降香黄檀内生细菌多样性研究
2021-11-02蓝柏成刘君昂周国英
蓝柏成,刘君昂,肖 柏,周国英
(中南林业科技大学 a.南方人工林病虫害防控国家林业局重点实验室;b.森林有害生物防控湖南省重点实验室;c.经济林培育与保护教育部重点实验室,湖南 长沙 410004)
降香黄檀Dalbergia odorifera是我国珍贵的濒危红木树种[1],因其心材质地坚硬、花纹美观、文理细密、不易开裂和变形、耐腐蚀等优点,通常被用来制作高档家具[2]。降香黄檀木材富含芳香油,尤其是其心材部位,降香油就是通过蒸馏技术从心材中提取的芳香油物质,该物质气味清香持久,不容易挥发,是良好的定香剂。同时在医学上具有阻止血液凝固和扩张心脏冠脉等作用是一种高级镇痛药材[3-4]。近年来,国家在我国广东、云南、广西等降香黄檀生长适宜地大力发展降香黄檀人工林的培育工作[5-7]。而降香黄檀在自然条件下的生长周期长,木材成型时间晚[8-9],同时降香黄檀纯林极易遭受风害、病害、虫害等自然灾害。根据董文统等[10]、刘成锋等[11]和周斌[12]对海南岛降香黄檀病害的全面调查结果显示,降香黄檀的常见病害包括叶枯病、细菌性穿孔病、炭疽病、黑痣病等。这些病害严重影响了降香黄檀的生长发育尤其降香黄檀的成材。因此对于促进降香黄檀的健康生长及抗病能力的提高等科学问题的解决迫在眉睫。
已有研究表明降香黄檀内生菌对降香黄檀的生长发育有重要的影响,降香黄檀心材的形成与降香黄檀的内生菌紧密相关[13]。2014年,贾瑞丰[14]研究发现4 种真菌都对降香黄檀心材的形成存在促进作用,其中以多毛孢Pestalotiopsis palmarum对心材的促进作用效果最为明显。2020年,刘昌霖等[15]对降香黄檀内生真菌的多样性及其与内环境因子的关系进行研究,初步探讨了内生真菌对降香黄檀生长的影响。但是都是关于降香黄檀内生真菌的研究,尚未见降香黄檀内生细菌的报道。
从植物的内生细菌中筛选出对于病原菌具有拮抗作用的细菌是十分重要的新型微生物资源,这些拮抗细菌往往更适应植物的内环境,能更好的定殖在植物体内[16]。对于降香黄檀病害的生物防治,内生拮抗细菌也有相关实例被报道。研究表明,降香黄檀叶部内生细菌中的一些芽孢杆菌属Bacillus细菌能有效抑制炭疽病病原菌的生长发育[17]。由此可见,降香黄檀病害相关的内生拮抗菌资源还有待开发。
因此,为全面了解内生细菌在降香黄檀中的群落结构及群落多样性,尤其是心材和边材的之间的差异,本研究使用HiSeq 高通量测序的方法,测序分析了降香黄檀木材样本中的内生细菌。同时测定降香黄檀心材边材的理化性质。结合两者的结果,综合分析两者之间的相关性。为后续人工促进降香黄檀健康生长及降香黄檀病害内生拮抗细菌的筛选提供科学的理论依据。
1 材料与方法
1.1 研究材料
采样地概况:地处海南省琼海市市郊。土壤肥力较高,土层不深厚,土层中夹有母岩半风化物,为砂红壤。主要树种包括降香黄檀Dalbergia odorifera、小叶榄仁Terminalia neotaliala、天料木Homalium hainanense、沉香Aquilaria sinensis。
采样方法:选取树龄均为15 a,胸径大小相近的降香黄檀。选取降香黄檀距离地面15 cm 处的茎干部分取。在无菌条件下,将样品处理成厚度0.5 cm 薄片。为了释放降香黄檀木材样本中的内生细菌,配置0.9% NaCl 溶液至于锥形瓶中,然后加入5 g 样本,最后将锥形瓶至于温度控制在25℃的摇床上,摇瓶培养2 h。培养后将的得到的样本,用0.22 μm 滤膜真空抽滤备用[15]。心材样本以下均使用代号HW,边材样本均使用代号SW。
1.2 内生细菌的高通量测序
使用总DNA 提取试剂盒(北京毕特博)提取降香黄檀内生细菌DNA。PCR 扩增实验方法参考刘昌霖等[15]研究论文,正向引物:341F(5′-CCTAYGGGRBGCASCAG-3′);反向引物:806R(5′-GGACTACHVGGGTWTCTAAT-3′)。后续的文库构建及高通量测序实验由mega genomics(美因健康)完成。
1.3 理化性质测定
含水率、pH 值、灰分含量、热水抽提物含量和矿质元素含量的测定,具体方法及步骤参考文献[15]。
1.4 数据处理
下机后,获得原始测序数据。对于原始数据的分析以及初始结果的获得,由Mega genomics(美因健康)完成。
初始结果的相对丰度制图使用R 软件完成;方差、相关性分析和冗余分析分别使用SPSS 19.0软件和Canoco 4.5 软件完成。
本研究OUT 聚类分析选用Unite 数据库,采用RDP classifier 贝叶斯算法对97%相似水平的OTU 代表序列进行分类学和生物信息统计分析[15]。
2 结果与分析
2.1 内生细菌测序数据质控结果
平均每个样品产生序列:心材115 985 条,边材185 600 条。经过过滤质控后,有效序列的平均长度为447.5 bp,平均降香黄檀心材和边材得到有效序列为75 638 条和140 889 条。降香黄檀心材和边材总共获得碱基数为96 481 212 bp(表1)。

表1 测序结果统计Table 1 Statistics of sequencing results
心材和边材的内生细菌Observed OTUs 稀释结果看出(图1),降香黄檀心材和边材的稀释曲线末端都逐渐达到平坦,说明了对于本研究的样本,此次测序数据量的结果是合理的。
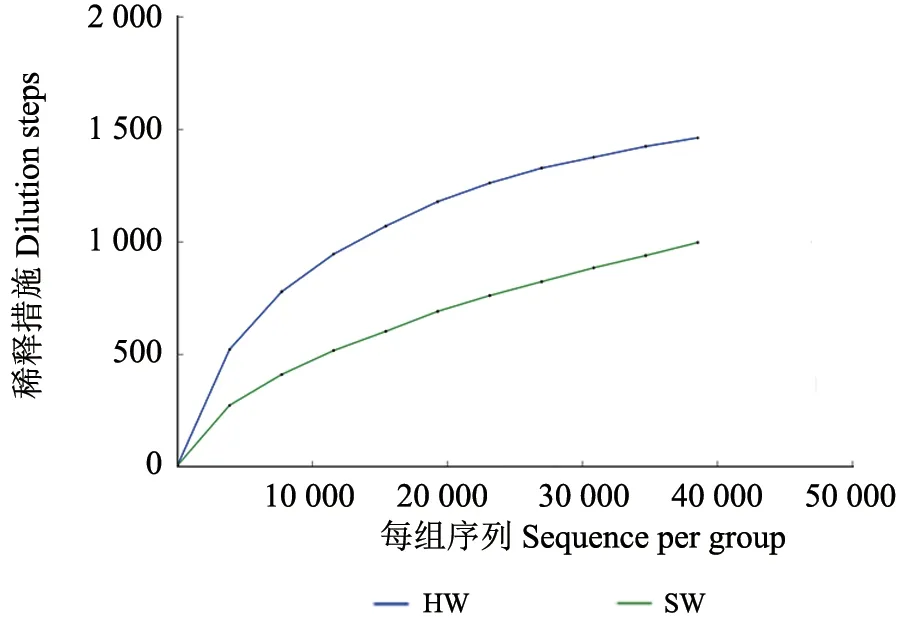
图1 Observed OTUs 稀释曲线Fig.1 Observed OTUs rarefaction curves
2.2 内生细菌OTU 聚类分析
为了更直观的展示降香黄檀的OUT 在心材和边材中的分布情况,根据OTU 的分析结果绘制Venn 图(图2)发现降香黄檀木材内生细菌总共有4 678 个OTU,其中心材和边材共有的OTU 数量有1 494 个,心材特有的OTU 有2 368 个,边材特有的OTU 有816 个。

图2 心材和边材OTU 的Venn 图Fig.2 Venn of HW and SW OTU
统计降香黄檀心材和边材注释到界、门、纲、目、科、属、种以及未鉴定的序列数目,统计结果如图3所示。属分类水平上,心材和边材的OTU注释数量最多,且心材和边材之间较大的差异。
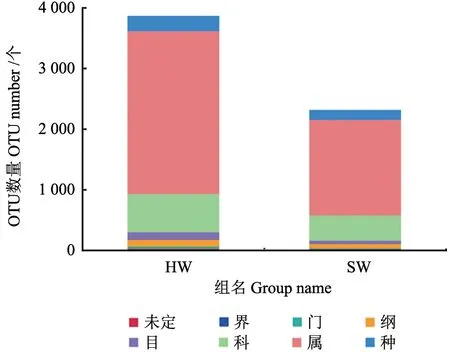
图3 心材和边材在各分类水平上的OTU 数目Fig.3 Number of OTU at each classification level for HW and SW
2.3 降香黄檀心材和边材内生细菌群落结构
在门分类水平上(图4),降香黄檀心材内生细菌群落中丰度最大的是变形菌门Proteobacteria(73%),其次是蓝细菌门Cyanobacteria(13%),除此之外主要的内生细菌群落还有厚壁菌门Firmicutes、酸杆菌门Acidobacteria 和拟杆菌门Bacteroidetes,这些菌群丰度均高于1%,低于10%。边材内生细菌优势菌群为变形菌门,占比96%,丰度远远高于其他菌群(丰度均低于1%)。由此可见,无论是在心材还是边材,变形菌门都是优势菌群。

图4 心材和边材内生细菌组成Fig.4 Endophytic bacteria of HW and SW
在属分类水平上,内生细菌在降香黄檀心材和边材的分布结果如图5,可以看出无论是降香黄檀的心材还是边材肠杆菌属Enterobacter都是明显优势菌属相对丰度都超过了10%。与心材不同的是,不动杆菌属Acinetobacter在边材中也占有很大的比例,它与肠杆菌属在边材的相对丰度达到了85%。在心材中也有其他菌属相对丰度超过了1%,相对丰度由大到小分别泛菌属Pantoea、伯克氏菌属Burkholderia、乳杆菌属Acidobacterium和柠檬酸杆菌属Citrobacter。而在边材内生细菌相对丰度超过1%的菌群依次为:肠杆菌属、不动杆菌属Acinetobacter、泛菌属、肠球菌属Enterococcus和克洛诺菌属Cronobacter。
肠杆菌属菌群在心材和边材中都属于优势菌群,且优势很大(图5),说明其在降香黄檀边材和心材两种环境中适应性都很强。不动杆菌属菌群在边材中为优势菌群,且心材和边材中丰度差异较大,更加适应边材环境。泛菌属菌群在心材和边材中相对丰度差异很小,且都是主要菌群。对于剩余的相对丰度低于1%的菌群,这些菌群中的大部分在边材中的丰度低于心材。

图5 心材和边材内生细菌组成Fig.5 Endophytic bacteria of HW and SW
2.4 内生细菌群落多样性分析
通过对比降香黄檀心材和边材的OTU 的数量,发现心材的数量(1 426)明显多于边材(808)。3 项指数即丰度指数、香侬指数和辛普森指数计算结果显示降香黄檀心材均大于边材。说明无论是内生细菌的物种数量还是多样性,心材都大于边材。

表2 多样性分析Table 2 Diversity index
2.5 降香黄檀心材和边材理化性质
降香黄檀心材含水率(39.87%)高于边材含水率(36.99%)。通过单因素方差分析结果得知,含水率在降香黄檀心材和边材之间并未存在显著性差异(P=0.05)(图6A)。
在常温常压下,降香黄檀木材pH 显酸性,与林场的土壤性质一致。心材和边材pH值分别为4.52和5.47。经过单因素方差分析表明,心材和边材pH 值存在极显著差异(P<0.01)(图6B)。
灰分(Ash)的高低可反映不同的植物的不同部位对矿质元素选择吸收积累的特点[18]。心材Ash 含量明显较边材Ash 含量低。心材的含量0.18%,边材的含量0.32%。单因素方差分析结果表明,降香黄檀心材和边材灰分含量存在显著性差异(P<0.05)。由此可以推测,边材对矿物元素(挥发性氧化物除外)的吸收和累积要比心材多(图6C)。
通常热水抽提物成分是糖类营养物质和可溶性无机盐等[19]。降香黄檀心材热水抽提物含量高于边材部分,分别为13.51%和11.69%,心材为边材的1.16 倍。单因素方差分析结果显示,降香黄檀心材、边材热水抽提物含量存在显著差异(P<0.05)。心材中抽提物沉积物越多,心材颜色越深。热水抽提物含量多少,反映了边材和心材所含的营养物质多少。心材中具有更多糖类营养物质(图6D)。
矿质元素在树体内的循环是从衰老边材向其他活体部分转移的[20]。研究发现心材、边材矿质元素的移动呈现一定的规律,部分移动的Mg 元素和不移动的Ca 元素在心材部分含量较边材高,可移动的K 元素比边材低[21]。降香黄檀心材钙元素含量较边材部位高,分别为5.76 和2.87 mg/g,单因素方差分析显示存在极显著差异(P<0.01)。镁元素含量心材和边材差距不大,心材镁元素含量(0.49 mg/g)比边材(0.43 mg/g)稍高一点,单因素方差显示不存在显著差异(P=0.113)。在钾元素含量方面,边材钾元素含量(1.69 mg/g)远高于心材钾元素含量(0.58 mg/g)。单因素方差表明,心材和边材钾元素含量存在极显著差异(P<0.01)(图6E)。
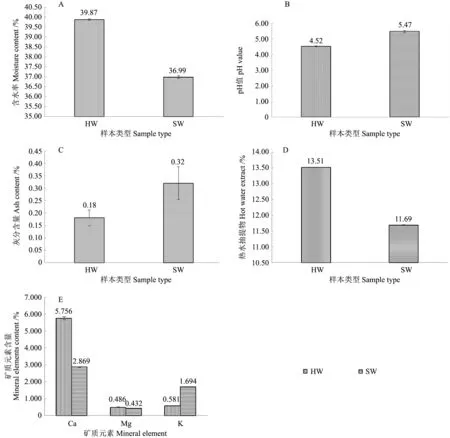
图6 心材和边材的理化性质Fig.6 Physical and chemical properties of HW and SW
2.6 内生细菌群落结构与理化性质的关系
冗余分析结果如图7所示,图中横向坐标轴解释64.9%的信息,纵向坐标轴解释35.1%的信息。从图7可以看出,降香黄檀边材中的优势菌群在图中分布相对集中,说明这些菌群的相关性较大。但是降香黄檀心材优势菌群在图中分布相对分散。说明这些菌群的相关性较小。pH 值和钾元素同时处在同一个方向,所以他们呈极显著正相关,且pH 值和钾元素刚好处在矿质元素钙和镁元素的反方向,呈极显著负相关。心材优势菌肠杆菌属受到木材因子影响大小依次为:HWE、Ash、Ca、K、pH、Mg、MC,与Ash、K 和pH 值正相关,与MC、Mg、Ca 和HWE 含量负相关。木材因子对边材优势菌不动杆菌属的影响大小与之正好相反,且同样与含水率、镁、钙和热水抽提物含量负相关。肠球菌属与pH 值和矿质元素钾极显著正相关,与矿质元素钙和镁极显著负相关。

图7 内生细菌菌群与理化性质的冗余分析Fig.7 RDA analysis of endophytic bacteria and physical and chemical properties
2.7 内生细菌群落多样性指数与理化性质的关系
对心材和边材样品内生细菌多样性指数和理化性质进行相关性分析(表3),结果表明,丰度指数、香侬指数与辛普森指数都互为正相关。pH值与丰度指数极显著正相关性,丰度指数、辛普森指数和香侬指数与含水率、灰分和矿质元素钾负相关,与热水抽提物、矿质元素钙和镁正相关。木材理化因子中含水率和pH 值对内生细菌群落多样性影响最大,具有显著相关性。其中内生细菌群落多样性受到木材因子pH 值的影响最大。

表3 理化性质与内生细菌多样性的相关性分析†Table 3 Correlation analysis of physical and chemical diversity and endophytic bacteria diversity
3 结论与讨论
3.1 讨 论
3.1.1 降香黄檀内生细菌和木材内环境因子的关系
影响降香黄檀内生细菌群落结构的木材内环境因子主要是灰分含量、热水提取物、pH 值、Ca含量和K 含量。内生细菌群落多样性受到木材因子pH 值的影响最大。pH 值在心材边材中呈显著差异,在含水率相近的情况下,木材pH 值对内生细菌的群落结构影响更大。降香黄檀心材和边材的热水抽提物含量分别为13.51%和11.69%,热水抽提物反映的是有机质等营养物质的含量。营养物质含量高的微生物种类和数量都多。这与心材内生细菌群落丰富度大于边材的结果相符合。灰分含量的结果表明,降香黄檀中边材比心材矿质元素累积的要多。其中Ca 含量和K 含量是引起内生细菌群落结构差异的主要矿质元素。心材矿质元素中钙元素的含量比边材高,研究表明钙含量的提高会降低变形菌门的丰度[22],而心材中变形菌门的相对丰度低于边材。解钾菌称硅酸盐细菌(Silicate bacteria),能有效分解并利用磷、钾等矿物元素;主要为厚壁菌门中的芽孢杆菌属Bacillus的细菌[23]。在心材内生细菌群落中,厚壁菌门Firmicutes丰度明显大于边材,因此的钾的含量会低于边材。
3.1.2 降香黄檀心材和边材的内生细菌多样性与差异性
研究表明,不同植物内生菌在不同植物体内的分布不同,在宿主不同器官的分布也存在着差异[24-25]。本研究显示,心材的多样性指数丰度指数和OUT 数量都高于边材,说明心材内生细菌的物种总数多于边材。辛普森指数和香侬指数,心材中内生细菌多样性要多于边材。这说明降香黄檀心材内生细菌的物种丰富度大于边材。也反映了降香黄檀内生细菌的分布受到了心材边材不同木材结构性质及理化性质的影响,使降香黄檀心材边材内生细菌的分布表现出了明显的差异。
分析降香黄檀内生细菌的群落结构组成上,心材边材也存在差异性。在门的分类水平上,变形菌门在心材和边材中都占绝对优势,同时在属水平上,肠杆菌属无论在心材还是边材也都是优势菌,这反映变形菌门的细菌能很好地适应心材边材的环境。有研究表明变形菌门是植物根际最丰富的优势菌群,主要原因是其生长速度快,同时能够充分利用植物根际分泌的各种物质[26],因此,这一观点在沙棘[27]、红豆杉[28]、醉马草[29]、人参[30]和甘草[31]等植物上得到了很好的证明。同时本研究显示变形菌门在心材和边材中都占绝对优势,同时变形菌门在边材的丰度大于心材,说明降香黄檀心材和边材的一些细菌可能来源于根际土壤。
蓝细菌门在心材的中的丰度达到了13%。但在边材中并不是优势菌群。蓝细菌门细菌分布广泛同时生存能力极强,还能与一些植物共生[32]。本研究没有对蓝细菌门进一步研究。后续应当对这一现象进一步深入研究,进一步明确降香黄檀心材和边材的内生细菌群落结构,阐述蓝细菌门与降香黄檀的关系。
不动杆菌属菌群在边材中为优势菌群,是多种植物的内生细菌,广泛存在于胡杨[33]、银砂槐[34]等植物体内,从各种植物组织中分离出的几种不动杆菌已被报道为潜在的生物防治剂。除了抑制病原菌的活性外,不动杆菌还表现出多种促进植物生长的特性,例如生长素的产生,磷酸盐的增溶,铁载体的产生和氨的产生[35-36]。与其他肠杆菌属不同的是,不动杆菌属心材和边材中丰度差异较大,更加适应边材环境。
3.1.3 降香黄檀内生细菌对降香黄檀生长的影响
内生细菌对于植物生长的促进作用主要分为直接和间接两种形式[37]。直接促进作用包括:固氮、溶磷、解钾和产生植物生长调节激素等。间接促进作用主要有对于植物病原菌的拮抗、产生ACC 脱氧酶、产生铁载体和诱导系统抗性等。如朱艳蕾[34]对银砂槐的6 株内生细菌进行促生抗逆作用的研究,研究发现6 株菌种均具备溶磷、固氮以及产生IAA 和ACC 脱氨酶等不同的促生能力。陆蓝翔等[38]对2 株樟树组织中内生细菌进行分离鉴定,其中有11 株菌对樟树病害病原菌具有拮抗效果,4 株菌具有解无机磷能力,2 株菌具有解有机磷能力,2 株菌具有产IAA 能力。田红雨[39]通过调控内生细菌的群落结构增强了桉树抗青枯病的能力,其中提高黄杆菌属Flavobacterium在桉树体内的比例可能是微生态调控的关键。降香黄檀内生细菌的对其的间接促生作用已有报道。2017年陈彧等[17]从降香黄檀叶片分离筛选了芽孢杆菌属Bacillus的炭疽病拮抗细菌Y23,而对于降香黄檀内生细菌的直接促生作用目前研究却很少。但是在降香黄檀的根际土壤中发现了大量的解钾和固氮的细菌,土壤是内生细菌的一个重要来源,这说明降香黄檀内生细菌资源还有待进一步的开发。
本研究由于降香黄檀树种的特殊性及国家政策等原因,考虑到其内生细菌群落结构及理化性质结果与实际情况的差距,可以选择对正常生长的降香黄檀直接取样进行研究。在后续的研究中可以对降香黄檀内生细菌的促生功能、内生拮抗细菌的筛选及应用等方面展开研究。为充分利用降香黄檀的内生细菌资源奠定理论基础同时也为降香黄檀病害的无公害防治提供科学依据。
3.2 结 论
对降香黄檀的心材边材理化性质进行分析,降香黄檀心材和边材各项理化性质均存在不同程度的差异。灰分含量、热水提取物、pH 值、Ca 含量和K 含量上存在明显差异。在对降香黄檀内生细菌的分布与木材理化因子的相关性分析中,含水率和pH 值对内生细菌群落结构的影响最大,其中降香黄檀内生细菌群落结构多样性受到木材因子pH 值的影响最大。
对降香黄檀心材边材的内生细菌HiSeq 高通量测序并优化后,结果表明,降香黄檀心材和边材内生细菌种群资源丰富多样,内生细菌群落丰富度心材大于边材。内生细菌多样性心材大于边材,说明降香黄檀心材和边材的内生细菌的入侵定殖也有差异性。
