不同来源大白猪总产仔数近交衰退评估
2021-11-04史良玉王立刚张鹏飞莫家远王立贤赵福平
史良玉,王立刚,张鹏飞,莫家远,李 洋,王立贤,赵福平
(中国农业科学院北京畜牧兽医研究所,农业部动物遗传育种与繁殖(家禽)重点实验室,北京 100193)
近交是将有亲缘关系的个体进行交配。在畜牧生产中,群体规模有限,近交难以避免。近交水平的增加会导致基因组中有害纯合子的增加,产生近交衰退,导致后代生产性能降低[1]。目前众多研究表明,在不同物种中,近交会影响动物的生产性能[1-3],尤其是牛的繁殖性能[4-6],但近交对猪繁殖性能影响的研究相对较少[7]。传统的近交系数计算方法是基于完整的系谱记录信息。随着基因组数据的不断涌现,从基因组层面能找到共同祖先传递下来的相同单倍型片段,即连续纯合基因型片段(runs of homozygosity, ROH)。根据ROH片段提供的遗传信息可以有效的估计出个体实现的近交系数,这为计算个体近交系数提供了新的途径[8]。另外,还可以利用ROH对每条染色体或某个染色体片段进行特定基因组区段的近交系数估计,并进一步估计其对近交衰退的影响程度。
本研究分别使用系谱、全基因组SNP标记和ROH计算个体的近交系数,并利用混合线性模型检测近交系数与加系和法系大白猪群体的总产仔数(total number of piglets born, TNB)的相互关系,从而评估近交衰退程度,并进一步定位导致大白猪总产仔数近交衰退具体的染色体区域,为基因组选种和选配提供参考。
1 材料与方法
1.1 试验材料
本研究选取上海某公司1 937头大白猪,其中1 039头 来自加系,898头来自法系,且两群体无亲缘关系。加系和法系群体分别饲养在两个不同的场,且两场营养管理水平基本相同。两群体系谱共由3 086头大白猪组成,系谱记录时间为2012—2019年,大约8个世代。仅对有芯片且有系谱的1 937头 大白猪个体进行近交系数的计算及近交衰退分析。
1.2 基因型数据及质控
对1 937头大白猪使用GeneSeek GGP Porcine HD 芯片进行分型。由于该芯片根据Susscrofa10.2版本设计,将其更新为Susscrofa11.1版本进行后续分析。使用PLINK v1.90软件[9]进行质控,质控条件如下:个体基因型检出率>90%,标记基因型检出率>90%,最小等位基因频率>0.05,仅保留常染色体SNP标记。质控后剩余37 310 个SNPs,1 937头大白猪。
1.3 近交系数估计
1.3.1 基于ROH的近交系数估计 使用PLINK v1.90软件[9]进行ROH检测。ROH的鉴定标准为:1)ROH的最小长度为1 Mb;2)每个ROH至少由45个SNPs组成,该参数计算公式由Lencz等[10]提出:
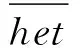
基于ROH估计的近交系数(FROH)计算公式为:
其中,LROHi为个体i的ROH长度,Lauto为芯片中常染色体SNPs覆盖长度。
1.3.2 基于SNPs的近交系数估计 基于SNPs估计的近交系数(FGRM)计算公式[11]为:
其中,xi为第i个SNP主等位基因的数量,m为SNPs数量,pi为主等位基因频率。
1.3.3 基于系谱的近交系数估计 使用BLUPF90家族程序(http://nce.ads.uga.edu/wiki/doku.php?id=application_programs)对系谱进行检测校正。校正后的系谱可追溯世代数为1~8代,平均世代数为5.33代。使用R pedigreemm 包[12]利用校正后的系谱估计近交系数(FPED)[13]。
1.4 近交衰退分析
由于加系和法系大白猪拥有不同的遗传背景,因此本研究使用DMU软件分别对两个品系大白猪的总产仔数进行近交衰退评估,采用分析模型如下:
yijk=μ+YSi+Pj+βFk+ak+Pek+eijk

2 结 果
2.1 表型数据描述性统计
对于加系群体,共有4 710条产仔记录,平均胎次为3.220;法系群体共有3 546条产仔记录,平均胎次为3.049。两品系总产仔数基本统计量见表1。

表1 两品系总产仔数基本统计量
2.2 两品系近交系数估计
如表2所示,对于两个品系,基于基因组信息估计的近交系数(FROH和FGRM)均值均高于基于系谱估计的近交系数(FPED)。加系群体FROH和FPED均值均高于法系群体,FGRM均值低于法系群体。在两个品系中,不同方法估计的近交系数之间的皮尔逊相关均为正相关(表3)。对于加系群体,FROH与FPED之间的相关最高,相关系数为0.358;对于法系群体,FROH与FGRM之间的相关最高,相关系数为0.371。

表2 基于基因组信息和系谱估计的近交系数分布

表3 不同方法估计的近交系数的皮尔逊相关
两品系近交系数随个体出生年份变化如图1所示。由于2014年仅1头个体出生,故在趋势分析中不考虑。2015—2019年间出生的个体,随着年份的增加,两品系FPED均呈现上升趋势,且加系群体FPED均高于法系群体。FROH波动趋势基本相同, 2018—2019年出生个体的FROH高于2015—2017年出生个体。对于加系群体,FGRM随着个体出生年份增加呈现上升趋势;对于法系群体,FGRM随着个体出生年份增加呈现先下降再上升趋势。
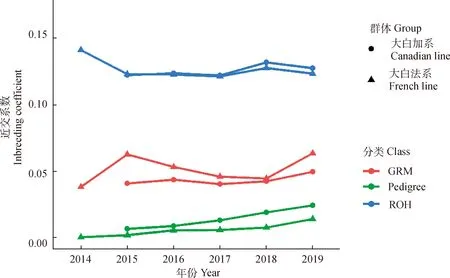
图1 两品系2014—2019年出生个体不同方法估计的近交系数Fig.1 The inbreeding coefficients for pig born between 2014 and 2019 estimated by different methods
2.3 两品系大白猪不同染色体及片段近交衰退估计
基于基因组近交系数(FROH及FGRM)和系谱近交系数(FPED),利用混合线性模型对大白猪总产仔数的近交衰退进行评估,以近交系数这一协变量的回归系数β作为近交衰退效应大小,结果见表4。尽管不同方法估计近交系数所得近交衰退效应大小不同,但随着近交系数的增加,加系群体总产仔数均显著减少。FPED每增加10%,加系群体总产仔数减少(0.823±0.276)头。FROH每增加10%,加系群体和法系群体总产仔数分别减少(0.571±0.178)、(0.690±0.324)头。

表4 两品系近交系数每增加10%总产仔数近交衰退估计
使用FROH对每条常染色体进行两个品系总产仔数近交衰退估计,结果显示,加系群体第6、7、8和13号染色体效应显著,而法系群体未发现显著产生近交衰退的常染色体(图2)。为了检测加系群体中导致近交衰退特定的基因组区域,将对随着近交系数增加总产仔数显著降低的染色体进行分段分析。根据第6、7、8和13号染色体在芯片中覆盖的长度分别平均分成2、4、6、8条片段,利用在相应片段内的FROH进行近交衰退分析。不同染色体不同片段结果如表5、表6、表7、表8所示,当染色体分割成2个片段时,6号和8号染色体的第一段显著,7号染色体第一段极显著。当将染色体分割为8条片段时,对于6号染色体,仅第一条片段效应显著,该片段位于6号染色体0.052~21.347 Mb之间;对于7号染色体,第三条片段(30.445~45.667 Mb)效应极显著,第六条片段(76.112~91.335 Mb)效应显著;对于8号 染色体,第一条片段(0.075~17.360 Mb)效应极显著,第三条片段(34.719~52.079 Mb)效应显著,第五条片段(69.438~86.798 Mb)效应显著。无论将染色体分割为几段,第13号染色体中的片段均不显著。

表5 加系群体6号染色体不同区域总产仔数近交衰退估计

表6 加系群体7号染色体不同区域总产仔数近交衰退估计

表7 加系群体8号染色体不同区域总产仔数近交衰退估计
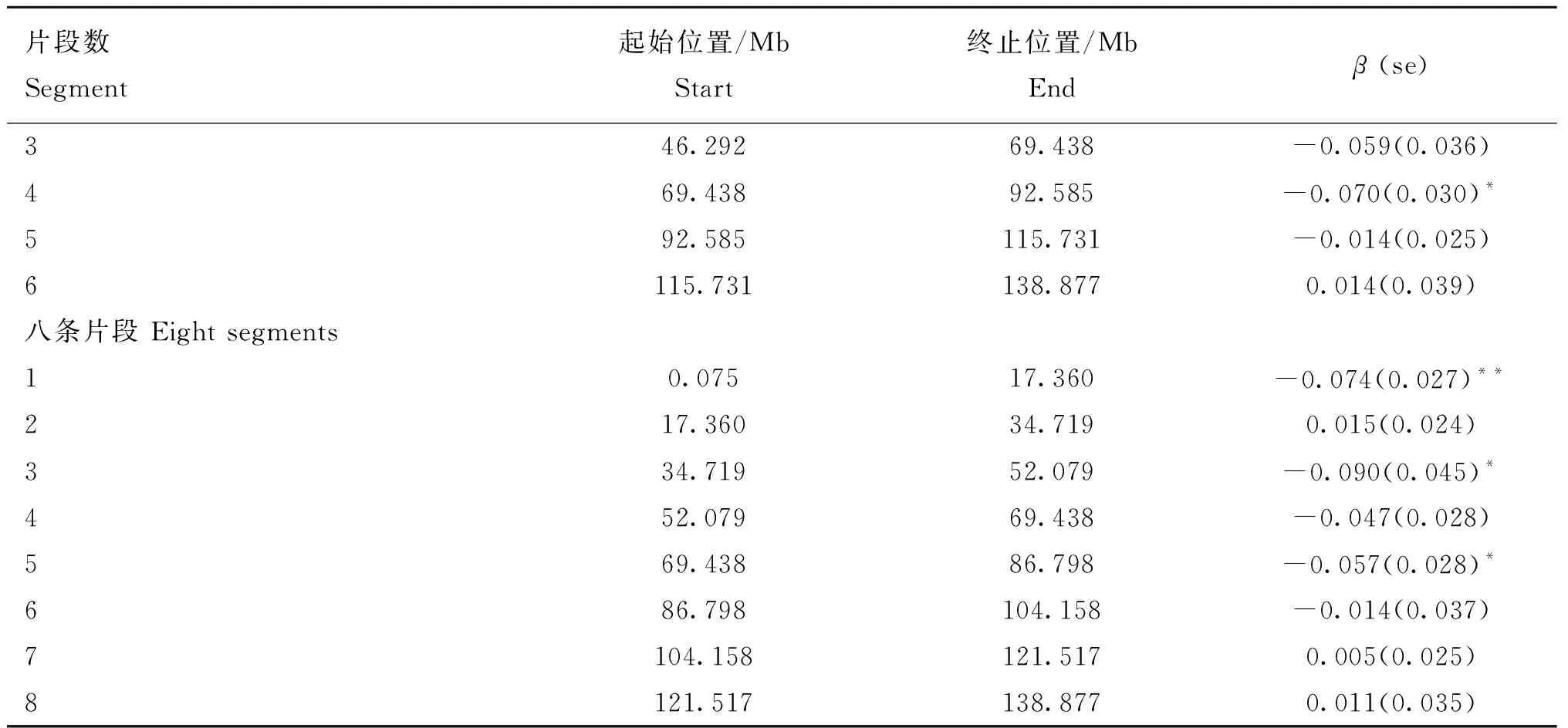
(转下页 Carried forward)

表8 加系群体13号染色体不同区域总产仔数近交衰退估计

图2 加系群体和法系群体总产仔数每条染色体每增加10% FROH近交衰退估计值及95%置信区间Fig.2 Inbreeding depression estimation in total number of piglets born per 10% increase in FROH and 95% confidence interval across autosomes for Canadian line and French line
3 讨 论
本研究采用了3种计算近交系数的方法对大白猪总产仔数的近交衰退进行评估。FPED的计算完全依赖于系谱信息,而实际生产中系谱常会出现记录错误和缺失的现象[14-15],并且假定基础群个体间无亲缘关系,这些都可能会低估近交水平。利用SNP计算近交系数时,很难辨别SNP是否是同源相同(identical by descent, IBD)还是同态相同(identical by state, IBS),常会高估近交水平[16]。另外,SNP对最小等位基因频率更加敏感,可能会导致FGRM估计准确性降低[17]。当有基因型的个体较少时FGRM估计不够准确,这会降低近交衰退估计的统计能力[18]。ROH是父母将来自共同祖先同一段单倍型传递给后代所形成的,所以ROH一般都属于IBD单倍型片段。因此,ROH近交系数既能避免系谱近交系数低估情况,又能避免SNP近交系高估的问题。而且FROH主要基于个体的基因组数据,不再依赖于系谱记录,能提供更准确的近交系数[19]。另外ROH还能够精细的定位因近交而造成衰退的特定基因组区段。所以,FROH已被认为是评估畜禽亲缘关系和近交系数的一种有效且准确的替代方法[20]。
本研究中法系群体FROH与FGRM与之间的相关高于其他对大白猪的研究报道[21],可能是由于本研究中大白猪群体产生了遗传漂变。尽管相比系谱使用基因组数据能够提高亲缘关系估计的准确性[22],但是FGRM有可能会得到负值。随着个体出生年份的增加,两品系FPED呈上升趋势。系谱中个体最早出生于2012年,而且假设基础群个体间无亲缘关系,因此2015年出生个体近交程度相对较低。法系群体在2018年之前使用系谱进行选择,自2018年起使用基因组选择,选择更加准确,可能会使基因型的同质性增加。另外,法系群体相对较小,利用SNP进行近交系数计算时,由于该方法对最小等位基因频率敏感,使近交系数有波动的现象。本研究群体中并未再引种而导入外源血统,FPED会随着年份的增长而增加。FROH与FPED之间的相关在两个品种中较低,分别为0.358、0.275,该结果与先前其他研究一致[23-24]。加系群体FROH与FPED与之间的相关高于法系群体,可能是加系群体具有更加完整的系谱记录,可追溯的平均世代数为5.493。
目前关于猪繁殖性状近交衰退的研究较少。周平等[25]根据近交系数在0~20%范围内将大白猪分为4组,结果表明,近交程度对相同胎次产仔数的影响不大,与胎次关系较为密切。但该研究仅采用简单的方差分析,未能消除胎次等效应因近交对产仔数的影响。Miglior等[26]使用系谱对奶牛泌乳期平均体细胞评分进行近交衰退估计,结果表明,近交系数每增加1%奶牛泌乳期平均体细胞评分增加0.012。已有研究利用SNP和ROH进行近交系数估计,从而评估近交衰退程度,其中仅少数几个研究定位了近交衰退片段[27-30]。Saura等[16]利用ROH计算109头母猪近交系数并鉴定到13号染色体一个近交衰退片段。本研究检测出13号染色能引起加系大白猪群体的总产仔数产生近交衰退现象,但未能检测到具体的染色体片段。分析原因有:1)在13号染色体86.188~160.936 Mb 每个个体均未检测到ROH;2)本研究只是按照染色体的长度将其平均划分为2~8段,这种分段方法也可能使原有近交衰退片段被打断。
本研究使用BioMart工具(ensembl.org/bio-mart)进一步对第6、7和8号染色体上存在近交衰退的ROH片段进行基因注释,共注释到601个基因。其中与繁殖性状相关的功能基因有5个,分别位于7号染色体30.5~45.7 Mb的ROH区段中的CUL7、MAPK14、PPARD基因和位于8号染色体69.4~86.8 Mb处ROH片段上的AREG、EREG基因。研究发现,CUL7[31-32]、MAPK14[33]和PPARD基因[34-35]与胎盘发育相关,AREG和EREG基因与卵母细胞成熟相关[36-38]。由于近交导致以上基因中隐性有害纯合子的增加,造成总产仔数的下降。
4 结 论
本研究针对两个不同背景来源的大白猪在经过约8个世代的选育后,利用基于系谱和基因型数据信息的3种方法估计近交系数,并评估近交对总产仔数的负面影响。加系和法系大白猪FROH的平均值分别为0.124、0.123。随着个体出生年份的增加,FPED也随之呈现上升趋势。对于加系群体,3种方法都检测到随着近交系数的增加,大白猪总产仔数呈现不同程度的降低,FROH与FPED结果更为接近。而在法系群体中,仅FROH检测到了近交衰退。利用FROH还检测到4条染色和多个特定染色体区域存在近交衰退的现象。本研究不仅为系谱数据缺失的群体,开展近交衰退估计提供新的研究方法,也为避免近交实施猪基因组选种选配工作提供了参考依据。
