乳酸菌作为生物保护菌的抑菌机理及其在食品中应用的研究进展
2021-10-31吕懿超李香澳王凯博孔保华
吕懿超,李香澳,王凯博,孔保华,陈 倩
(东北农业大学食品学院,黑龙江 哈尔滨 150030)
由食品中有害微生物造成的食品腐败和安全问题一直受到广泛关注。为了有效抑制有害微生物的生长,延长食品保质期,提高食品安全性,食品工业中常通过物理保鲜、添加化学防腐剂或使用新型包装方式等来抑制有害微生物[1]。近期研究发现,虽然化学防腐剂按照规定添加量单一添加时不会对人体造成损害,但防腐剂与增味剂和色素等多种添加剂混合添加时,会影响人体代谢,增加发生慢性病的风险[2]。另外,随着生活水平的提高,消费者希望食品行业减少塑料等包装材料的使用[3]。因此寻找天然、安全的保鲜方法成为近年来的研究热点。近年来“生物保护菌”的提出满足了消费者的这一需求。
生物保护菌是指可以添加到食品中的具有延长食品货架期和(或者)抑制有害微生物生长的活的微生物[4]。乳酸菌因可以产生具有抑菌活性的代谢产物(有机酸、细菌素和环肽等)[5],并可以通过微生物群体感应[6]和竞争作用[7]来抑制腐败菌生长,成为生物保护菌筛选的重点。2016年,明串珠菌4010(Leuconostoc carnosum4010)被加拿大政府批准作为生物保护菌添加到真空包装肉制品中[8]。随着乳酸菌作为生物保护菌的合法化,将乳酸菌作为生物保护菌应用在食品领域中成为研究热点。因此,近年来越来越多的研究人员将乳酸菌应用于不同食品的生物保护中。本文将主要从乳酸菌产生的抑菌物质、微生物群体感应以及竞争作用等方面介绍乳酸菌的抑菌原理,同时综述了近些年乳酸菌作为生物保护菌在乳制品、肉制品、水产品和果蔬产品中的应用,并对生物保护菌未来的研究方向进行了展望。
1 乳酸菌的抑菌机理
对于乳酸菌抑菌机理的研究,早期一直关注其代谢产生的抑菌物质,如有机酸、细菌素等,这些物质主要通过增加微生物膜通透性来抑制有害微生物的生长[9]。随着研究的进一步深入,近些年发现乳酸菌可以通过微生物群体感应作用有效抑制有害微生物的生长和生物膜的形成[6]。此外,研究还发现乳酸菌可以通过发酵产品产生抑菌蛋白或通过与有害微生物竞争[7]等方式来抑制有害微生物生长。
1.1 乳酸菌代谢产生有机酸类
乳酸菌代谢产生的有机酸,如乙酸、乳酸、碳原子数较多的脂肪酸和苯乳酸等可通过竞争作用(图1机制一)、增加细菌外膜通透性(图1机制二)、改变胞内渗透压(图1机制三),以及抑制大分子的合成(图1机制四)等方式来抑制有害微生物生长[10]。
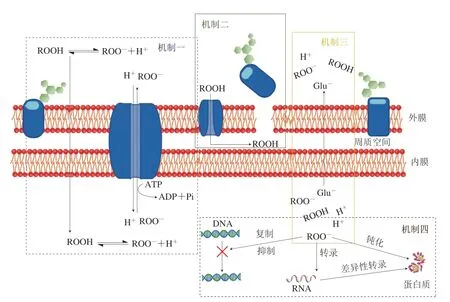
图1 乳酸菌代谢产生有机酸的抑菌机制[10]Fig. 1 Antimicrobial mechanism of organic acids produced by lactic acid bacteria[10]
1.1.1 竞争作用
1.1.1.1 能量竞争
乳酸菌代谢产生的有机酸属于弱酸,这些弱酸在细胞外液中以解离和未解离两种状态存在,而未解离的有机酸分子可以通过自由扩散进入到细胞内[11]。由于细胞内处于偏碱性环境,使得有机酸解离平衡向产生氢离子和酸根离子方向移动,进而产生两种离子。但是这两种离子不能通过自由扩散的方式扩散到细胞外[12],所以为了维持细胞内环境的稳定,细胞需要消耗ATP,通过主动运输的方式将氢离子转运到胞外。然而这是一个额外耗能过程,需要同细菌正常代谢所需能量产生竞争[13]。与此同时,有机酸可以消除ATP和电子传递的偶联作用,从而降低细菌的产能效率。乳酸菌通过这种能量竞争的方式可抑制微生物的生长。
1.1.1.2 营养竞争
除了对能量竞争外,有机酸还可以与微生物中营养物质发生作用,造成同微生物的营养竞争。Mani-López等[14]研究发现,乳酸在细胞外形成的乳酸盐可改变沙门氏菌周围水分活度,通过对水分子的竞争降低水分活度达到抑菌的目的。此外,一些有机酸可螯合金属离子,与微生物生长所需的金属离子形成螯合物,从而抑制微生物的生长[15]。
1.1.2 增加膜通透性
由于有机酸具有一定的脂溶性,可以作用于细胞膜,使得膜脂多糖和磷脂大量释放造成细胞膜稳定性降低[16]。同时有机酸还可以通过膜表面的孔道蛋白,进入细胞外膜和细胞内膜之间的周质,并导致脂多糖羧基和磷酸基团质子化,从而使得细胞外膜被破坏,细胞内物质泄漏[17]。参与竞争机制模式的氢离子在被转运到胞外的同时,钾离子被泵入到细胞内,造成细菌跨膜质子动力势能被破坏,胞内渗透压升高,从而破坏细菌细胞膜,导致内容物外泄,从而起到抑制微生物生长的作用[16]。
1.1.3 改变胞内渗透压
有机酸在胞内解离,以氢离子和酸根阴离子形式存在于细胞内。因此有机酸进入细胞质,不仅会导致胞内渗透压升高,还会导致阴阳离子浓度升高。为了恢复胞内渗透压,保持内外有机酸分子和阴阳离子浓度平衡,细胞需要释放阴阳离子。但是释放没有选择性,一些带有负电的细菌生长所需前体物质及辅助因子会同酸根离子一起被释放到胞外,导致微生物生长受阻[18]。Roe等[19]研究发现大肠埃希氏菌在乙酸作用下,胞内乙酸根浓度升高,为了降低胞内渗透压,大肠埃希氏菌释放谷氨酸根,最终导致大肠埃希氏菌生长速率减慢。
1.1.4 抑制大分子合成
有机酸可以通过影响微生物DNA的合成影响转录过程,进而影响合成相应蛋白质来抑制微生物生长。由于乳酸菌产生乳酸并在胞内聚集,原本糖酵解的中间产物丙酮酸由于反馈抑制不能继续转化成乳酸,而使得丙酮酸在细胞中积累。为了减少丙酮酸在细胞中的积累,细胞通过增强磷酸戊糖途径所需要酶的转录能力,提高酶的含量,减少丙酮酸的积累,同时降低了细胞产能,从而抑制细菌生长[20]。此外,有机酸可抑制胞内酶的活性,导致细胞内代谢活动受阻。Roe等[21]研究指出乙酸可以抑制大肠埃希氏菌K-12(Escherichia coliK-12)中高半胱氨酸转化为蛋氨酸时所需酶的活力,蛋氨酸的缺失影响了细菌的生长,同时高半胱氨酸由于不能及时转化,在细胞内不断积累,对细胞产生毒害作用。
1.2 细菌素类
细菌素是细菌在代谢过程中产生的多肽或蛋白质类抗菌物质,具有抗细菌和真菌的作用[22]。并且大部分细菌素具有膜活性,可通过连接、嵌入作用在细胞膜上形成孔道,使得胞内容物外泄,导致细胞死亡[23]。细菌素可分类4 类:I类是羊毛硫抗生素,II类是热稳定、无修饰的小分子肽,III类为热不稳定的大分子肽,IV类为蛋白质化合物[24]。
1.2.1 羊毛硫抗生素
I类细菌素由于结构不同可以被分为两类,分别为带正电且具有疏水基团,具有双亲性、螺旋状肽链的A型和分子质量较小、含有很多小型环状结构的B型[25]。乳酸菌产生的大多为A型羊毛硫抗生素。
A型羊毛硫抗生素对革兰氏阳性菌具有较好的抑制作用,但对革兰氏阴性菌、真菌和酵母基本没有效果[26]。通过体外模拟A型羊毛硫抗生素对磷脂双分子层的研究发现,细胞膜表面的孔道主要经过4 步形成(图2):1)细菌素通过静电相互作用与靶细菌的细胞膜相结合;2)细菌素氮端与细菌细胞膜结合;3)细菌素碳端跨过细菌细胞膜;4)在细菌细胞膜上形成孔道,造成细胞膜通透性增加[27]。
以典型的A型羊毛硫抗生素乳酸链球菌素(Nisin)为例:Nisin作用于细菌细胞膜时,其碳末端首先通过静电相互作用与带负电的磷脂结合。革兰氏阳性菌细胞膜含有大量的负电脂质(磷脂酰甘油和心磷脂),而革兰氏阴性菌和酵母菌的细胞膜通常是两性脂质,因此Nisin对革兰氏阳性菌更为有效。由于Nisin的氮端与表面蛋白共享一个同源的氮环,因此Nisin的氮端可以与细胞膜通过肽聚糖前体脂质II相互作用特异性结合,使得Nisin分子平行贴在细胞膜表面。此外,肽聚糖是细菌细胞壁的主要成分,但是由于肽聚糖前体物质与Nisin结合,导致脂质II不能正常合成肽聚糖,从而干扰细菌细胞壁的正常合成和细胞正常分裂。Nisin在细胞膜表面通过两种方式形成孔道,分别称为“桶板”模型(图2A)和“楔形”模型(图2B)。“桶板”模型指出Nisin疏水残基与脂质的酯酰基相互作用,其极性残基位于膜-水界面,由于大量聚集使得膜局部移位变薄,Nisin碳端以电势依赖方式插入双层膜疏水中心,最后由多个Nisin分子一起形成孔道。而另一种“楔形”模型中,质子能耗散,膜内外存在高跨膜电势,导致磷脂膜表面弯曲,膜平面方向改变产生楔形孔隙[28]。
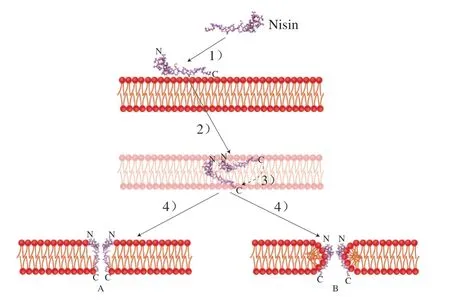
图2 Nisin抑菌机制[27]Fig. 2 Antimicrobial mechanism of nisin[27]
1.2.2 肽类细菌素
II类细菌素是一类热稳定、未修饰的小肽,分为4 类:IIa类(类乳酸片球菌素细菌素)是一类热稳定性小肽,是通过两个甘氨酸残基加工形成的前体合成的,对单核细胞增生李斯特菌有强烈的抑菌活性,氮端具有共有序列YGNGVX-C;IIb类(双肽、未修饰细菌素)是由两个不同的肽组成一个具有抗菌活性的络合物;IIc类(环状细菌素)和IId类(未修饰、线性、非类乳酸片球菌素细菌素)[24,29-30]。
植物乳杆菌TN635(Lactobacillus plantarumsp.TN635)产生的IIa类细菌素BacTN635可以通过碳端α-螺旋结构吸附于细胞膜上,同样通过在细胞膜形成孔道,使得细胞膜通透性增加,从而引起能量消耗,内容物外泄,导致细胞死亡(类似于图1机制二)[31]。而IIb类细菌素一般需要两个肽的相互作用,而IIb类细菌素也被分为两类:一类是Lactococcin G,当这类细菌素的两个肽分开时,就不具备抑菌活性,而两个肽相互作用时可以消耗质子势能,减少ATP的生成,同时抑制氨基酸的摄取;而另一类的代表则是由嗜热链球菌产生的由两个肽组成的Thermophilin13,其中一个是抗菌肽ThmA,而另一个肽是ThmB。这种嗜热链球菌素可以抑制李斯特菌,并且ThmB可作为增强因子将ThmA的活性提高约40 倍[32]。IIc和IId细菌素都可以在细菌细胞膜表面形成孔道,从而增加细胞膜的通透性,且都对革兰氏阳性菌敏感。格氏乳杆菌和肠球菌分别可以产生IIc细菌素Gassericin A、Enterocin AS-48,并且可以作用于膜表面ABC转运蛋白,从而形成孔道[33-34]。而唾液乳杆菌和乳酸乳球菌分别产生的IId细菌素Bactofencin A和LsbB则是通过作用于金属多肽酶来达到抑菌的目的[35-36]。
1.2.3 热敏感大分子蛋白
III类细菌素是一种热敏感大分子蛋白,分子质量一般大于30 kDa。这类细菌素也是通过在细菌细胞膜上形成孔道,从而导致细胞膜通透性增加,达到抑菌目的。但是与A型羊毛硫抗生素相比,III类细菌素有较好的广谱性,可以有效抑制革兰氏阴性菌和革兰氏阳性菌的生长。但这类细菌素由于研究信息较少,还没有具体作用受体的相关报告[37]。卷曲乳杆菌(Lb.crispatus)[38]、瑞士乳杆菌CNRZ450(Lb. helveticusCNRZ450)和瑞士乳杆菌481(Lb. helveticus481)[39-40]均可以产生III类细菌素Helveticin M和Helveticin J。Helveticin M可以破坏革兰氏阳性菌细胞壁和革兰氏阴性菌外膜的稳定性,因而对革兰氏阴性菌和阳性菌均有抑制作用。
1.2.4 大分子复合物
IV类细菌素是由碳水化合物、蛋白质和类脂基团组成的大分子复合物,可以有效抑制真菌和革兰氏阴性菌的生长[9]。赵娜等[41]发现瑞士乳杆菌AJT(Lb. helveticusAJT)可以产生对大肠埃希氏菌、金黄色葡萄球菌和青霉有抑制作用抗菌物质。排除酸性物质和过氧化氢的影响后,抗菌物质经蛋白酶处理,发现抗菌活性下降但依然具有抗菌活性,从而表明抗菌物质是由蛋白和非蛋白物质复合而成,属于IV类细菌素。同时,一些研究人员发现从埃及盐腌发酵鱼制品中分离出的粪肠球菌F4-9(Enterococcus faecalisF4-9)可以产生一种对蛋白酶敏感、pH值活性范围较宽、耐热性较好的细菌素,称为Enterocin F4-9。该细菌素作用范围较窄,对革兰氏阴性菌大肠埃希氏菌JM109(E. coliJM109)有抑制作用[42]。此外,乳酸乳球菌L9(Lactococcus lactisL9)也可以产生此类细菌素Lactococcin GJ-9[43]。目前发现的细菌素Pediocin SJ-1[44]、Leuconocin S和Lactocin 27也属于IV类细菌素[36]。
1.3 群体感应抑制剂
微生物群体感应是指微生物可以产生一类被称为自诱导物的信号分子,信号分子浓度与微生物密度成正比,当信号分子积累到一定阈值时,开始通过激活并结合其受体蛋白来启动相关基因的表达[45]。一些腐败菌和致病菌的腐败能力、遗传能力、毒力以及生物膜的形成都与群体感应有关[46]。并且研究发现导致乳制品、肉制品、水产品和蔬菜等变质的微生物的腐败能力都受到群体感应的调控[47-48]。因此抑制或阻断细菌群体感应可以有效抑制腐败菌或致病菌的生长和毒性,从而有效延长食品保质期,提高安全性。
群体感应抑制剂就是一类通过调控微生物群体感应系统来抑制腐败菌的生长或者降低其对环境耐受性的物质[48]。群体感应抑制剂一般通过阻断信号分子的合成(图3A)、降解群体感应信号分子(图3B)和降低或抑制信号分子受体蛋白的活性(图3C)来抑制群体感应的进行,从而对腐败菌或致病菌产生抑制作用。
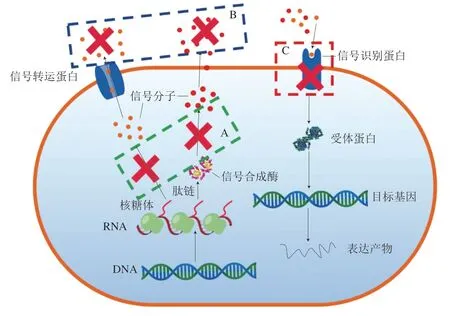
图3 群体感应抑菌机制[49]Fig. 3 Antimicrobial mechanism of quorum sensing[49]
1.3.1 细菌群体感应抑制剂
铜绿假单胞菌PAO1(Pseudomonas aeruginosaPAO1)毒力因子的产生和生物膜的形成依赖于细菌群体感应系统,植物乳杆菌产生的3-苯基乳酸可以作为细菌群体感应抑制剂抑制该菌的生长[49]。此外,水杨酸是一种乳酸菌代谢产物,可以通过抑制高丝氨酸内脂合成酶的活性,阻断信号分子的合成,抑制腐败菌的生长(图3A)[48]。除了抑制信号分子的合成可以有效抑制细菌群体感应,降解信号分子也可以抑制细菌群体感应。林洋等[50]发现乳酸菌培养液的粗提取物可以通过降解细菌群体感应信号分子N-酰基高丝氨酸内脂抑制嗜水气单胞菌属生物膜的形成(图3B),由于粗提物经蛋白酶处理失去降解作用,猜测粗提物是一种酶类。Park等[51]从泡菜中筛选出一株清酒乳杆菌NR28(Lb. sakeiNR28),这种乳酸菌虽然不能抑制大肠埃希氏菌的生长,但是可以通过抑制革兰氏阳性菌及革兰氏阴性菌共同利用的呋喃酮酰硼酸二酯类信号分子(autoinducer-2,AI-2)转导途径,对大肠埃希氏菌ATCC-43894(E. coliATCC-43894)菌株的致病性有一定的抑制作用(图3C)。此外,乳酸乳球菌LY3-1(L. lactisLY3-1)可以产生由10 个氨基酸(序列为:ALSLVKIHGR)组成的群体感应抑制剂,有效抑制哈维氏弧菌胞外多糖、生物膜和鞭毛的合成[48]。
1.3.2 真菌群体感应抑制剂
白念珠菌是一种常见的人类条件致病菌,通常以无治病作用的酵母相和具有治病作用的菌丝相两种形式存在。白念珠菌可以通过群体感应在这两种形式之间转换,随着数量的增长,白念珠菌会由菌丝相向酵母相转移[52]。Okkers等[53]鉴定出一种戊糖乳杆菌中的长肽TV35b,这种肽可以通过群体感应系统使白念珠菌转化为酵母相,诱导假菌丝的形成,从而降低白念珠菌致病性并抑制其生长。
1.4 蛋白类物质
乳酸菌产生的蛋白质类化合物一般分为两类:一类是由核糖体合成的抑菌肽,上述提到的细菌素类就属于此类;另一类是通过蛋白酶酶解产生的肽类,而非核糖体合成,以下提及的蛋白类物质就是通过细菌蛋白酶酶解作用产生的。
Niku-Paavola等[54]在植物乳杆菌VTT E-78076(Lb.plantarumVTT E-78076)培养滤液中发现了具有抑制革兰氏阴性菌成团泛生菌(Pantoea agglomeransVTT E-90396)和燕麦镰刀菌VTT D-80147(Fusarium avenaceumVTT D-80147)生长的抗菌物质,其中包括细菌蛋白酶酶解产物甘氨酸-L-亮氨酸环肽。除了对细菌的抑制作用,一些报道中也提到植物乳杆菌MiLAB 393可以通过酶解作用产生一种具有抑制真菌生长的环肽(L-Phe-L-Pro),同时清酒乳杆菌、冠状乳杆菌和戊糖片球菌酶解产生的环肽(L-Phe-trans-4-OH-L-Pro)也具有抑制真菌生长的作用[55]。而Magnusson等[56]进一步研究发现,棒状乳杆菌Si3(Lb. coryniformisSi3)通过酶解作用产生的抑菌环肽,可以通过其疏水性基团迅速吸附到真菌细胞上,或者形成自聚体从而对霉菌和酵母菌有一定抑制作用。乳铁蛋白是一种具有抗炎、抗氧化和抗菌等活性的蛋白质,同时也是抗菌肽的重要来源[57]。而近期的研究发现乳酸菌可以酶解乳铁蛋白,可以产生一种抗真菌肽——乳铁素[58],这种物质是带有1 个二硫键、多个正离子、具有疏水性和18 个残基环的多肽,可以通过破坏真菌细胞膜和膜上质子梯度达到抑制真菌的作用[59]。而另一种乳铁蛋白肽是由乳铁蛋白氮端酶解掉的前11 个氨基酸残基组成,在结构上具有高度的阳离子性质并包含疏水性的缬氨酸和色氨酸,因此也可通过破坏细胞膜来抑制白念珠菌和烟曲霉的生长[60]。此外,从阿尔及利亚传统发酵食品中发现一种抗菌肽SSSEESII,这种物质是通过乳球菌BR16菌株胞外丝氨酸金属蛋白酶水解牛酪蛋白得到的。经鉴定,该肽是酪蛋白αs2亚基的24~31片段,并且经实验发现该肽对两种革兰氏阳性细菌(李斯特菌ATCC 33090和黄体微球菌ATCC 4698)和两株革兰氏阴性菌(大肠埃希氏菌ATCC 25922和肠化沙门氏菌ATCC 13076)均具有抗菌活性[61]。
1.5 其他物质
1.5.1 胞外多糖
胞外多糖是乳酸菌的一种代谢产物,可分为只含单一类型单糖单体的同聚多糖和含多种不同类型单糖单体的杂聚多糖[62]。近些年的研究中发现胞外多糖具有抑菌活性,Bajpai等[63]发现植物乳杆菌8513和BT8513产生的胞外多糖可以有效抑制金黄色葡萄球菌的生长。同时Trabelsi等[64]发现乳酸杆菌Ca6产生的胞外多糖可以有效抑制常见致病菌的生长。此外,植物乳杆菌YW32胞外多糖可以有效抑制腐败菌和致病菌生物膜的形成,从而提高食品安全性[65]。
1.5.2 二氧化碳
乳酸菌异型发酵可产生二氧化碳,二氧化碳可以有效抑制革兰氏阴性需氧菌和霉菌。二氧化碳的抑菌机制通过以下途径:二氧化碳的产生可以形成厌氧环境,从而使得需氧型微生物受到抑制;并且二氧化碳可以溶于细胞液中,导致pH值下降从而导致细胞内一些酶活性下降,不仅影响了细胞代谢,还降低了细胞膜的传递能力[40]。
1.5.3 过氧化氢
过氧化氢是由乳酸菌中黄素氧化酶在有氧条件下产生的,并且因为大多数乳酸菌中没有过氧化氢酶,使得过氧化氢在环境中积累,而过氧化氢可以通过氧化作用破坏羟自由基,从而抑制微生物生长[66]。
1.5.4 罗氏菌素
3-羟基丙醛又称罗氏菌素,是罗伊氏乳杆菌(Lactobacillus reuteri)、短乳杆菌(Lactobacillus brevis)、布氏乳杆菌(Lactobacillus buchneri)等在厌氧条件下分解甘油产生的一种广谱抑菌物质,可以有效抑制革兰氏阳性菌和阴性菌、霉菌和酵母[67]。罗氏菌素可以抑制核糖核酸还原酶活性,而这种酶是DNA合成所必需的,因此罗氏菌素可以通过干扰细菌DNA复制,来抑制细菌增殖[68](机制类似于图1机制四中抑制DNA复制过程)。除此之外,一些研究也发现乳酸菌代谢产物2-丙烯基酯具有抑制真菌的活性[69]。同时,乳酸菌在柠檬酸发酵过程中产生的双乙酰,在低pH值条件下也具有抑菌作用[70]。
1.6 竞争作用
虽然乳酸菌可以产生抑菌物质,但是生成量一般低于它们各自的抑菌浓度,所以除了产生抑菌物质,推测还存在其他的抑菌途径。营养竞争是一种重要的竞争方式,通常是指乳酸菌同腐败菌对营养物质进行竞争,其可通过较快的速度利用营养物质,造成腐败菌失去营养,进而生长受到抑制。这种营养竞争既可以单独作为一种作用机制,也可以与乳酸菌产生的抑菌物质协同作用[7]。Honoré等[71]监测3 种副乳杆菌对营养物质的消耗与生长抑制的关系时发现这些乳杆菌对葡萄糖和谷氨酰胺的消耗与腐败霉菌的生长呈负相关,进而证明了乳杆菌可以竞争利用营养物质从而导致腐败菌缺少营养而生长受到抑制。
2 乳酸菌作为生物保护菌在食品中的应用
2.1 乳及乳制品
乳制品因其具有较低pH值、中等水分活度和丰富的营养物质极易被微生物污染而腐败变质,其中霉菌和酵母菌是造成乳制品变质的主要微生物[72]。霉菌属中的青霉菌属(Penicillium)、毛霉菌属(Mucor)和枝孢菌属(Cladosporium),酵母菌属中的假丝酵母属(Candida)、克鲁维酵母属(Kluyveromyces)和亚罗酵母属(Yarrowia)都是乳制品中常见有害微生物[73]。近些年的研究发现多种乳酸菌不仅可以作为生物保护菌抑制乳制品中真菌的生长,还能作为一种发酵剂赋予乳制品特殊风味。
解脂亚罗酵母(Yarrowia lipolytica)被认为是一种乳制品中很难抑制的致腐微生物,而Mieszkin等[74]发现哈尔滨乳杆菌K.V9.3.1Np(Lb. harbinensisK.V9.3.1Np)产生的代谢产物尤其是有机酸如乙酸、乳酸、2-吡咯烷酮-5-羧酸、己酸和2-羟基苯甲酸,虽然不会改变酵母细胞膜通透性,但可通过诱导酵母细胞膜去极化的方式阻止解脂亚罗酵母的生长。而且哈尔滨乳杆菌K.V9.3.1Np在对酵母有良好的抑制作用的同时不会破坏发酵乳的感官特性,因此十分适合作为一种生物保护菌应用于发酵乳的生产[75]。而Garnier等[76]从发酵乳制品中分离出的3 株乳酸菌——詹氏丙酸杆菌CIRM-BIA1774(Propionibacterium jenseniiCIRM-BIA1774)、鼠李糖乳杆菌CIRM-BIA1952(Lb. rhamnosusCIRM-BIA1952)和Mucor lanceolatusUBOCC-A-109193也被发现具有抗真菌活性。通过对这3 株乳酸菌上清液的分离和鉴定,一共发现56 种具有抗真菌活性的物质,其中含量最多的抗真菌物质是詹氏丙酸杆菌发酵产生的丙酸和乙酸,鼠李糖乳杆菌发酵产生的乳酸和乙酸以及M. lanceolatus发酵产生的丁酸。除了通过产生有机酸抑制有害微生物的生长,最新的研究发现鼠李糖乳杆菌还可以发酵酪蛋白αs2产生含有9 个氨基酸且具有抗菌活性的多肽。通过体外实验证实了这种抗真菌肽可以有效抑制葡萄状白霉(Mucor racemosus)和胶红酵母(Rhodotorula mucilaginosa)的生长[76]。此外,植物乳杆菌L244、哈尔滨乳杆菌L172、短乳杆菌L128和鼠李糖乳杆菌CIRM-BIA113被应用于乳制品中抑制青霉菌和毛霉菌的生长,这些乳酸菌可以通过代谢产生乙酸、羟基苯乳酸、苯乳酸、3-苯基丙酸、3-(4-羟基苯基)丙酸和5-氧吡啶-2-羧酸、二乙酰基、乙酰乙酸等达到抑制真菌的目的[77]。
2.2 肉及肉制品
肉及肉制品含有丰富的营养物质及大量的水分,因此在加工、贮藏和销售环节很容易受到假单胞菌、大肠埃希氏菌、单核细胞增生李斯特菌、金黄色葡萄球菌和沙门氏菌等有害微生物的污染[78]。肠杆菌科、假单胞菌和热杀索丝菌是真空包装生牛肉切片在4 ℃贮藏过程中的主要腐败菌,其中假单胞菌随着贮藏时间的延长逐渐成为优势腐败菌,而Zhang Yimin等[79]发现在真空包装牛肉切片中接种清酒乳杆菌和弯曲乳杆菌可以显著抑制肠杆菌科、假单胞菌和热杀索丝菌3 种腐败菌的生长,并且能够有效降低生牛肉切片微生物的多样性及贮藏过程中挥发性盐基氮类物质的产生。Orihuel等[80]发现肠球菌CRL35(Enterococcus mundtiiCRL35)因产生一种蛋白类物质,可以抑制大肠埃希氏菌O157:H7 NCTC12900(E. coliO157:H7 NCTC12900)氨基酸和核苷酸的代谢和运输,从而导致大肠埃希氏菌生长受阻,同时添加肠球菌也可以有效降低大肠埃希氏菌胞外基质蛋白的黏附能力,使大肠埃希氏菌致病性降低。此外,Li Peijun等[81]将发酵乳杆菌(Lactobacillus fermentum)接种到灌肠原料中,发现发酵乳杆菌不仅可以有效抑制其中肉杆菌属、热杀索丝菌属和粪肠球菌属等腐败菌的生长,还可以将高铁肌红蛋白转化为红色的亚硝基亚铁肌红蛋白,使肉色得到改善。正是因为发酵乳杆菌在灌肠原料中的抑菌作用和护色作用,其被认为是一种亚硝酸盐的替代物在肉制品生产中具有潜在应用价值。乳酸菌作为生物保护菌除了可以应用于冷鲜肉和原料肉中还可以应用于熟肉制品中,徐梅[82]将生物保护菌清酒乳杆菌B2(Lactobacillus sakeiB2)和植物乳杆菌R2(Lactobacillus plantarumR2)喷涂于切片低盐香肠表面,结果表明在贮藏期间两种生物保护菌能明显改善香肠的菌群结构,显著抑制假单胞菌和热杀索丝菌的生长,有效延长香肠的货架期;同时,能有效缓解香肠贮藏过程中的脂肪氧化和蛋白氧化,提高了产品的感官品质。此外,Danielski等[83]在研究熟火腿保鲜时,发现将乳酸菌麦芽香肉杆菌作为生物保护菌添加到产品中,可以有效抑制单核细胞增生李斯特菌的生长,提高食品安全性。
2.3 水产品
与肉制品类似,水产品富含蛋白质和不饱和脂肪酸等营养物质,易受到微生物污染发生腐败变质[84]。为了防止因水产品的腐败而造成的浪费和安全问题,乳酸菌作为生物保护菌近几年被广泛用于水产品中。其中乳酸乳球菌是一种良好的生物保护菌,Ibrahim等[85]将乳酸乳球菌添加到真空包装生三文鱼中后发现乳酸乳球菌不仅可以显著延长产品货架期,而且不会改变产品原有感官特性。除此之外,一株可以产生细菌素的戊糖乳杆菌也被应用于生鲜三文鱼片保鲜,其可以控制鱼肉中嗜水气单胞菌和单核细胞增生李斯特菌的生长,并且即使冷链遭到破坏也可以起到保护作用[86]。除了应用于生鲜水产品中,乳酸菌作为生物保护菌还被应用于加工后的水产品中。Saraoui等[87]将鱼乳球菌CNCM I-4031(L. pisciumCNCM I-4031)和广布肉杆菌V41(Carnobacterium divergensV41)应用于气调包装去皮熟虾的8 ℃贮藏过程中,研究发现两株乳酸菌作为生物保护菌可以有效抑制单核细胞增生李斯特菌、希瓦氏菌和肠杆菌的生长。并且鱼乳球菌CNCM I-4031除了可以抑制微生物的生长,还可以有效消除不良风味物质,减缓去皮熟虾贮藏过程中的品质劣变。此外,Aymerich等[88]研究表明清酒乳杆菌CTC494(Lb. sakeiCTC494)、弯曲乳杆菌CTC1742(Lb. curvatusCTC1742)和麦芽香肉杆菌CTC1741(Carnobacterium maltaromaticumCTC1741)在熏鱼贮藏过程中可作为生物保护菌延长其货架期。其中,虽然弯曲乳杆菌和麦芽香肉杆菌可以利用肉中蛋白产生类细菌素物质来增强抑菌能力,但这3 种乳酸菌中清酒乳杆菌保护作用最强,在贮藏期间可以有效抑制单核细胞增生李斯特菌的生长,而弯曲乳杆菌则只能减缓单核细胞增生李斯特菌的增长速度,相对以上两种乳酸菌麦芽香肉杆菌作为生物保护菌应用范围较窄,虽然在体外实验中表现出了抗菌活性,但仅在熏鱼产品中发现其抑菌性能,而在其他实际产品中还没有呈现出显著抑菌效果。
2.4 水果及蔬菜
新鲜果蔬由于富含有益于人体健康的物质(例如维生素、矿物质和膳食纤维等)深受消费者喜爱。但新鲜果蔬容易受到微生物污染,货架期相对较短。而常用的热加工处理和添加化学防腐的杀菌方法会破坏果蔬的感官和营养特性89]。相比之下,将生物保护菌——乳酸菌喷涂在果蔬表面因既不会破坏果蔬品质和营养,又可以起到抑制有害微生物的作用,而深受研究者青睐。Ma Jiahong等[90]将栖树汁乳杆菌JT03(Lb. sucicolaJT03)和乳酸片球菌JY03(Pediococcus acidilacticiJY03)应用于防治柑橘霉菌病中,研究发现这两种乳酸菌可以利用产生的蛋白类抑菌物质来抑制指状青霉的生长。除了霉菌之外,单核细胞增生李斯特菌也是果蔬中常见的有害细菌。Trias等[91]在对10 株肠系膜明串珠菌(Ln.mesenteroides)和1 株柠檬明串珠菌(Ln. citreum)对抗单核细胞增李斯特菌的实验中发现,这11 株明串珠菌一般通过产生有机酸、过氧化氢和细菌素等抑菌物质来抑制单核细胞增生李斯特菌的生长。而这11 株明串珠菌中的2 株肠系膜明串珠菌CM135和CM160因其可以产生IIa类细菌素而对单核细胞增生李斯特菌有更好的抑制作用。同时在模拟金冠苹果和生菜叶被单核细胞增生李斯特菌污染的实验中,这两株肠系明串珠菌同样表现出对单核细胞增生李斯特菌明显的抑制作用。
3 结 语
乳酸菌作为生物保护菌可以通过多种机制抑制食品中细菌和真菌生长,降低食源性微生物腐败和致病风险,可以有效延长食品保质期。并且大量的研究已经证实乳酸菌在食品中除了具有抑菌活性,还具有降解真菌毒素、降低不良风味、改良食品颜色和赋予食品特殊风味等作用。因此乳酸菌作为生物保护菌应用于食品中日益受到研究者青睐。
然而由于乳酸菌种类较多,代谢方式多样,代谢物种类丰富,目前研究多集中在小分子代谢物及其抑菌机制,而对蛋白类物质研究较少,因此针对乳酸菌产生的蛋白类抑菌物质及其抑菌机理有待进一步的探究。与此同时,乳酸菌已被证实对很多细菌和真菌有抑制作用,但是很少有研究关注乳酸菌作为生物保护菌添加到食品中后,乳酸菌自身对食品感官、风味和营养等性质的影响,而这一问题将会成为乳酸菌作为商用生物保护菌必须考虑的问题。此外,乳酸菌作为生物保护菌通常被应用于发酵制品和原料食品保鲜中,扩展生物保护菌的应用范围也是未来研究和应用中的重点。
