云南高黎贡山保山茜天然更新的分布特征1)
2021-11-03赵欣凤李贵祥柴勇和丽萍邵金平
赵欣凤 李贵祥 柴勇 和丽萍 邵金平
(云南省林业和草原科学院,昆明,650201)
保山茜(Fosbergiashweliensis)为茜草科(Rubiaceae)大果茜属(Fosbergia)常绿小乔木[1-2],原名为瑞丽茜树(Aidiashweliensis)[3]。保山茜为云南特有,高达10 m,花期为2—7月,当年生种子到翌年成熟,未受到干扰的保山茜种球不脱落。保山茜主要分布于云南高黎贡山一带,生长于海拔1 800~2 000 m[3]。云南省极小种群物种拯救保护规划纲要(2010—2020年)将其列为极小种群物种。
森林天然更新是森林资源再生产的一个重要的生态学过程,也是森林得以繁衍、持续不衰和森林生态系统稳定的基础[4-5]。森林天然更新主要受到环境因子与林分因子的影响[6],林下光照是影响树种更新及演替的重要因素,林冠层郁闭度是决定幼苗生长发育的关键[7]。郭连金[8]认为香果树幼苗受乔木盖度、灌木盖度、光照强度以及大气的温湿度影响较大;韩文娟等[9]认为凋落物是影响种子萌发的主要因素;王晗生[10]认为水分因子对人工林天然更新起着重要作用。许宁等[11]研究发现林窗或林缘更适合于水青树的自然更新;杨清培等[12]认为群落结构与物种组成是影响南方铁杉更新能力的主要因素;李荣等[13]认为近自然经营间伐措施能够促进辽东栎次生林的林木更新;影响林分天然更新的因子众多,森林类型或树种不同,影响林分天然更新的关键因子也不同[6,14],对于相同的林分其立地的差异也会导致其更新不一致[15]。
对保山茜的研究主要集中在该物种的确认[1]、染色体核型[2]、快速繁殖和离体保存[16]、物种名录[17]、微卫星位点标记[18]、干燥耐受力和发芽潜力[19]以及叶绿体基因组系列[20]等方面,有关保山茜天然更新的研究未见报道,本文通过对保山茜更新能力及空间格局、更新苗分布状况、距母树不同距离、不同方位的天然更新状况的调查分析,从而为保山茜种群保护和扩繁提供技术支撑。
1 研究区概况
研究区位于以云南高黎贡山南段(地处云南省保山市隆阳区的赧亢村、腾冲市的小地方村和龙陵县的小黑山交汇地段)保山茜较为集中的分布区域。地势东高西低,为我国西南部亚热带高原季风气候,海拔2 000~2 300 m。全年盛行西南风,干湿季分明,日气温温差大,兼有大陆性和海洋性气候特征。年均气温15 ℃,最冷月为1月,气温为7 ℃左右,年降水量1 200 mm左右。11月下旬至翌年4月为干季,日照充足,降水少,相对湿度50%~60%,5—10月为湿季,降水量占全年的87%,相对湿度比干季高20%左右。研究区土壤为黄棕壤、棕壤,主要植被类型为中山湿性常绿阔叶林。
2 研究方法
2.1 调查方法
在保山茜分布的区域内,通过充分踏查,随机选择已结实的10株保山茜作为母树,所选母树基本情况见表1,分别记录母树的位置、海拔、郁闭度、胸径、树高和冠幅等。
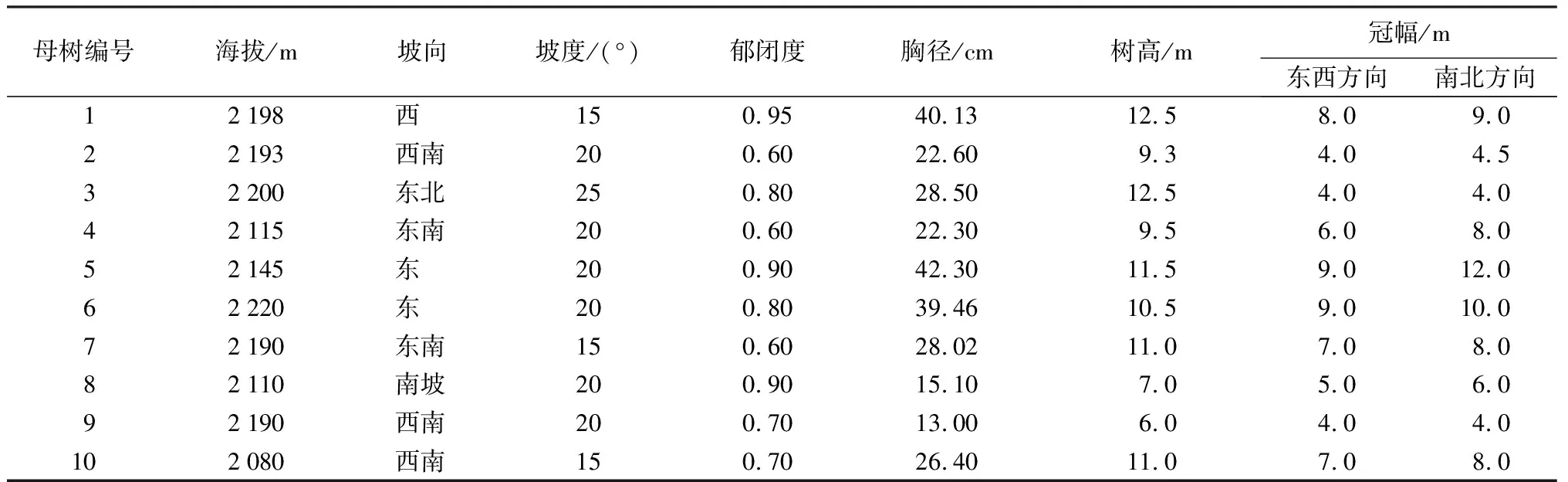
表1 保山茜天然更新调查母树基本情况
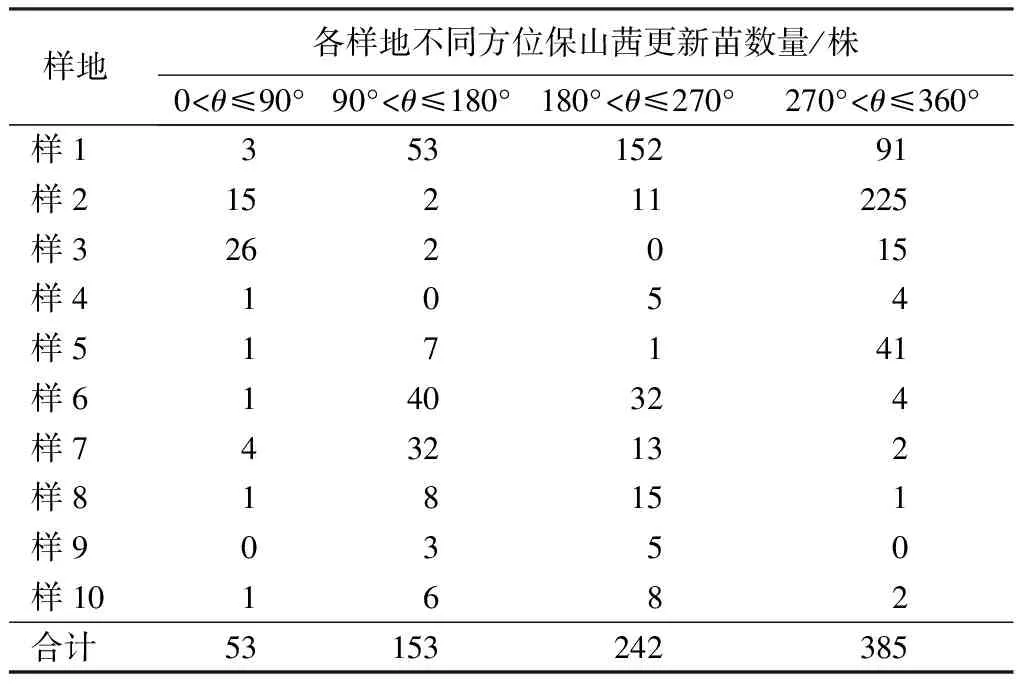
以母树为中心向外辐射,按半径(d)0 密度计算公式:D=x/s。式中x为样圆或样带内保山茜的个体数;s为样圆或样带面积;D为密度。 种群分布格局采用方差与平均数的比率确定,即S2/m[21-22],m=(∑fx)/n;S2=∑(x-m)2/(n-1)。式中x为样方中某个种的个体数;m为各样方中某个种个体平均数;n为取样个体数;S2为分散度(方差)。 年龄结构是种群动态的重要特征[23],一般采用高度和径级大小结构代替年龄结构的方法来分析其结构特征[8,11,23-26],但大小结构没有统一的划分标准,大多按植物特征和实际需要进行划分。因此,保山茜苗木分级按天然更新的实际情况并参照袁士云等[25]的划分方法,将保山茜天然更新苗高(H)划为Ⅰ级苗(H≥50 cm)、Ⅱ级苗(30 cm≤H<50 cm)和Ⅲ级苗(H<30 cm),母树作为大树不计入更新分析。 采用Microsoft Excel 97-2003对保山茜更新苗木进行统计整理,并在不同方位角和不同距离进行方差分析。 由表2可知,通过对研究区保山茜10株母树更新调查统计,保山茜的更新主要在以母树为中心的10 m半径范围内,在314 m2的样圆内更新株数为8~299株,10个样圆内共有833株;保山茜种群密度为254~9 522株/hm2,平均密度为2 652株/hm2。 表2 保山茜在各样圆中的更新数量 组成种群的个体在其生活空间中的位置状态或布局分为均匀型、随机型和集聚型[22],通过对保山茜种群内分布型的指标方差和平均数的比率(S2/m)计算,得到S2/m=130.69,即S2/m显著大于1,说明保山茜在调查样圆内为集聚型分布,表明保山茜种群个体分布不均匀,幼树的分布为集聚型。 由表3可知,在10个样圆分布的843株保山茜植株总数中,母树10株,更新苗833株,其中Ⅲ级苗有742株,占更新株数的89.08%;Ⅱ级苗有62株,占更新株数的7.44%;Ⅰ级苗29株,占更新株数的3.48%。从Ⅰ级苗、Ⅱ级苗和Ⅲ级苗更新数量来看,随着苗木龄级增长,大量的更新苗受种内竞争与环境因素的影响,数量急剧减少,而且所调查样圆内保山茜更新的苗木树高均小于1.3 m,说明保山茜虽然更新幼苗较多,但在母树下没有苗木长成小树或大树。 表3 保山茜不同龄级(苗高)的更新状况 通过对保山茜不同龄级的更新分布株数进行方差分析,得到F=4.597 822,P=0.019 123<0.05,表明不同龄级的保山茜更新分布株数差异显著。 由表4可知,保山茜在方位角0<θ≤90°、90°<θ≤180°、180°<θ≤270°、270°<θ≤360°的更新株数分别为53、153、242、385株,所占比例分别为6.36%、18.37%、29.05%和46.22%。从不同方位角的更新状况来看,不同方位角范围内,保山茜更新具有一定差异,但通过对保山茜不同方位角更新分布株数进行方差分析,得到F=1.037 789,P=0.387 52>0.05,表明不同方位角的保山茜更新分布株数差异不显著。 由表5可知,保山茜距离母树不同范围的更新情况,在0 表4 保山茜不同方位角上的天然更新状况 表5 保山茜不同距离范围内的天然更新状况 保山茜为云南特有[27],其分布较少,刀志林等[18]研究认为保山茜野外分布仅在100 km2的范围内,数量不超过200株的成年单株,处于濒临灭绝状态,云南省将其列为极小种群物种。因此,其天然更新和解濒技术就更加受到关注。在更新过程中,任何一种因素都可能改变整个群落的动态[28],使一些种群自然结实率低,难以获得种子,一些种群不能结实[29],从而影响种群的更新。通过项目组多年的观察,保山茜天然更新受自身特点的影响较大,保山茜成年母树大多能够结实,结实率在90%以上,种子聚集在结实种球内,结实种球不易脱落,在受到风、鼠等自然或人为干扰才会脱落。保山茜种子在野外的发芽情况较好,但由于保山茜种子集中在各个种球内,在各个种球内的种子呈丛壮发芽,范围基本就是种球的投影面积,种子发芽较为集中,不易扩散。从保山茜的更新研究来看,保山茜更新主要集中在以母树为中心的10 m半径范围内,更新密度在254~9 522株/hm2,种群内分布格局(S2/m)为130.69,显著大于1,说明保山茜天然更新在调查样圆内为集聚型分布,种群个体分布不均匀,与种子较为集中的特点是一致的,导致了保山茜种群扩大较为困难。 天然更新直接影响植物种群的数量和分布格局[30],保山茜在10个样圆中天然更新的株数是843株,Ⅲ级苗有742株,占更新株数的89.08%;Ⅱ级苗有62株,占更新株数的7.44%;Ⅰ级苗有29株,占更新株数的3.48%。保山茜在不同龄级的更新分布株数差异显著(P<0.05),表明保山茜虽然更新幼苗较多,但随着苗木龄级增长,数量急剧减少,能长成小树或大树的苗木不多。在方位角0<θ≤90°、90°<θ≤180°、180°<θ≤270°、270°<θ≤360°的更新株数所占比例分别为6.36%、18.37%、29.05%和46.22%,不同方位角保山茜更新分布株数差异不显著(P>0.05)。同种个体密度较高或者距离母树较近的地方,物种的种子、幼苗以及幼树的存活率较低[28]。从离母树不同距离范围的保山茜更新情况可看出,离母树2 m2.2 种群数量及分布格局
2.3 年龄结构划分
3 结果与分析
3.1 保山茜的天然更新能力及空间格局

3.2 保山茜更新苗年龄分布特征

3.3 保山茜在不同方位角上的更新状况
3.4 保山茜在不同距离的更新状况

4 结论与讨论
