新一代测序技术,获得环境微生物基因信息的总和[18]。与高通量测序不同的是,Clean Reads需经过Metagenome 组装后,进行常用功能数据库、抗性基因数据库的注释和一系列高级信息分析(如CCA/RDA分析,拷贝数变异分析等)。同时,还可结合环境因子、病理指标或特殊表型进行深入关联分析。宏基因组测序技术避免了实验中由环境引起的微生物变化的差异,更真实地反应样本中微生物组成和互作情况,可在分子水平对其代谢通路、基因功能进行更加深入的研究[19]。宏转录组测序是指从整体水平上研究某一特定环境,特定时期群体生命所有基因组的转录情况以及转录调控规律。相较于宏基因组测序,它以生态环境中的全部RNA为研究对象,从转录水平上研究复杂微生物群落的变化,可以更好的挖掘潜在的新基因[20]。
2 转基因作物对土壤微生物影响的研究进展
转基因作物在给我们带来效益的同时,外源基因的导入是否会引起土壤生态风险,这是我们急需回答的问题。为此,国内外研究学者做了大量的研究工作,但目前还没有明确的结论表明转基因作物是否一定会对土壤微生物生态系统产生持续、不利的影响。
2.1 抗虫类转基因作物
已转入作物中的抗虫类基因主要包括:苏云金杆菌(Bacillus thuringiensis)毒蛋白基因、蛋白酶抑制剂基因、淀粉酶抑制剂基因、外源凝集素基因、核糖体失活蛋白基因和豌豆脂肪氧化酶基因等[21]。目前,在评估抗虫类转基因作物对非靶标土壤微生物影响的研究中,最多的是Bt基因和CpT1基因(表1)。
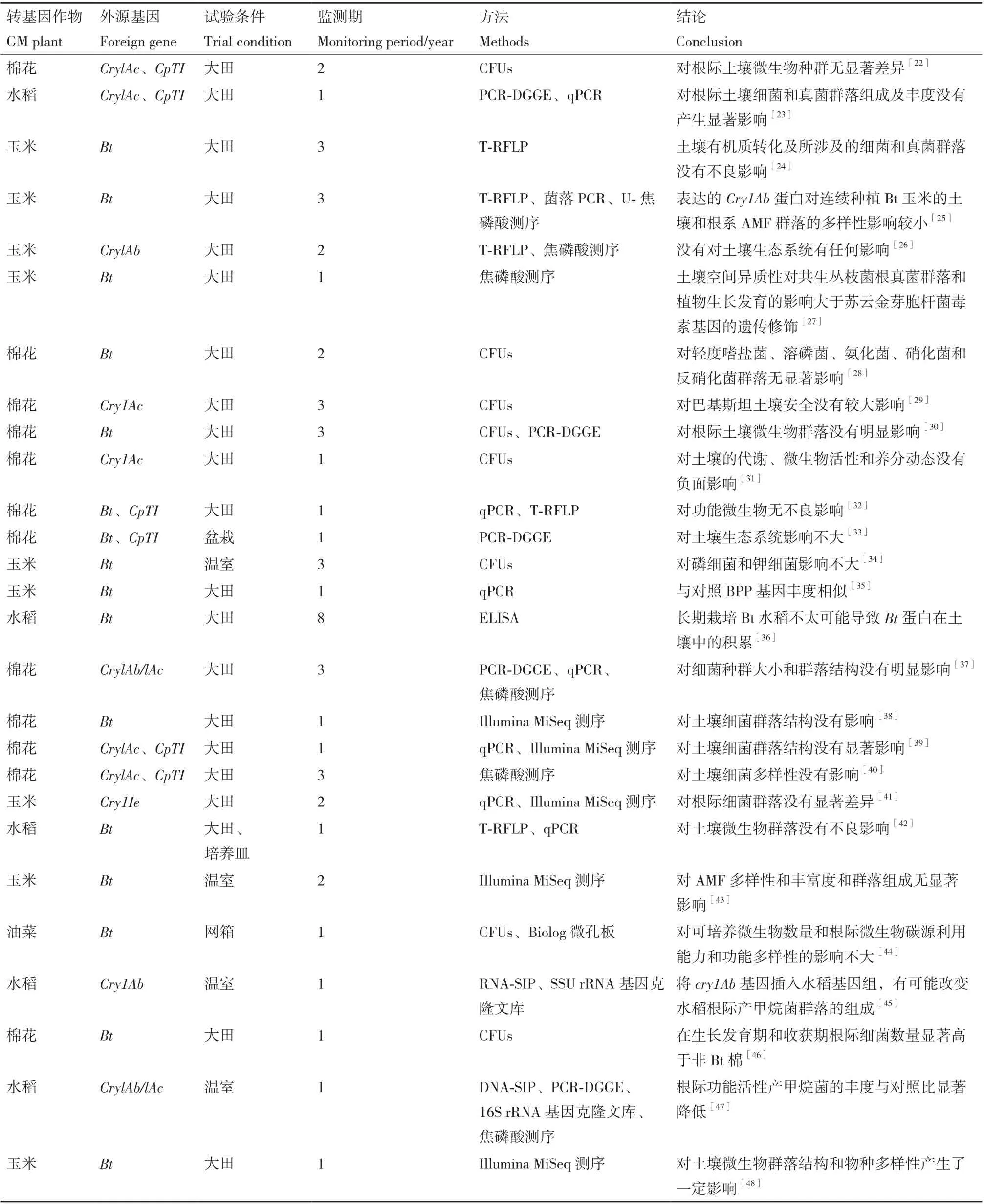
表1 抗虫类转基因作物对土壤微生物群落的影响Table 1 Impact of insect-resistant GM crops on soil microbial communities
上述研究中,大多数研究发现转基因抗虫作物对根际土壤微生物群落无显著影响,或远小于作物不同生育期和种植年份。如在转基因棉花中,研究学者已经运用多种研究方法(包括CFUs、qPCR、T-RFLP、PCR-DGGE和16S rRNA基因高通量测序)分析了转CrylAc+CpTI 基因棉花对根际土壤微生物的影响。结果表明,转基因棉花根际土壤中的细菌和真菌的优势菌群种类、物种相对丰度、群落多样性等指标与对照相比差异并不显著[28,30,32-33,40]。Liang等[41]的高通量测序结果显示,转Cry1Ie 基因玉米和非转基因玉米的根际土壤细菌丰富度和多样性指数以及群落结构均无显著差异,而主成分分析却显示苗期、花期、成熟期之间或不同试验年份之间的却存在显著差异。
另一方面,Zhu等[45]和Han等[47]利用同位素探测技术结合基因克隆文库分析显示,虽然Bt水稻、亲本Ck和远源亲本Dp三种水稻的根际土壤中占优势的微生物都是产甲烷古菌群落,但其中Bt水稻根际活跃的产甲烷古菌群落以甲烷八叠球菌科、甲烷鬃菌科和甲烷微菌科为主,而Ck和Dp水稻根际只有两个主要产甲烷菌群(甲烷鬃菌科和甲烷微菌科)。这些结果表明,将Cry1Ab/1Ac基因插入水稻基因组中,有可能通过改变根系产甲烷底物的分泌(如氨基酸、糖、可溶性碳水化合物、柠檬酸、乙酸和总有机酸等)、根系衰老过程或光合作用等其他途径,改变了水稻根际产甲烷群落的结构组成。Van wyk等[48]通过Illumina MiSeq 测序技术研究了成熟期Bt玉米和非Bt玉米的根际土壤细菌物种多样性和群落结构。结果表明,两者细菌alpha多样性和群落组成存在显著差异,变形菌门和酸杆菌门是Bt玉米根际土壤的优势菌,而非Bt玉米根际土壤的优势菌则是放线菌门。通过进一步研究,发现Bt玉米与对照根际土壤的有机碳、硝酸盐含量以及土壤酸性磷酸酶和β-葡萄糖苷酶活性也存在显著差异。这些土壤理化性质和相关酶活性的差异可能是导致细菌群落结构不同的主要原因。
2.2 抗病类转基因作物
在评估抗病类转基因作物对土壤微生物群落影响的研究中,研究较多的是转抗真菌和抗植物病毒基因的作物品系(表2)。
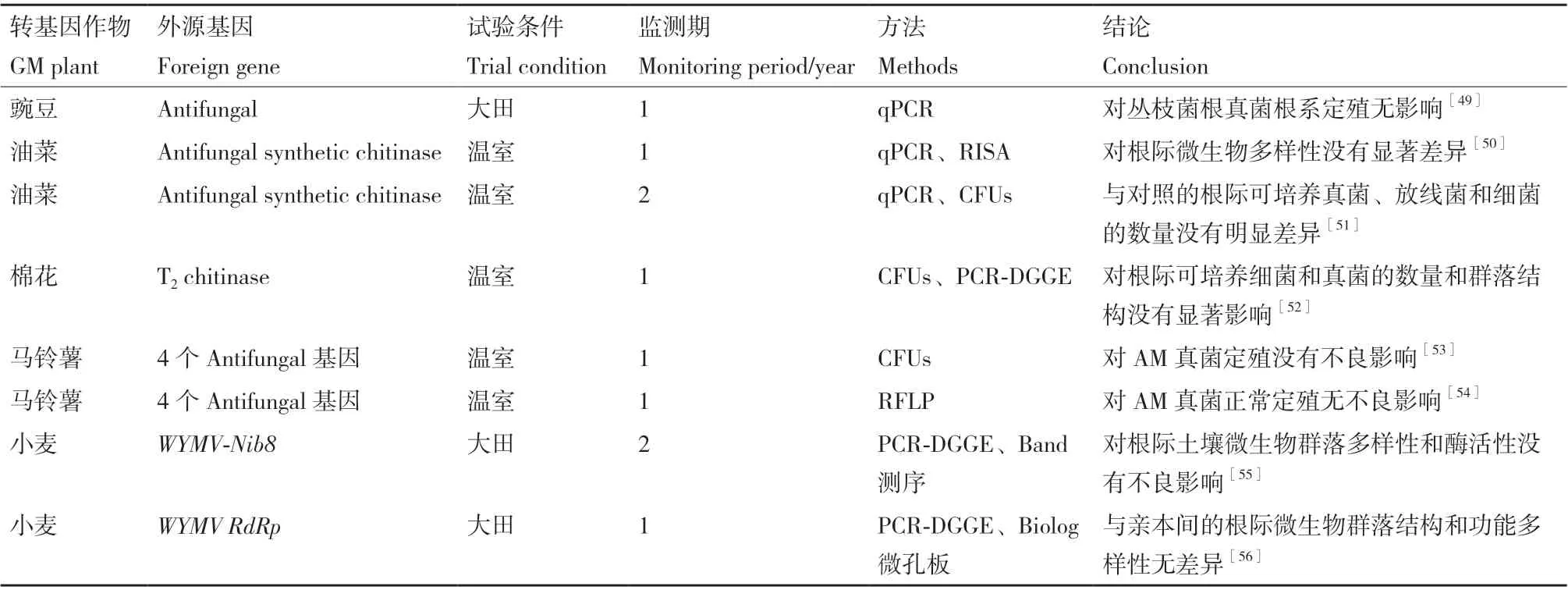
表2 抗病类转基因作物对土壤微生物群落的影响Table 2 Influence of disease-resistant GM crops on soil microbial communities
表达具有抗菌活性的外源蛋白(如几丁质酶、葡聚糖酶和溶菌酶),是一种提高作物抗病能力的主要分子育种手段。几丁质酶可以通过分解真菌细胞壁来实现抗真菌的特性,但据现有的文献报道,表达几丁质酶的转基因作物对非目标和潜在有益的微生物群落并没有产生不良影响。Khan等[50-51]的研究发现,转几丁质酶基因甘蓝型油菜对根际土壤细菌、真菌和放线菌数量、多样性和群落结构均无显著影响,两者间5种土壤酶活性(β-葡萄糖苷酶、脲酶、蔗糖酶、芳基硫酸酯酶和磷酸单酯酶)也无显著差异,但不同品种之间土壤的细菌和真菌群落结构差异显著。Shahmoradi等[52]通过盆栽实验,对转几丁质酶基因棉花T2代植株的根际和根区土壤微生物进行了CFUs和PCR-DGGE分析,结果显示,虽然根际土壤中的细菌群落会随生育期(幼苗、花蕾、开花和铃期)的变化而发生显著的改变,但同一生育期转几丁质酶棉花的根际细菌的群落结构和功能菌数量与对照差异不显著。此外,Bidondo等[53]和Stephan等[54]在盆栽条件下,利用RFLP技术监测了过表达抗真菌活性肽基因的转基因马铃薯对丛枝菌根真菌定殖的影响,结果表明转基因马铃薯丛枝菌根真菌的定殖和共生关系与对照相比没有显著的差异。以上结果表明,像几丁质酶这些具有广谱抗菌功能的蛋白可能并不会通过植物根系分泌至土壤中,或者说在植物体内表达的这些蛋白随着植物的衰老、凋亡并不会大量的释放到土壤环境中,因此没有显著改变土壤微生物的群落多样性和结构。
作物中导入一些植物病毒外壳的相关基因,如小麦黄花叶病毒(WYMV),已被证实可以显著提高对特定植物病毒的抗性。病毒外壳蛋白在转基因植物的所有部位均有表达,可能会对根际土壤微生物有潜在的非靶向效应。Wu等[55]分析了2个耕作年份转WYMV-Nib8基因小麦对土壤微生物群落结构的影响。结果显示,与受体扬麦相比,除第1年的灌浆期,其他时期(包括第2个试验年份)转基因小麦对土壤细菌的Shannon、Simpson和Evenness三个alpha多样性指数的影响不显著;除第1年苗期和第2年的成熟期外,转基因小麦与受体根际土壤真菌群落多样性差异也不显著。这些结果说明,WYMVNib8基因的导入并未对土壤根际微生物的群落产生较长期的影响。Zhang等[56]采用PCR-DGGE和Biolog微生态板法研究了转WYMV RdRp基因小麦和亲本小麦的根际微生物群落,发现不同生育期间(苗期、茎伸长期、花期和成熟期)物种相对丰度和微生物群落碳源利用能力存在明显不同,而不同基因型间的根际微生物群落结构和功能多样性指数没有显著差异,根际细菌多样性主要与土壤含水量、有效磷、pH等环境因子高度相关。
2.3 抗除草剂类转基因作物
转抗除草剂基因作物中运用最多的是抗草甘膦基因和抗草铵膦基因。其中,抗草甘膦基因有7种,分 别 是cp4 epsps、gat4601、goxv247、epsps、epsps grg23ace5、mepsps、2mepsps,抗草铵膦基因主要有bar和pat两种[57-58](表3)。
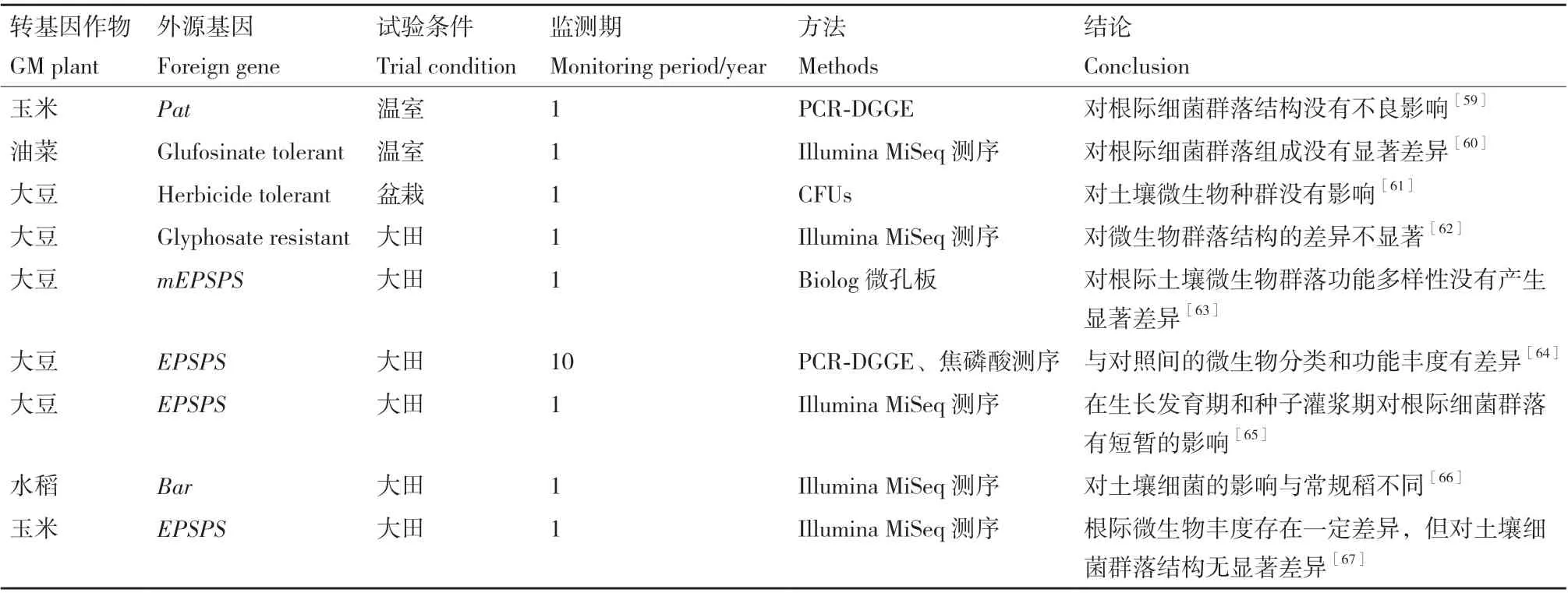
表3 抗除草剂类转基因作物对土壤微生物群落的影响Table 3 Effects of herbicide-resistant GM crops on soil microbial communities
和抗虫转基因作物的研究结果相似,评估抗除草剂类转基因作物对土壤微生物生态系统的影响研究中,也存在两种结果。一方面,大多数结果表明作物中转入抗除草剂基因对土壤微生物群落没有造成特异性的影响,而主要受季节和植物发育的影响较大。Vital-Lopez等[59]通过PCR-DGGE图谱分析,表明温室条件下转pat基因玉米对根际土壤细菌群落结构的影响不显著。Tang等[60]采用16S rRNA基因高通量测序技术,发现抗草铵膦油菜对根际细菌群落alpha多样性没有较大影响,相对丰度中较高的优势菌为变形菌门、拟杆菌门、酸杆菌门、芽单胞菌门和放线菌门,这些优势门在施用草铵膦的转基因油菜和非转基因受体油菜中都高度存在,但却在油菜的不同生育期(苗期、抽薹期、开花期和成熟期)存在显著的差异。而另一方面,也有一些研究表明,在较长时间的作用下,一些转抗除草剂基因作物会影响土壤中部分微生物的相对丰度,进而改变微生物群落的组成。Babujia等[64]对抗草甘膦转基因大豆进行了长达10年的田间监测,虽然抗草甘膦基因的导入并未对大豆的产量产生影响,但是通过宏基因组测序分析发现常规大豆根际土壤细菌群落中放线菌和酸杆菌丰富,而转基因大豆则变形菌、厚壁菌和绿藻相对丰度较高,与蛋白质代谢、细胞分裂周期相关的序列丰度也较高。He等[66]对成熟期细菌的16S rRNA基因进行了Illumina MiSeq测序发现,转bar基因水稻与常规水稻的土壤细菌多样性指数差异不显著,但是转基因粳稻和常规品种之间个别属的丰度(如绿丝菌属)存在显著差异,且转基因粳稻土壤中的厌氧菌含量显著高于常规水稻。
3 转基因作物土壤微生物生态风险评价中还应考虑的因素
虽然上述大量的研究结果显示作物土壤微生物生态系统主要和环境、气候、土壤类型、作物种类等密切相关,通过分子育种手段将外源有益基因导入到作物中,从短期来看并不会对土壤微生物的多样性指数和群落结构等产生较大的、持久的、不利的影响,但是确实也有一些证据表明,转基因事件可能会影响土壤中个别微生物种群的丰度,或者在某个特定时期影响微生物的多样性。因此,为了能够更加科学、准确、全面地评价转基因作物对土壤微生物的潜在风险,我们认为还应当考虑以下因素。
3.1 转基因产物在土壤中的持久性
转基因作物在生长发育时期,往往会通过根系向土壤中释放外源蛋白,其降解速率,或着说残留时间,也是影响土壤微生物群落的重要因素之一。Wang等[4]对Bt水稻土壤中Cry1Ab/1Ac蛋白的丰度和持久性等动态过程进行了为期3年的田间监测。结果显示,Bt水稻在生育期可通过根系向土壤和农田水体中持续释放Cry1Ab/1Ac蛋白,但是在土壤中可检测到的Bt蛋白不会持续存在超过2个月,并且其流动性不强,不会随着灌溉水进入相邻的土壤。Strain等[68]的研究却发现,Bt蛋白会通过地表径流和作物残留物被运输到附近的河道中,以致在非Bt玉米田中也检测到了Cry1Ab蛋白,且沉积物类型和温度对水中Cry1Ab蛋白的降解时间有一定的影响,最长可达2个月。
3.2 标记基因对土壤微生物的影响
标记基因主要用于阳性转化体的筛选,除了目的基因,这些标记基因也有可能会影响土壤微生物的群落,而这部分的研究报道较少。LeBlanc等[69]将CryIAb、uidA和nptII基因导入白云杉中,并对其根际微生物群落进行了研究。结果显示,转3种基因的与仅携带uidA和nptII标记基因的白云杉以及对照根际微生物物种相对丰度存在显著差异,说明uidA和nptII标记基因的导入也会对土壤根际微生物群落结构产生显著的影响。绿色荧光蛋白(GFP)也在植物遗传转化系统中有着广泛的应用。Lv等[70]发现,含GFP基因的烟草与对照野生型相比,在整个生育期(幼苗期、营养期、花期和衰老期)对土壤微生物种群没有产生显著影响。因此。标记基因对土壤微生物生态系统是否安全也存在争议,需要进一步的研究。
3.3 田间管理对土壤微生物的影响
轮作方法,肥料、杀虫剂和除草剂的使用等这些田间管理措施对土壤微生物也有着较大的影响。如种植转Bt基因作物会减少杀虫剂的使用,因此在这种情况下,还应同时评估农药用量的变化对土壤微生物产生的影响。再如,种植抗草甘膦作物的田间管理模式主要表现为免耕和除草剂的大量使用,这也会在一定程度上影响土壤的微生物群落[71-72]。Lupwayi等[7]用5年时间比较了抗草甘膦、抗草铵膦转基因玉米和常规品种在除草剂喷施、不同轮作方式等条件下的土壤细菌群落。在5个耕作年中,未发现Bt基因转化、杀虫剂施用或轮作对根际土壤微生物多样性产生影响,但却发现轮作2种Bt转基因玉米的非根际土壤微生物总量、功能多样性和土壤酶活性均高于1种Bt转基因玉米单作和常规玉米轮作。
一些研究学者认为长期种植转基因作物而导致的这些大田管理模式的变化会带来严重的生态和农艺问题,包括抗除草剂杂草的蔓延、土壤的腐蚀、土壤肥力和养分的丧失、物种和生物多样性的丧失等[73-74]。例如转基因大豆单作规模的扩大造成了农业生产的大规模集约化,土壤肥力下降和土壤腐蚀的加剧,也导致了部分土壤不能再持续利用。因此,我们在开展转基因作物土壤微生物生态风险评价工作中也应该考虑这些因素。
4 展望
土壤微生物在农业生产系统中发挥着极其重要的作用,外源基因的导入可能会影响植物的新陈代谢和根系分泌物的产生,从而改变土壤微生态环境。因此,我们要深入开展转基因作物对土壤生态系统的风险评估工作。转基因作物土壤生态系统安全风险的核心是要评估转基因作物的种植是否会改变土壤生态系统的功能。研究表明,土壤微生物多样性和功能之间没有内在的必然联系,物种丰富度的改变并不会对土壤功能产生实质性影响[75-76]。目前,只有极少数研究显示转基因作物可能会对土壤微生物功能多样性产生短暂的影响。如张卓等[63]利用Biolog微孔板法发现,抗草甘膦转基因大豆成熟期的土壤微生物碳源利用能力显著高于受体材料,但与另一常规种植品种无显著差异,到残茬期三者又均无显著性差异了。
迄今为止,已有大量的研究评估了转基因作物对土壤微生物存在的潜在影响,但是由于这些研究的对象(包括外源基因和植物物种)、环境、实验方法等均不完全相同,所得数据也用不同的方法进行分析,再加之大多数的研究结果都只是在1-2年的栽培期的基础上得出的,因此结果必然会显示出一定的差异。在未来的分子育种实践中,可能还会开发出更多的多性状叠加的转基因作物,这会对转基因环境安全评价工作带来更多的挑战,研究内容也会更加复杂。正是由于当前研究具有上述这些局限性,我们在今后的工作中,更应当重视土壤微生物群落结构组成和功能之间相关性及其对土壤系统自然变化(如季节、气候、轮作和农药使用)的响应等方面的研究,从而能够更加全面、准确的评估转基因作物对土壤微生物的影响程度。
转基因作物的安全风险评估是一项长期而艰巨的任务,我们相信随着时间的推移,人们可以更好、更全面地了解转基因作物种植的利弊,为人类的粮食安全问题做出贡献。
