初级纤毛在长骨生长板分布规律的小鼠实验研究
2021-11-05丁允鹏陶狄坷
丁允鹏, 陶狄坷, 张 帅, 孙 瑶
(上海牙组织修复与再生工程技术研究中心,同济大学口腔医学院,同济大学附属口腔医院口腔种植科,上海 200072)
初级纤毛是细胞表面的毛状感受器,存在于多种细胞类型中。 目前,初级纤毛已经被证实能够感受化学信号、机械力,并且广泛参与多种组织器官的发育,能够调控细胞增殖、谱系分化和维持干细胞干性[1-2]。 初级纤毛相关基因突变引起的纤毛病,如 Verma-Naumoff 综合征、Senior-Loken 综合征、Meckel-Gruber 综合征等常出现短指、短肋等长骨发育畸形[3-5]。 在小鼠中敲除初级纤毛相关基因也会导致长骨发育畸形,如在表达转录因子7(transcription factor 7,Sp7) 的细胞中敲除纤毛转运蛋白 80(intraflagellar transport 80,Ift80)会抑制成骨细胞分化[6],在表达 2 型胶原(collagen 2,COL2)的软骨细胞中敲除Ift88 会导致小鼠四肢短小, 生长板结构紊乱[7]。 上述研究结果说明,纤毛对于长骨的生长发育有重要影响,但其具体机制还有待进一步研究。
长骨生长的方式为软骨内成骨,其生长主要依赖于长骨中特异性的生长板结构。 生长板由软骨细胞构成,生长板软骨细胞沿长骨长轴向两端增殖使长骨增长。 按照软骨细胞的生物学特征可以将生长板分为4 层:静息层、增殖层、前肥大层和肥大层[8]。已有研究证实,软骨生长板中有大量的细胞纤毛存在[9],但是初级纤毛在生长板各层的分布规律有待系统研究。 研究软骨中初级纤毛分布特点,对阐释生长板发育和稳态维持具有重要意义。 我们拟通过分析初级纤毛在1、4、8 周龄小鼠股骨生长板的分布规律来揭示纤毛和生长板发育及稳态维持的关系。
1 材料和方法
1.1 小鼠取材固定和切片
取 1、4、8 周龄野生型 C57BL/6J 小鼠,用 1%戊巴比妥钠对其麻醉后使用4%多聚甲醛进行体循环固定, 剔除软组织后分离双侧股骨,4%多聚甲醛固定48 h,10%乙二胺四乙酸 (ethylene diamine tetraacetic acid, EDTA)脱钙21 d。 脱钙完成后用清水冲洗 30 min,脱水浸蜡,石蜡包埋。 以 4 μm 的厚度进行石蜡切片, 随后进行甲苯胺蓝和免疫荧光染色。
1.2 甲苯胺蓝染色
将石蜡切片置于65 ℃烘箱60 min, 脱蜡并水化。 甲苯胺蓝染液(Sigma 公司,美国)室温孵育30 min,65 ℃烘箱烘干,二甲苯透明,中性树胶封片。
1.3 免疫荧光染色
用透明质酸酶37 ℃抗原修复60 min, 磷酸盐缓冲液 (phosphate buffered saline,PBS) 洗 3 次,0.5%Triton 室温打孔 10 min,PBS 洗 3 次,1%小牛血清封闭60 min,吸弃血清,按1∶200 的比例稀释并孵育COL2(博士德生物工程有限公司,中国),ADP核糖基化因子样蛋白13b (ADP ribosylation factor like GTPase13b,Arl13b) 或乙酰化微管蛋白(acetylated tubulin,ACE-tubulin)抗体抗体(Proteintech 公司,美国),4 ℃孵育过夜,用 PBS 洗 3 次,37 ℃孵育荧光二抗 60 min,PBS 洗 3 次,4′,6-二脒基-2-苯基吲哚(DAPI)复染,37 ℃孵育 5 min,用 PBS 洗 3 次,用防淬灭封片剂封片。
1.4 软骨细胞培养
取 2 只 1 周龄野生型 C57BL6/J 小鼠双侧股骨生长板,吹打洗去表面骨髓组织,将生长板置于4 mg/mL 的Ⅱ型胶原酶(Sigma 公司,美国)中,37 ℃消化 40 min,用 PBS 10 倍稀释,300×g 离心 10 min,3 mL PBS 重悬,70 μm 滤网过滤, 将过滤所得细胞悬液接种于60 mm 培养皿中培养并记为P0 代,将P1 代用于实验。
1.5 软骨细胞肥大化诱导
将P1 代软骨细胞以5×105个/孔的密度接种于6 孔板。肥大软骨细胞诱导液构成:1 nmol/L 地塞米松(Sigma 公司,美国);50 μg/mL 抗坏血酸(Sigma 公司,美国);100 μg/mL 丙酮酸钠;1%ITS 添加物(赛业生物科技有限公司,中国);1%双抗(凯基生物技术股份有限公司,中国);5 mmol/L 甘油磷酸钠(Sigma 公司,美国)。
1.6 细胞免疫荧光染色
4%多聚甲醛于 4 ℃ 下固定细胞 60 min,用PBS 冲洗 3 次,0.5%Triton×100 室温打孔 5 min,用PBS 液冲洗 3 次,1%小牛血清室温封闭 60 min, 按照 1∶200 的比例孵育 COL2,10 型胶原(collagen10,COL10)抗体(博士德生物工程有限公司,中国),Arl13b 抗体 (Proteintech 公司, 美国),4 ℃过夜,用PBS 洗 3 次, 孵育二抗,37 ℃孵育 60 min, 用 PBS洗 3 次,DAPI 复染,37 ℃孵育 5 min, 用 PBS 洗 3 次,将细胞爬片取出,于载玻片上封片。
1.7 RT-qPCR
用 TRIzol 法(TaKaRa 公司,日本)提取细胞总RNA,使用 PrimeScriptTMRT 试剂盒(TaKaRa 公司,日本)进行逆转录,然后使用SYBR Mix 和LightCycler96(Roche 公司,瑞士)进行 RT-qPCR 实验及分析, 以甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)作为内参(引物序列见表 1),通过 2-ΔΔCT法计算 mRNA 的倍数变化,并根据对照组对数据进行标准化。
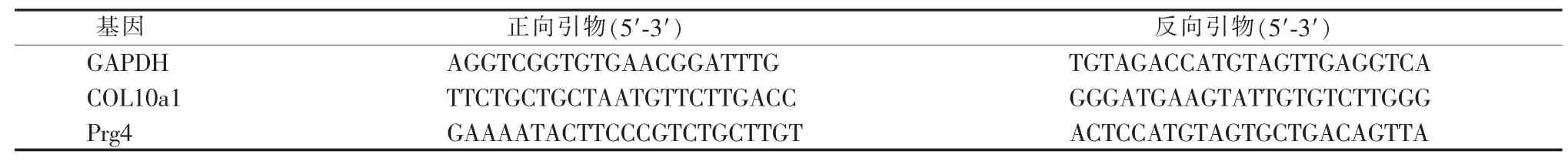
表1 RT-qPCR 引物序列Table 1 Primer sequence in RT-qPCR
1.8 统计学分析
统计学差异分析使用SPSS 22.0 软件进行两独立样本t 检验或方差分析,P<0.05 为差异具有统计学意义。
2 结果
2.1 长骨发育过程中生长板软骨细胞存在初级纤毛
通过甲苯胺蓝染色,我们发现小鼠股骨生长板在其 1、4 周时较厚,8 周时, 随着股骨发育接近尾声,股骨生长板逐渐变薄(图1A~C);在所有的时间点,生长板的软骨细胞均存在初级纤毛(图1D~J),随着年龄增长,生长板软骨细胞中的纤毛细胞比例降低。
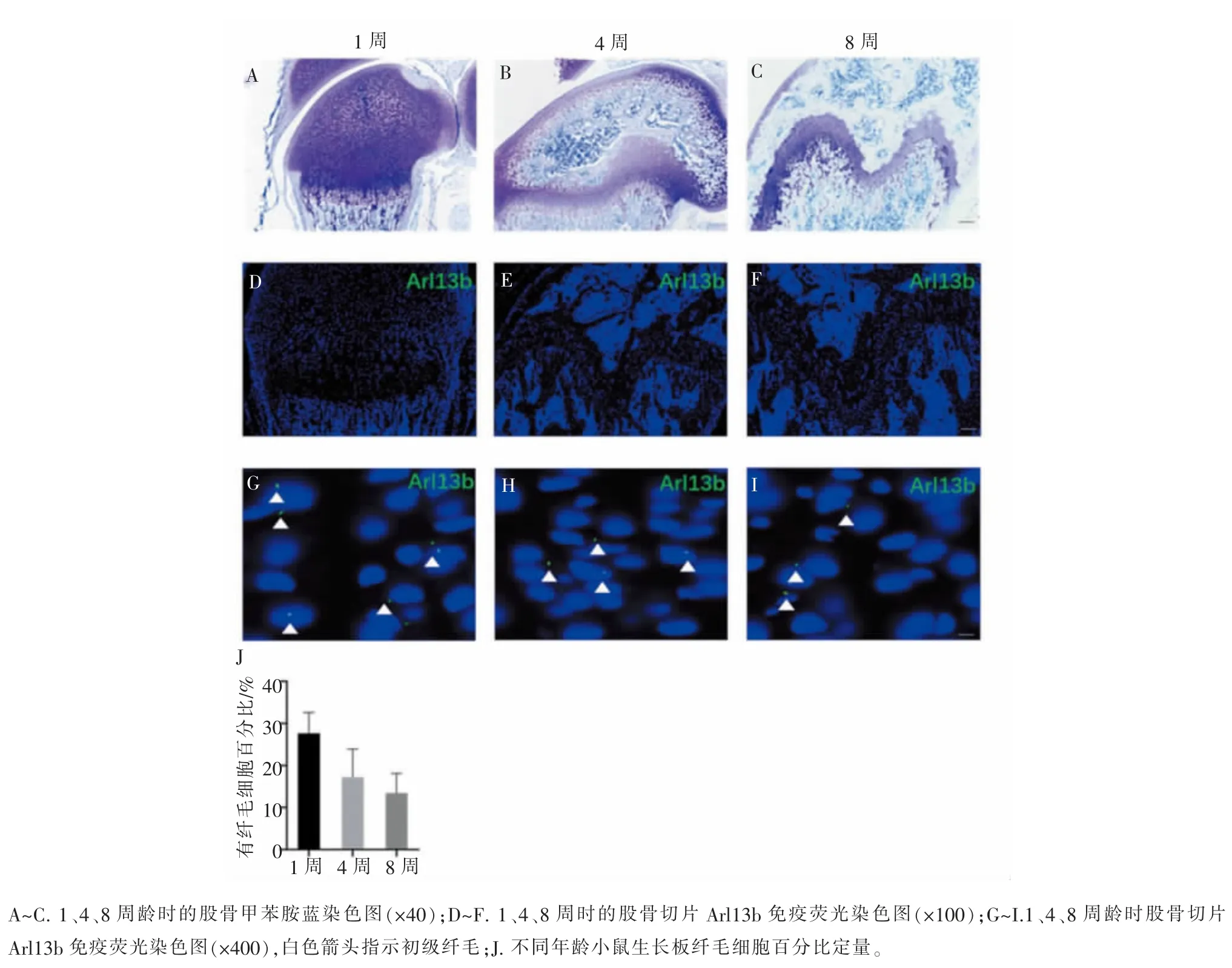
图1 长骨发育过程中生长板软骨细胞存在初级纤毛Figure 1 Primary cilia of chondrocyte in femur growth plate of developing mouse
2.2 初级纤毛主要存在于生长板静息层、增殖层和前肥大层
为了进一步研究初级纤毛在生长板的分布规律,我们通过免疫荧光染色共同观察软骨细胞标志物COL2 和初级纤毛标志物乙酰化微管蛋白(ACE-tubulin),结果发现,初级纤毛主要分布于静息层、增殖层和前肥大层,肥大层的肥大软骨细胞少有初级纤毛(图 2)。
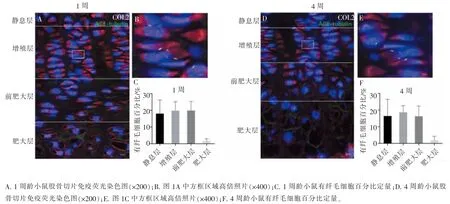
图2 有纤毛的软骨细胞主要存在于静息层、增殖层和前肥大层Figure 2 Ciliated chondrocytes mainly exist in resting zone, proliferative zone and prehypertrophic zone
2.3 生长板软骨细胞肥大化过程中初级纤毛略减少
为了研究初级纤毛在软骨细胞肥大化中的作用,我们在体外培养了软骨细胞并诱导其向肥大软骨细胞分化,7 d 后, 肥大软骨细胞特异性基因COL10、Prg4 表达升高,差异具有统计学意义,免疫荧光显示大部分细胞表达 COL10 (图 3A、B、D、E),肥大软骨细胞诱导成功。 同时我们发现,在诱导7 d后,初级纤毛细胞的比例略降低,但差异没有统计学意义(图 3I)。
图3 生长板软骨细胞肥大化过程中,初级纤毛略减少Figure 3 The primary cilia were slightly reduced during growth plate chondrocytes differentiated into hypertrophic chondrocytes
2.4 提高有纤毛细胞比例伴随肥大软骨细胞标志性基因的表达降低
本实验通过无血清饥饿软骨细胞48 h 阻滞细胞周期,显著提升了有纤毛细胞的比例(图4A~E),之后进行肥大化诱导,7 d 后RT-qPCR 和细胞免疫荧光结果显示,经过无血清饥饿的软骨细胞COL10 表达显著低于对照组,差异具有统计学意义,Prg4 的表达略低于对照组,但差异没有统计学意义(图4F、G)。
3 讨论
长骨生长板有一套维持稳态的机制,静息层的骨骼干细胞周期性产生软骨祖细胞,软骨祖细胞向增殖层迁移并快速增殖促进长骨沿长轴的生长,增殖层末端的软骨细胞向肥大软骨细胞分化,肥大软骨细胞分泌软骨基质为骨化提供条件[10-11]。 生理条件下,增殖层软骨祖细胞和肥大软骨细胞维持一定的比例,软骨祖细胞提前分化或增殖过度活跃都会导致骨骼发育异常[12]。 生长板增殖分化的调控依赖于印度刺猬因子(indian hedgehog,IHH)-甲状旁腺激素相关肽(parathyroid hormone-related peptide,PTHrP)负反馈机制,前肥大软骨细胞分泌的IHH 通过旁分泌作用促进软骨祖细胞的增殖, 静息层软骨祖细胞在IHH 的刺激下分泌PTHrP 抑制软骨祖细胞的增殖,并促进其向肥大软骨细胞分化[13]。这套负反馈机制能够抑制软骨的过度增殖或者分化,维持生长板的动态平衡。
Arl13b 是定位在初级纤毛的小GTP 酶,ACE-tubulin 是初级纤毛轴丝的组成部分,两者均是常用的初级纤毛标志物。 通过免疫荧光标记Arl13b 和ACE-tubulin,我们发现在 1、4、8 周龄小鼠长骨生长板中都存在初级纤毛,在1、4 周龄小鼠股骨生长板中初级纤毛主要存在于静息层、增殖层、前肥大层的软骨细胞,在分化成熟的肥大软骨细胞中则较少观察到初级纤毛。 小鼠股骨静息层和增殖层存在大量有纤毛的软骨细胞,与此前报道的软骨细胞增殖调控机制有关。 在出生后第9 天,前肥大软骨细胞分泌的IHH 能够促进静息层骨骼干细胞的产生[14],同时IHH 还可以刺激增殖层软骨细胞的快速增殖以维持生长板宽度[15],而经典的IHH-GLI 信号经过初级纤毛介导。 因此,在软骨细胞中敲除初级纤毛相关基因常导致生长板增殖层变短[16]。
初级纤毛在生长板增殖层的富集与其调控软骨细胞增殖相关,但当软骨细胞分化为肥大软骨细胞后,初级纤毛会消失,并且在体外诱导软骨细胞分化为肥大软骨细胞后, 有纤毛细胞比例显著降低,说明初级纤毛可能能够抑制软骨细胞向肥大软骨细胞分化, 其原因尚不清楚, 这可能与经典的Hedgehog-GLI 信号对软骨细胞的促增殖作用有关[17-19]。 Hedgehog-GLI 信号能够促进软骨细胞的增殖,而初级纤毛是介导HH 信号的关键细胞器。 向肥大软骨细胞分化的过程中,初级纤毛消失可能能够抑制软骨细胞增殖,为其向肥大软骨细胞分化提供条件,所以在软骨细胞分化过程中,初级纤毛会逐渐减少直至消失。 除此之外,我们发现有纤毛比例更高的软骨细胞经过肥大化诱导之后,肥大软骨细胞标志性基因表达更低,这也提示初级纤毛可能对软骨细胞向肥大软骨细胞分化有抑制作用。 以往的研究认为,生长板软骨细胞的增殖分化受生长板微环境中多种因子的调控[20],我们通过研究初级纤毛在生长板的分布规律发现,软骨细胞可能通过解聚初级纤毛来抑制IHH 的促增殖作用,促进自身向肥大软骨细胞分化,为研究生长板发育和稳态维持提供了新思路。
综上所述, 初级纤毛主要分布在生长板静息层、增殖层和前肥大层,分化成熟的肥大层软骨细胞初级纤毛消失。 通过无血清饥饿法提高有纤毛细胞的比例后,软骨细胞肥大化诱导过程中,肥大软骨细胞标志性基因表达降低。
