外泌体非编码RNA在肝纤维化中的作用及机制
2021-11-04钱南南唐露露魏涛华郝文杰杨文明
钱南南, 唐露露, 魏涛华,, 杨 悦, 郝文杰, 杨文明,3
1 安徽中医药大学 研究生院, 合肥 230038; 2 安徽中医药大学第一附属医院 神经内科, 合肥 230031;3 新安医学教育部重点实验室, 合肥 230038
肝纤维化是由多种病因(如血吸虫、慢性病毒性肝炎感染、非酒精性脂肪性肝病、酒精性肝病、铜代谢性疾病、胆汁淤积和自身免疫性肝病)引起的慢性肝损伤,可逐渐发展为肝硬化,甚至引起肝癌的发生。近些年研究发现,肝纤维化是一个可逆转的过程,给肝纤维化的临床治疗带来了希望。肝纤维化发展过程中主要是由肝星状细胞(HSC)活化为成纤维细胞,并分泌细胞外基质(ECM)。因此,控制HSC的活化过程将是肝纤维化的理想治疗策略。大量研究表明,非编码RNA(ncRNA)对HSC活化、增殖、迁移、转化等过程起着重要作用。ncRNA主要包括长链非编码RNA(lncRNA)、短链非编码RNA和环状RNA(circRNA)等几大类,其中与肝纤维化相关的主要有:lncRNA包括MALAT1、H19、GAS5、MEG3、PVT1和P21等;短链非编码RNA包括微小RNA(miRNA)、小核仁RNA(snoRNA)、PIWI-interacting RNA(piRNA)和小干扰RNA(siRNA)等。细胞外囊泡包括外泌体、微囊泡和凋亡小体,外泌体是由大小为30~100 nm的膜囊泡构成。外泌体可以由各种动物的大部分细胞所分泌。在正常和病理情况下,外泌体参与众多的生物学过程。有证据[1]表明,外泌体内含有蛋白质、脂质、DNA和各种形式的RNA(例如miRNA、lncRNA等)。现就外泌体ncRNA在肝纤维化中的作用进行深入探析,以期为肝纤维化治疗药物寻找潜在新靶点。
1 外泌体lncRNA与肝纤维化
1.1 外泌体MALAT1 MALAT1基因位于人染色体11q13和小鼠染色体19qA内。MALAT1与许多蛋白质编码基因(如β-肌动蛋白,GAPDH)相当甚至具有更高的表达水平。MALAT1基因在人类中约为7 kb。先前有研究[2]发现,MALAT1可与miR-101b竞争来调节rac1的表达,影响HSC的增殖、细胞周期和激活,从而增加ECM的沉积。此外,MALAT1也可能通过介导SIRT1的下调,来活化肝星状细胞系LX-2细胞,导致肝纤维化的形成[3]。而Dai等[4]研究发现,使用砷酸盐可诱导肝细胞系L-02细胞MALAT1的过表达,并且MALAT1可以通过外泌体转运到LX-2细胞中,并认为MALAT1可通过miR-26b调节Ⅰ型胶原蛋白A2促进LX-2细胞的激活。由此可见,MALAT1可通过多种方式导致肝纤维化的形成。因此,外泌体中MALAT1可作为肝纤维化逆转的一个潜在靶点。
1.2 外泌体H19 H19是一种lncRNA,受H19和IGF2之间的位点甲基化的基因组印迹调控,越来越多的研究表明,H19具有许多不同的生物学功能。包括参与细胞增殖和分化,以及其在癌症中作为癌基因角色和肿瘤抑制剂等。H19主要在胆管细胞中表达,其能通过外泌体转移至肝细胞中显著下调小分子异二聚体伴侣的表达、调节S1PR2/SphK2和LET-7/HMGA2介导的途径、增加G1/S细胞周期转变促进HSC的增殖和激活等途径进而在调节胆管细胞增殖和促进肝纤维化方面发挥重要作用[5]。此外H19还可以作为miRNAs的分子海绵(如let-7 miRNAs),在胆汁性肝硬化的发展过程中发挥作用。
1.3 外泌体GAS5 GAS5 RNA基因位于细胞遗传学带1q25.1,GAS5基因有31个转录本,其中20个为保守的内含子,而11个为lncRNA。GAS5可以结合DNA结合结构域,从而使其无法调节靶基因的转录。GAS5可通过充当miR-222的竞争内源RNA(ceRNA)而增加p27蛋白的水平,从而抑制HSC的活化和增殖[6]。GAS5可作为miR-23a的分子海绵,从而竞争性地降低miR-23a的表达水平。miR-23a与PTEN的相互作用以及PTEN的降解进一步影响了下游信号通路PI3K/Akt/mTOR/ Snail,导致E-钙黏蛋白表达水平降低和α-平滑肌肌动蛋白(α-SMA)、Ⅰ型胶原蛋白的表达水平增加,从而导致肝纤维化的发生与发展[7]。据报道[8],外泌体GAS5的上调参与细胞凋亡过程,另外,有研究[9]表明,GAS5可通过外泌体途径调节巨噬细胞和内皮细胞的凋亡,来发挥抗动脉粥样硬化的作用。作为细胞外囊泡,外泌体在细胞间通讯中至关重要,并且可能是GAS5的关键载体[10],因此,推测GAS5亦可通过外泌体途径影响上述通路发挥抗纤维化作用。
1.4 外泌体MEG3 MEG3基因位于人染色体14q32.3区域的印迹DLK1-MEG3基因座上。MEG3在正常组织中表达,但在许多人类肿瘤和肿瘤衍生细胞系中丢失或减少。近些年已有研究[11-13]表明,含有MEG3的外泌体在肿瘤积液、宫颈阴道灌洗液、尿液中检测到,并在宫颈癌、高度浆液性癌和Hunner型间质性膀胱炎中起到作用。而关于肝纤维化的外泌体MEG3暂未有相关报道。有报道[14]指出,MEG3的过表达可激活p53并介导细胞色素c的释放,随后导致TGFβ1处理的LX-2细胞发生caspase-3依赖性的凋亡。这些发现表明,MEG3可能在HSC活化和肝纤维化进展中起重要作用,并作为肝纤维化的新型潜在治疗靶标。此外,MEG3亦可通过SMO蛋白和miR-212抑制Hh信号通路介导的肝纤维化上皮-间充质转化过程[15]。由此,预测MEG3可通过外泌体途径发挥抗肝纤维化作用,但还需要进一步研究。
1.5 外泌体PVT1 PVT1是一类lncRNA,人PVT1基因位于8q24,这是公认的与癌症相关的区域。PVT1已被证明与肝纤维化相关。有研究[16]表明,通过PVT1-miR-152-ATG14信号通路的自噬诱导有助于缺氧条件下HSC的激活。目前还未有外泌体PVT1在肝纤维化中的相关研究。但是,Meng等[17]报道外泌体介导的PVT1通过miR-93-5p调节的HMGB1/TLR4/NF-κB通路来调节脂多糖(LPS)诱导的骨关节炎进展。Wu等[18]通过实验发现,M2巨噬细胞来源的外泌体携带的lncRNA PVT1共同作用于miR-21-5p,以上调SOCS5并使JAKs/STAT3途径失活,从而减少炎症并保护自身免疫性脑脊髓炎小鼠。由此,将外泌体PVT1扩展在肝纤维化中,预测亦可发挥相应作用。
1.6 外泌体P21 P21基因位于人类染色体6p21.2上,位于细胞周期调控基因p21/Cdkn1a的上游,首次被描述为小鼠胚胎成纤维细胞p53依赖性凋亡的诱导剂。P21有两种亚型,它们都包含一个外显子和Alu反向重复序列。据报道[19],P21在HBV感染者血清中的水平低于健康对照者,且血清P21水平与感染者的肝纤维化阶段呈负相关。Zheng等[20]研究发现,P21的过度表达在体外抑制HSC的激活。慢病毒介导的P21转移至小鼠体内可降低肝纤维化的严重程度。Tu等[21]发现P21作为TGFβ信号的下游效应,通过与miR-30相互作用来增强TGFβ信号并介导其促进肝纤维化的作用。有报道[22],在非小细胞肺癌患者引流静脉血液及前列腺癌尿液中检测到含有P21的外泌体。肝纤维化有关外泌体P21尚未有文献报道,有待进一步研究。
2 外泌体短链非编码RNA与肝纤维化
2.1 外泌体miRNA miRNA是一种由22个左右的核苷酸构成的短链非编码RNA。通过与靶mRNA的3′-非翻译区结合参与转录后基因调控,影响多种细胞过程,包括细胞分化、能量代谢和代谢应激。根据近些年报道,与肝纤维化相关的miRNA有近百种,如能促进肝纤维化的有miR-200c[23]、miR-942[24]、miR-302c[25]、miR-182[26]、miR-195[27]等;具有抗肝纤维化作用的包括miR-30/miR-193[28]、miR-30[29]及miR-326[30]。有趣的是,一些研究发现miR-21[31-32]和miR-145[33-34]具有相反作用,既有促进肝纤维化又具有抗纤维化作用,因此具体的机制有待进一步研究。在外泌体miRNA相关研究[35]中发现,肝硬化肝癌患者血清和腹水中外泌体miR-182、miR-301a和miR-373的表达水平显著增加。有研究[36]表明,miR-122和miR-214可通过外泌体介导抑制HSC的活化,从而达到抗纤维化的作用。此外,miR-181-5p则可通过外泌体介导抑制STAT3/Bcl-2-Beclin 1途径来增加自噬并减少TGFβ1诱导的肝纤维化[37]。又有报道[38],miR-199a-5p可通过外泌体作用于结缔组织生长因子(CCN2),导致CCN2及其下游靶标α-SMA和Ⅰ型胶原蛋白α1的表达降低,从而发挥抗纤维化的作用。miR-223通过外泌体途径调控NLRP3和caspase-1发挥保肝作用[39],而miR-103-3p可通过外泌体介导靶向Krüppel样因子4(KLF4)促进HSC的增殖和激活[40]。外泌体miRNA与肝纤维化的关系总结详见表1。
2.2 外泌体snoRNA snoRNA是一类保守的核RNA家族,在核糖体亚单位成熟过程中参与小核RNA(snRNA)或核糖体RNA(rRNA)的修饰或加工。snoRNA越来越多地被证实参与调控新类型的转录后过程,例如RNA乙酰化、剪接模式的调节、mRNA丰度的控制和翻译效率,或者它们本身被加工成更短的稳定的RNA种类。先前已证实,在肝硬化患者体内snoRNA包括SNORD115-31、SNORD37、SNORD121B被下调。一项研究[41]表明,LPS可以在小鼠模型、人类受试者及巨噬细胞培养基中刺激snoRNA U32a(SNORD32a)、U33(SNORD33)、U34(SNORD34)和U35a(SNORD35a)的分泌,分泌的snoRNA和外泌体共同体被受体细胞吸收。在鼠模型中,证明外泌体snoRNA通过循环并在远端的组织中发挥功能。然而关于snoRNA在肝纤维化中的研究较少。有研究[42]表明,通过抑制SNHG7(一种snoRNA)可以抑制肝纤维化。进一步实验表明,可能是SNHG7充当ceRNA,通过与miR-29b结合来影响DNMT3A(miR-29b的下游靶基因)的表达,从而影响HSC的活化、自噬和增殖。
2.3 外泌体piRNA piRNA是一类小RNA,长度为24~31个核苷酸。它们与PIWI蛋白缔合,后者构成Argonaute家族的种系特异性亚群,形成称为piRNA诱导的沉默复合物的效应物复合物,该复合物通过转录或转录后机制抑制转座子并维持种系基因组完整性。除了在转座子沉默中起作用外,多种生物中的piRNA还可以调控细胞基因。越来越多的证据表明,piRNA在癌症发展中同时具有致癌作用和抑癌作用。但piRNA在肝纤维化中的作用研究较少。有研究[43]表明,激活的HSC中piR-823显著上调,进一步实验表明,活化的HSC可促进piR-823的合成,另一方面piR-823上调又可以促进HSC的活化。其机制可能为piR-823与EIF3B(真核起始因子3B)结合,并且该复合物进一步募集TGFβ1的mRNA并产生TGFβ1,TGFβ1激活静止的HSC所致。先前已有相关报道[44],在血清、脑脊髓液、尿液等体液中检测到含有piRNA的外泌体。因此,在肝纤维化中的外泌体piRNA需要进一步研究。
2.4 外泌体siRNA siRNA是一种由21~23个碱基对构成的双链RNA,包括两个突出核苷酸的羟基化3′末端。与靶mRNA具有序列互补性的siRNA反义链可诱导序列特异性基因表达沉默,这被称为RNA干扰(RNAi)。RNAi首先在秀丽隐杆线虫中被发现,后来在植物、动物和人类细胞中发现。越来越多关于siRNA的研究,其中也涉及到治疗肝纤维化,Toriyabe等[45]研究发现siRNA可抑制HSCⅠ型胶原蛋白α1、TGFβ、α-SMA的表达。Zhang等[46]用siRNA可干扰CTGF、TIMP-1、procol-α1、PCⅢ在肝组织中的蛋白表达水平。Ge等[47]则发现,GRB2 siRNA消除了HMGB1诱导的HSC增殖以及上调Ⅰ型胶原蛋白α1和α-SMA的作用。虽然越来越多关于siRNA的应用研究,但siRNA如何到达靶细胞发挥作用从而限制了siRNA发展
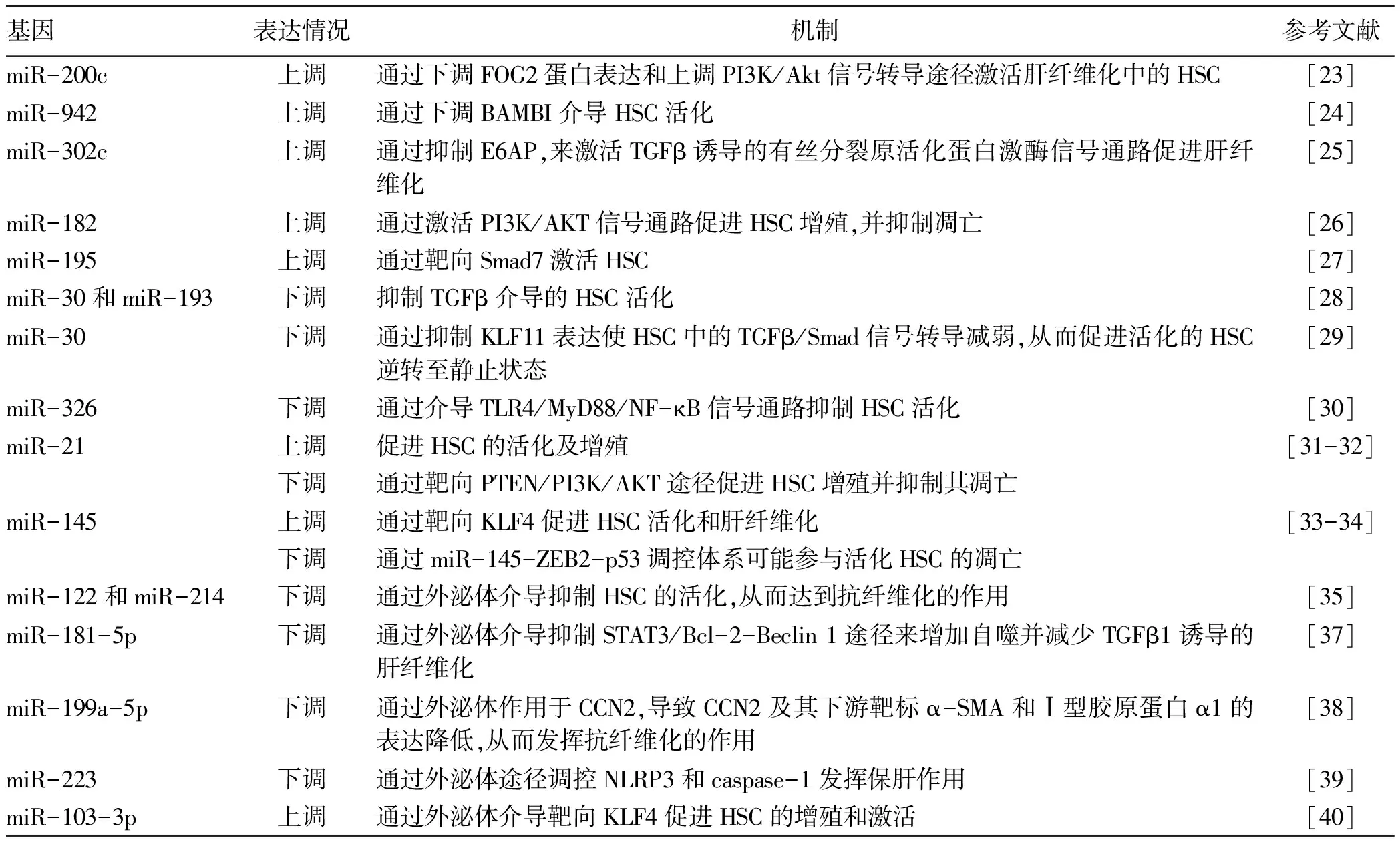
表1 外泌体miRNA与肝纤维化的关系
的机制仍不明确。目前除了常规局部及血清用药,还有化学修饰、脂质体、病毒载体、纳米颗粒等传递系统。近些年以外泌体为载体的研究越来越多应用于临床中,如神经系统疾病、眼部疾病、肝脏疾病、肾脏疾病等。Pan等[48]发现从人肝癌细胞释放的外泌体可以使siRNA在肝细胞之间穿梭,并抑制小鼠肝细胞中CD81的表达,这些结果使基于外泌体的siRNA在肝纤维化的治疗领域得到进一步扩展。
3 外泌体circRNA与肝纤维化
circRNA是一种共价闭合环状结构的RNA,没有5′端和3′端之分,主要由前体mRNA通过外显子的反向剪接形成。circRNA在肝病中有很多作用机制,包括microRNA海绵、蛋白质翻译、RNA支架、mRNA制动等。有研究[49]表明,通过分析CCl4小鼠肝纤维化模型肝组织中circRNA的表达谱,发现肝纤维化模型组和正常对照组之间有69个circRNAs的差异表达,其中14个上调,55个下调。进一步研究[50]发现,circ-PWWP2A可通过充当miR-203和miR-223的分子海绵促进HSC的活化和增殖。Ji等[51]研究发现,Hsa_circ_0070963作为miR-223-3p的分子海绵,通过调节miR-223-3p和LEMD3来抑制HSC的激活,从而抑制肝纤维化。Wang等[52]研究表明circMTO1通过调节miR-17-5p和Smad7抑制肝纤维化,而Jin等[53]发现circMTO1亦可通过miR-181b-5p介导的PTEN表达抑制HSC活化。Li等[54]发现hsa_circ_0004018/hsa-miR-660-3p/TEP1轴有助于HSC的增殖和激活。Zhu等[55]研究表明,脂肪来源的间充质干细胞分泌的外泌体传递mmu_circ_0000623,通过激活自噬防止肝纤维化。外泌体circRNA研究处于起步阶段,但其在寻找治疗肝纤维化药物及新靶点的道路上具有巨大的潜力。
4 小结与展望
部分研究已经证实外泌体中含有多种可调节肝纤维化的ncRNA,迄今为止,已经发现ncRNA可通过多种途径调节肝纤维化,包括TGFβ/Smad、Wnt/β-catenin、S1PR2/SphK2和LET-7/HMGA2、AKT/mTOR/p27、PTEN/PI3K/Akt/mTOR/Snail、NF-κB、Notch、Hh、SIRT/P53、STAT3/Bcl-2-Beclin 1和MAPK等信号传导途径。并且ncRNA调节肝纤维化的中心环节是HSC。此外,ncRNA不同种类之间,也会产生相互作用,比如前面提到的circRNA充当miRNA的分子海绵,lncRNA作为miRNA的ceRNA以及siRNA对于其他ncRNA的干扰作用等。目前对于ncRNA之间的关联性研究相对较少,因此,进一步研究不同类型的ncRNA之间的串扰和表观遗传网络的复杂级联,可能会突出关于肝纤维化进展的新发现,最终为肝纤维化的治疗提供一些新的策略。然而外泌体和ncRNA及其之间的调控机制还需要进一步研究。从而为寻找肝纤维化治疗药物及寻找潜在靶点提供帮助。肝纤维化是一个可逆转的病理过程,因此肝纤维化的早期干预在肝纤维化的治疗过程中显得尤为重要。但由于肝纤维化的发病机制复杂,调控的通路涉及众多,目前西药没有特效药物,且调控单一靶点的药物很难在临床上发挥良好的疗效,因此具有复合成分,多靶点特点的中药显示了独特的优势。
利益冲突声明:所有作者均声明不存在利益冲突。
作者贡献声明:钱南南负责课题设计,资料分析,撰写论文;唐露露、魏涛华负责课题设计,修改论文;杨悦、郝文杰参与收集数据;杨文明负责拟定写作思路,指导撰写文章并最后定稿。
