长链非编码RNA-EPS 对小鼠牙釉质发育的影响
2021-11-05郝晨笛苏俭生
郝晨笛, 苏俭生
(上海牙组织修复与再生工程技术研究中心,同济大学口腔医学院,同济大学附属口腔医院口腔修复科,上海 200072)
牙釉质是在多种细胞因子调控下,由成釉细胞合成、分泌有机基质并不断矿化成熟形成的高度矿化组织[1]。 成熟的牙釉质由95%~97%的无机物和小于1%的有机物组成,是无细胞性组织,因而无法进行生理性自行修复[2]。 在釉质基质形成及矿化的过程中, 如基因调控出现异常而引起遗传学水平改变,则会导致牙釉质结构异常,即釉质发育缺陷性疾病[3]。 之前对釉质发育的分子机制研究大多围绕着编码釉基质蛋白的牙源性基因[4],如釉原蛋白 (amelogenin,AMELX)、 釉 蛋 白 (enamelin,ENAM)、成釉蛋白(ameloblastin,AMBN)、釉成熟蛋白(amelotin,AMTN)基因,以及编码釉基质蛋白水解酶的基因,如金属基质蛋白酶-20(matrix metalloproteinase-20,MMP-20) 和激肽释放酶 4(kallikrein 4,KLK4)基因进行研究。 此外,BMP 家族、Wnt 家族等信号通路与牙发育的研究也取得了重大进展[5],但釉质发育的具体调控网络和机制还有待探究。
随着高通量技术的发展,人们发现,原本认为不具有蛋白质编码功能的非编码RNA(non-coding RNA,ncRNA) 可通过转录或转录后机制来调控基因的表达。 MicroRNA(miRNA)和长链非编码RNA(long noncoding RNA,lncRNA) 作为 2 种调控性非编码RNA,已被证实广泛参与了细胞周期与分化等多个生物学过程。 近年来, 越来越多的证据表明,miRNA 是牙齿发育信号通路的重要调节因子[6-8]。Funada 等[9]发现,miR875-5p 通过血小板衍生生长因子(plateletderived growth factor,PDGF)信号通路诱导间充质细胞向牙上皮细胞迁移并凝结,参与上皮-间充质相互作用,从而调控成釉细胞分化及釉质形成等牙齿形态发生过程。 2011 年,Salmena 等[10]提出,lncRNA 可作为竞争性内源RNA(competitive endogenous RNA,ceRNA) 结合 miRNA, 从而调控miRNA 对mRNA 的抑制作用, 最终实现对细胞功能的调控。 此前已有研究证实,lncRNA 参与了骨形成等器官发育过程[11],但lncRNA 是否参与调控牙发育还未见报道。 本研究旨在通过对比观察lincRNA-EPS 基因敲除型和野生型小鼠下颌磨牙的差异,探究基因间长链非编码RNA lincRNA-EPS 对牙釉质发育的影响。
1 材料和方法
1.1 材料
1.1.1 实验动物 lincRNA-EPS 基因敲除纯合型C57BL/6J 小鼠(knockout,KO)由上海南方模式生物科技股份有限公司提供,C57BL/6J 小鼠 (wild type,WT)由同济大学实验动物中心提供。 实验动物均饲养于上海牙组织修复与再生工程技术研究中心SPF级实验动物房中。选用雄性及雌性8~10 周龄KO 组和WT 组小鼠,按1∶2 的比例将2 组小鼠分别合笼,胎龄计算以新生小鼠出生当日中午为出生后(postnatal,PN)第 0.5 天。选用 PN0.5 及 8 周龄 KO 和 WT小鼠用于本研究。 经同济大学附属口腔医学院伦理委员会许可对实验动物进行操作(编号:〔2018〕伦审字 026 号)。
1.1.2 实验试剂及设备 树脂包埋剂(Technovit公司,德国),TRIzol 试剂盒(TaKaRa 公司,日本),逆转录试剂盒(TaKaRa 公司,日本);micro-CT 机(SCANCOMedical 公司, 瑞士), 体视显微镜(Leica公司,德国),扫描电子显微镜(Hitachi 公司,日本),X 射线能谱仪 (IXRF 公司, 美国), 实时荧光定量PCR 仪(罗氏公司,瑞士)。
1.2 方法
1.2.1 小鼠基因型鉴定 剪取PN0.5 KO 和WT 小鼠约1 mm 鼠尾片段, 小鼠基因组使用引物对P1/P2,P3/P4 分别进行扩增,聚合酶链式反应(PCR)鉴定 lincRNA-EPS 基因。 PCR 反应条件:98 ℃预变性2 min,98 ℃ 10 s,63 ℃ 15 s,68 ℃ 1 min,34 个循环 后 68 ℃ 延 伸 5 min。 P1 引 物 序 列 :5′ -CCAGGATGTCAGAAGAGGAAGC-3′;P2 引物序列:5′-ACACAGAGCAGC-ATAAGGACA-3′;P3 引 物 序列 :5′-AAGGGACA-GGAGGAGGGATTGG-3′;P4 引物序列:5′-CTAG-GGCATACTGAGGAGGTTTTG-3′。
1.2.2 取材 取8 周龄雄性KO 和WT 小鼠, 将其颈椎脱臼处死并完整解剖分离其双侧下颌骨(含牙),剃净附着软组织后用磷酸盐缓冲液(phosphate buffered saline,PBS)冲洗 3 次,置于 4%多聚甲醛溶液中以4 ℃固定24 h,于体视显微镜下观察并拍摄其外形, 随后长期保存于0.5%多聚甲醛溶液中,用于后续 X 线检查、micro-CT 扫描、 扫描电镜(SEM)分析及 X 射线能谱仪(EDX)分析。 选取 PN0.5 同窝出生的KO 小鼠和WT 小鼠各6~9 只, 体视显微镜下解剖分离小鼠双侧下颌第一磨牙牙胚,用于实时定量聚合酶链式反应(RT-qPCR)检测。
1.2.3 X 线和micro-CT 扫描 将固定完成的8 周龄KO 和WT 小鼠下颌骨放置于micro-CT 样本扫描管中进行扫描(管电压:70 kV;管电流:200 μA;管精度:14.4 μm;曝光时间:300 ms),扫描完成后通过软件对样本进行三维重建及分析。
1.2.4 RT-qPCR 检测 用 TRIzol 法分别提取PN0.5 KO 和WT 小鼠下颌第一磨牙牙胚总RNA,使用TaKaRa 逆转录试剂盒将其逆转录合成cDNA;参照 SYBR 体系, 配制 RT-qPCR 反应体系。 以GAPDH 作为内参对照,置于RT-qPCR 仪上反应,检测KO 和WT 小鼠牙胚中釉质发育相关基因AMELX、ENAM、AMBN、AMTN 的 mRNA 表达水平差异。 引物序列见表1。

表1 釉质发育相关基因引物序列表Table 1 Primer sequence of amelogenesis related genes
1.2.5 SEM 分析 固定好的8 周龄KO 和WT 小鼠下颌骨用PBS 冲洗3 遍, 去离子水超声荡洗5 min,干燥,表面喷金,SEM 分析其表观形貌。扫描完成的样本用 PBS 冲洗, 用 50%、70%、80%、90%和 100%的乙醇溶液梯度脱水后浸入树脂包埋剂中,光固化13.5 h 形成树脂包埋块。 以下颌磨牙近远中方向为轴抛光至观察层面,去离子水荡洗5 min,37%磷酸酸蚀30 s,5.25%次氯酸钠浸泡1 min, 去离子水超声荡洗10 min,干燥,喷金,SEM 观察釉质超微结构。
1.2.6 EDX 分析 固定好的 8 周龄 KO 及 WT 小鼠下颌骨用PBS 冲洗 3 遍, 去离子水超声荡洗10 min,将样本固定在标本载物台上,每个样本选择3 个扫描位点,用X 射线能谱仪检测其元素组成。
1.2.7 统计学分析 使用SPSS 20.0 软件对所得数据进行统计学分析,各组数据经过正态分布及方差齐性检验后使用。 采用t 检验法分析比较2 组之间的差异,P<0.05 表示差异具有统计学意义。
2 结果
2.1 PCR 鉴定转基因动物基因型
PCR 产物电泳鉴定结果如图1 所示。WT 小鼠扩增出382 bp 条带,lincRNA-EPS KO 小鼠扩增出825 bp条带。经鉴定,通过繁殖获得的纯合型lincRNA-EPS缺失的子代可用于后续实验。
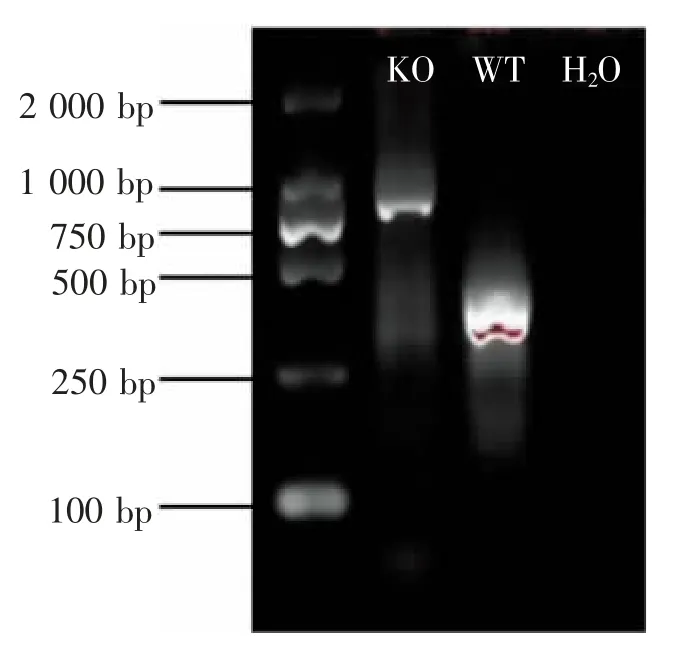
图1 PCR 产物电泳结果Figure 1 Electrophoresis of PCR products
2.2 lincRNA-EPS 基因敲除导致小鼠牙釉质缺损
8 周龄KO 和WT 小鼠下颌骨体视显微镜下图像及micro-CT 三维重建图像(图2)显示,与WT小鼠相比,KO 小鼠下颌磨牙舌侧出现大面积牙体组织缺损,且缺损程度由近中向远中方向逐渐降低,釉质色泽和透明度未见明显差异。 X 线图像显示,KO 和WT 小鼠下颌骨X 线透光密度无明显差异。
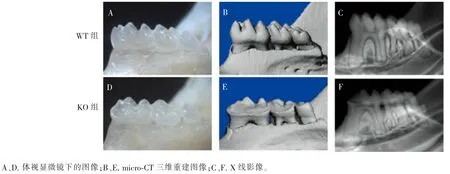
图2 8 周龄KO 和WT 小鼠下颌磨牙大体观察Figure 2 Microscopic evaluation of the mandibular molars from KO and WT mice at 8-week-old
2.3 lincRNA-EPS 调控釉质发育相关基因的表达
RT-qPCR 结果如图 3 所示, 与 WT 组小鼠相比,KO 组小鼠釉质发育相关基因AMELX、AMBN、ENAM、AMTN 的 mRNA 表达量均显著下调 (P<0.05)。
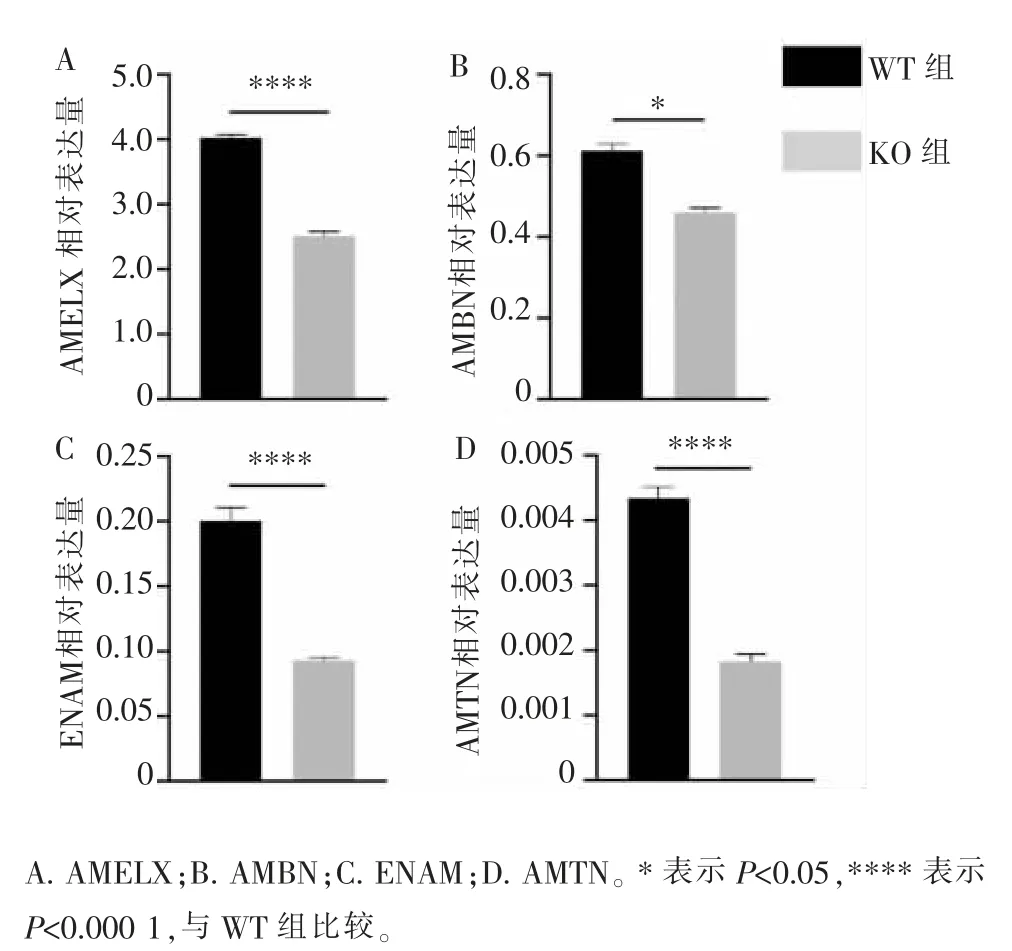
图3 KO 和WT 小鼠釉质发育相关基因mRNA 表达水平Figure 3 mRNA expression level of amelogenesis-related genes in KO and WT mice
2.4 lincRNA-EPS 缺失导致小鼠釉质表面形貌及微观结构改变
WT 和KO 小鼠下颌磨牙表面形貌扫描电镜图像如图4 所示。WT 小鼠牙体硬组织结构完整,釉质表面光滑平坦, 牙尖可见正常生理性磨损;KO 小鼠牙体硬组织大面积缺损,第一、第二、第三磨牙舌侧正常牙尖结构(L1、L2、L3)均出现一定程度的磨耗缺损, 第一磨牙甚至出现凹坑状缺损。 放大图像(图4D)中见KO 小鼠第一磨牙釉质边缘结构粗糙,牙本质暴露,可见数条沟壑状裂纹(箭头处)。
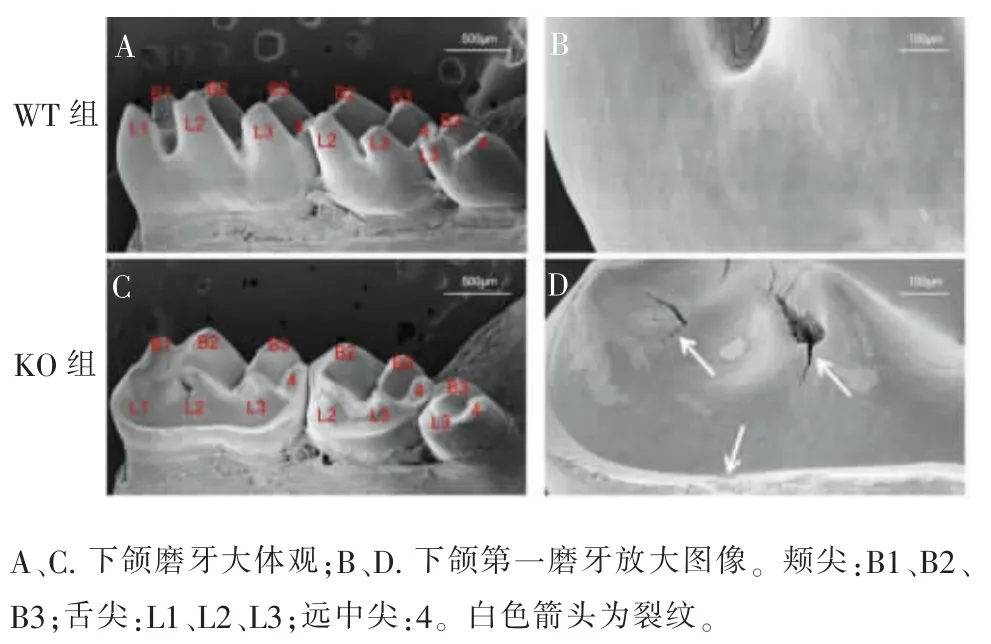
图4 8 周龄WT 与KO 小鼠下颌磨牙表面形貌分析(×5)Figure 4 Morphology of mandibular molars from WT and KO mice at 8-week-old (×5)
WT 及KO 小鼠下颌磨牙蚀刻后釉质区横截面SEM 图像如图 5A、D 所示,WT 小鼠下颌磨牙釉柱排列整齐有序,呈螺旋状交替;KO 小鼠下颌磨牙釉柱排列紊乱,且不连续,呈断裂状(箭头处)。 WT 及KO 小鼠下颌第一、 第二磨牙邻接区釉质图像如图5B、E 所示,WT 小鼠釉质边缘无釉柱结构光滑平整,KO 小鼠无釉柱结构被破坏, 表面粗糙 (箭头处)。 放大图像(图 5C、F)下见 WT 小鼠单个釉柱内羟基磷灰石晶体结构排列紧密有序,KO 小鼠釉柱内羟基磷灰石晶体间出现间隙(箭头处)。
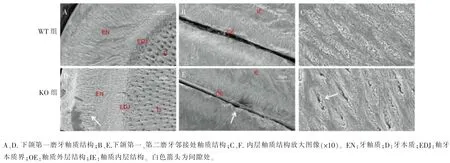
图5 8 周龄WT 和KO 小鼠下颌磨牙横断面釉质结构分析Figure 5 SEM micrographs of enamel cross sections of mandibular molars from WT and KO mice at 8-week-old
2.5 lincRNA-EPS 的缺失对釉质表面钙/磷比无影响
EDX 分析结果如表 2 所示。 KO 组和 WT 组小鼠牙釉质中含有均匀分布的钙、磷等微量元素,2 组小鼠间釉质的钙/磷质量百分比和原子百分比差异均无统计学意义(P>0.05)。
表2 WT 和 KO 小鼠釉质 EDX 分析结果()Table 2 EDX analysis of enamel in WT and KO mice()

表2 WT 和 KO 小鼠釉质 EDX 分析结果()Table 2 EDX analysis of enamel in WT and KO mice()
组别 EDX 分析结果钙/磷质量百分比/wt% 钙/磷原子百分比/at%WT 组 2.16±0.16 1.62±0.17 KO 组 2.29±0.09 1.65±0.15 P 值 0.07 0.66
3 讨论
Hu 等[12]在 2011 年首次发现 lincRNA-EPS,并获取其全长序列, 对该基因进行功能注释后发现,lincRNA-EPS 具有抗凋亡的活性, 此后对lincRNAEPS 的研究大多集中在免疫炎症调节方面。 为探究lincRNA-EPS 是否参与牙发育过程, 我们前期通过CRISPR/Cas9 技术构建了 lincRNA-EPS 基因敲除小鼠模型, 对小鼠下颌磨牙进行表型研究, 发现lincRNA-EPS 基因敲除小鼠的牙齿数量、 萌出时间与野生型小鼠相比没有差异, 但成年lincRNA-EPS基因敲除小鼠下颌磨牙区釉质出现大面积缺损,提示lincRNA-EPS 参与了小鼠下颌磨牙牙釉质发育及稳态维持过程[13]。
哺乳动物牙釉质的发育是在多种信号分子和转录因子有序诱导下完成的动态过程[14], 经过了2 个相互交叠的时期:分泌期和成熟期。 确定牙釉质发育的影响因子及调控机制对于了解牙釉质发育过程及预防釉质发育缺陷性疾病具有重要意义,还可为牙齿再生的研究提供理论基础。 在釉质形成的分泌期,成釉细胞合成并分泌釉基质蛋白(enamel matrix proteins,EMPs)直至达到应有的厚度。 羟基磷灰石(hydroxyapatite,HAP)晶体是组成牙釉质的主要成分。 研究表明,EMPs 在釉质分泌期诱导HAP 晶体沿c 轴生长至全长,从而参与控制其形成的大小形态、生长方向及生长速度[15]。Fan 等[16]发现,釉原蛋白和釉蛋白还可通过合作效应控制大分子自组装,从而影响晶体形态。 为探究lincRNA-EPS 是否通过对EMPs 的分泌产生阻碍从而影响釉质发育, 本研究对处于EMPs 分泌成熟交叠时期的磨牙牙胚进行mRNA 表达量分析。 与野生型小鼠相比,lincRNAEPS 基因敲除小鼠中 AMELX、ENAM、AMBN、AMTN的 mRNA 表达量均显著降低(P<0.05),说明 lincRNAEPS 参与了EMPs 的分泌成熟过程, 并推测其参与调控了釉质晶体的生长。
当釉质发育进入成熟期,成釉细胞分泌基质蛋白酶MMP-20 和KLK4 降解EMPs,釉质中的有机成分逐渐被无机成分所代替,无机成分最终可达釉质重量的95%~97%[17]。 HAP 作为釉质中主要的无机物, 其交织排列组成了釉质的基本结构——釉柱,釉柱又按一定规律紧密排列, 旋转扭曲成束状结构[18],这样的化学成分和结构组成使釉质拥有了能够长期使用的机械强度。 为探究lincRNA-EPS 基因敲除小鼠牙釉质结构是否发生改变,通过扫描电镜对釉质超微结构进行分析,发现lincRNA-EPS 基因敲除小鼠下颌磨牙釉柱排列紊乱,螺旋状的有序结构被破坏,釉柱内的HAP 晶体排列出现间隙,釉质外层无釉柱结构区域遭到破坏。 以上研究结果表明,lincRNA-EPS 的缺失造成EMPs 表达的下调,并可能因此影响了其成熟时的吸收过程, 造成HAP晶体及釉质排列紊乱。 由于无序的牙釉质结构会使釉质的应力强度大大降低,因此,在进行咀嚼、咬合等正常生理活动时,lincRNA-EPS 基因敲除小鼠的釉质结构更易被破坏, 加速了牙体组织的磨耗,甚至会出现与人牙隐裂相似的症状。 为进一步探讨lincRNA-EPS 缺失对小鼠釉质矿化的影响, 本研究从X 线影像和EDX 2 个层面进行矿化分析。 研究结果显示,2 组小鼠矿化程度及钙/磷比值间差异无统计学意义,说明lincRNA-EPS 基因敲除后,小鼠釉质晶体的排列虽然发生了变化,但晶体的原子结构及原子量并未改变。
综上所述,lincRNA-EPS 的缺失影响了釉基质蛋白分泌和吸收的过程, 并引发牙釉质结构的紊乱,最终导致了釉质结构的缺损。 研究首次证明了lincRNA-EPS 在牙釉质发育过程中的调控作用,为进一步探究釉质发育机制提供了重要研究基础。
