内陆干旱区典型旱生植物蒸腾耗水量模拟研究
2021-10-29张阳阳刘秀强
张阳阳, 陈 喜,4,*,高 满, 刘秀强
1 天津大学地球系统科学学院,天津 300072 2 天津大学表层地球系统科学研究院,天津 300072 3 天津市环渤海关键带科学与可持续发展重点实验室,天津 300072 4 河海大学水文水资源学院,南京 210098
西北干旱区降水量少、潜在蒸散发量大、土壤含水率低,生长一系列依赖地下水的旱生植被。随着气候变化和人类活动影响,过量开采地下水造成的水位下降,引发植被退化、沙漠化等一系列生态环境问题。近年来,我国实施的一系列生态恢复工程,迫切需要掌握植被耗水规律,为区域水资源和生态环境演变影响评估提供科学支撑。
植物耗水量计算分为单株植物耗水量、植物群落耗水量[1]。单株植物耗水量观测和计算主要方法有:整容器法、快速称重法、液流法(热脉冲、热扩散、热平衡)。整树容器法简便易行、计量精确,通常将其作为基准测算树木蒸腾的准确性[2]。快速称重法通过剪取枝叶在田间进行两次间隔称重,用离体失水量和间隔时间换算蒸腾速率,代表正常生长状况下的蒸腾速率[3]。该方法的前提是假设枝叶离体短时间内蒸腾改变不大。液流法通过测定树干部液流量来计算树冠部蒸腾量,方法简单且精度高[4]。植被群落主要计算方法有:水量平衡法、微气象法、红外遥感法[1]。水量平衡法通过计算区域内水量的收入支出差额来推求蒸散量。微气象法则主要基于修正的Penman-Monteith公式计算植物蒸腾量[5]。红外遥感法根据植被的光谱特性、红外信息,结合微气象参数计算蒸腾量。受观测条件的限制,整树容器法等测量方法难以大范围推广,水量平衡法、微气象法等一般未考虑植物生理特征的差异,难以辨别区域内不同植物蒸腾量的差异。
西北干旱区不同流域典型植物耗水量已有较多研究(表1),已有结果表明:由于植物生长状态和供水条件、计算和实测方法的差异,蒸腾耗水量变化范围大。同一种植物在不同供水条件下植物耗水量存在差异,如胡杨(Populuseuphratica)在常年输水河段和间歇性输水河段的生长季耗水量分别为990、414 mm[15];柽柳(Tamarixchinensis)在土壤含水量为70%田间持水量下的生长季耗水量为500 mm,当土壤含水量为35%田间持水量时,生长季耗水量降低为235 mm[24];灌溉的人工梭梭林(Haloxylonammodendron)耗水量为534 mm[9],荒漠区天然梭梭耗水量仅为49 mm[28];湿地芦苇(Phragmitesaustralis)耗水量可达1500 mm[31],沙生芦苇耗水量可减少到173 mm[18]。植物耗水量还与树龄(通常用基径表示)有关,如民勤地区基径3.03 cm的柽柳耗水量是基径1.5 cm耗水量的6倍[9];梭梭直径从22.9 cm降低至9.9 cm,生长季耗水量从113 mm减少至92 mm[29]。不同测量方法实测的植物耗水量存在差异,如在黑河流域下游,液流法(热脉冲技术)得到胡杨、梭梭生长季耗水量分别为154[7]、127 mm[29],而基于水量平衡法计算结果分别为520[6]、213 mm[18];在塔里木河流域,柽柳生长季耗水量液流法得出761 mm[18],模型模拟得出472 mm[25]。

表1 干旱区典型植物生长季耗水量已有研究方法和结果
以上实测和计算方法,通常仅得到实测土壤水分、气象或者植物生长状态下蒸腾耗水量,难以预测不同气候、供水和生长状态下植物蒸腾量变化。自Philip提出“土壤-植物-大气”连续体概念后,构建的植物耗水量模型,具有分析植物生理特征参数(如气孔导度等)与气象条件以及土壤水分因子关系的功能,成为模拟和预测植物蒸腾量及其变化的重要途径。估算气孔导度对环境因子响应的模型主要为经验模型(如Jarvis模型)和半经验模型(如Ball-Woodrow-Berry(BWB)模型两类[34]。Jarvis模型建立气孔导度与环境因子的单一函数[35],BWB模型旨在描述气孔导度与光合速率的关系[36]。但无论是Jarvis模型还是BWB模型,都未考虑由根部到冠层的水分运输过程中植物根源信号对植物气孔导度的影响。
本文运用Tardieu-Davies模型[37],考虑植物根源信号对植物气孔导度的影响,参照石羊河尾闾湖(青土湖)气象条件和典型植物的生理特征数据,利用前期适宜和极限生态地下水埋深下土壤水分状态研究成果[38],计算适宜和极限生态地下水埋深下植物耗水量,将模型计算结果与国内外现有研究成果对比,分析计算结果合理性和模型适用性,为流域水资源管理和生态环境修复提供科学依据。
1 Tardieu-Davies模型
SPAC水分传输理论明晰了水分传输途径,即由土壤到达植物根表皮、根系、茎,最后到达叶片,再由叶片气孔扩散到空气层。Tardieu-Davies模型[39]将植物体简化为根、主干和叶片三部分(图1)。根源信号可作为植物在日尺度上感知土壤水分获取能力(土壤水分状况或对水流的阻抗)的指标[26],脱落酸(ABA)参与对作物气孔行为调控。根源信号ABA的强弱依赖于土壤水势(Ψsoil)和根水势(Ψr)的共同作用;主干对根源信号起到传输和稀释作用;叶片气孔导度受传导来的根源信号ABA的影响;最终,水汽压差(VPD)和气孔导度(gs)等决定植物的蒸腾耗水量。

图1 依赖地下水的植物蒸腾过程及土壤水分剖面概化图Fig.1 Transpiration of Phreatophytic Vegetation and soil water characteristic curveJin:根系蒸腾流 Water flux entering into roots;Jxyl:木质部水流 Water flux in the xylem;Jout:叶片蒸腾流 Transpiration flux;Ψsoil:土水势 Water potential in the rhizosphere;Ψr:根水势 Water potential in the root;Ψxyl:木质部水势 Water potential in the xylem;Ψbundle:叶片水势 Water potential in the bundle sheaths;[ABA]xyl:木质部ABA浓度 The amount of synthesized ABA in the xylem;VPD:水汽压差 Air vapour pressure deficit;Rn:净辐射强度 Net radiation;ETg:地下水蒸发 Groundwater evaporation;Ψ0:潜水位 Groundwater table;Ψr1:根水势 Water potentials in the root;Ψbundle:叶片水势 Water potential in the bundle sheaths;θ:含水率 Water content;Ψ:水势 Water potential;P:降水 Precipitation;Tp:蒸腾 Transpiration
植物叶片蒸腾流的计算公式为:
(1)
式中,Jout为植物叶片蒸腾流(mg/s),S为叶面积(m2),s为饱和水气压对温度的斜率(Pa/℃),Rn为净辐射强度(W/m2),ρ为空气密度(kg/m3),Cp为定压比热(1012 J kg-1K-1),ga为边界层阻力(mol m-2s-1),VPD为水汽压差(KPa),λ为蒸发潜热(J/kg),γ为干湿球常数(Pa/K),gs为气孔导度(mol m-2s-1)。式中各分量计算公式为:
gs=gsmin+αexp[β[ABA]xylexp(δΨbundle)]
(2)
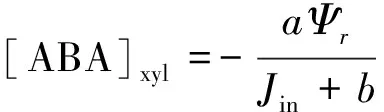
(3)
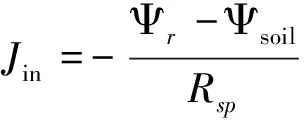
(4)

(5)
式中,gsmin为气孔导度最小值(mol m-2s-1);α、β和δ为常数;[ABA]xyl为木质部ABA浓度(pmol/m3),Ψbundle为叶片水势(MPa),Ψr为根水势(MPa),Jin为根系蒸腾流(mg/s)。a(μmol kg m-3s-1MPa-1)、b为经验系数,通过拟合根水势与木质部ABA浓度进行确定。Ψsoil为土壤水势(MPa),Rsp为根土界面阻力(MPa s-1mm-3);d为相邻根系平均距离(m),r为根系半径(m),k(Ψsoil)为某一水势下的导水率(cm/d)。
将植物蒸腾流Jout转化为蒸腾耗水量Tp(mm/s):
(6)
模型中主要参变量计算方法如下:
(1)气象要素相关参数(Rn、s、VPD、ga):
净辐射强度(Rn):
(7)
式中,PPFD为光照强度(μmol m-2s-1),PPFD=4.72R,R为太阳辐射(W/m2)。
饱和水气压对温度的斜率(s):
s=-0.1849T2+1.005T+50.87
(8)
式中,T为摄氏温度(℃)。
水汽压差(VPD)[40]:
(9)
式中,RH为相对湿度(Pa/Pa),在没有实测资料时,可根据比湿(g/g)、压强(Pa)计算[41]。
(2)植物生理特征参数(gsmin、d、r、Ψr、Ψbundle):
植物气孔导度(gsmin)一日内变化最小可达0.01[42],因此gsmin=0.01。Ψr依据土水势、根土界面阻力和根系蒸腾流进行计算。
相邻根系平均距离d:
(10)
式中,Lv为根长密度(m/m3),即单位体积土壤中的根系总长度。根水势(Ψr)、叶水势(Ψbundle)为:
Ψr=Ψsoil-Rsp×Jin
(11)
(12)
式中,Gr为根土界面到木质部的水力导度(mg MPa-1s-1),计算公式为:
(13)
式中,timephoto为光周期(h),一天中PPFD大于10 μmol m-2s-1的时段即为光周期长,Ψxylmax、Ψxylmin分别为木质部最大、最小水势(MPa)。木质部根水势最小、最大采用-0.45、0.05 MPa[39,43]。
(3)土壤水力参数(k(Ψsoil)、Ψsoil):
土壤导水率(k(Ψsoil))采用Van Genuchten模型(V-G)[44]:
(14)
土水势(Ψsoil)计算公式为:
(15)

采用V-G模型计算θ:
(16)
式中,α1为与进气值相关的参数,h为压力水头(cm),λ为与土壤特性相关的形状系数,m、n为土壤水分特征曲线的形状参数。
将式(7)—(16)计算所得参数带入式(1)—(5),最终依据式(6)可计算出植物蒸腾水量。
2 气象数据及模型参数
甘肃省民勤县青土湖(38°52′30″—39°7′30″ N、103°22′30″—103°37′30″ E)为典型大陆性干旱荒漠气候区,以该区域的气象数据为背景,采用全球陆地数据同化系统(GLDAS:Global Land Data Assimilation System)2009—2018年气象数据,时段间隔3 h,空间分辨率为0.25 °(http://www.shareresearch.me/)。气象要素包括:太阳辐射(R)、温度(F)、比湿(qe)、压强(Pa)。研究期内,GLDAS数据库与中国气象数据网(http://data.cma.cn/)逐日数据相比,得出两个数据太阳辐射、温度、水汽压差的决定系数(R2) 为0.89、偏离率(bias)为20%。
植物生长期(4月1日—9月30日)各气象要素多年平均日变化过程中(图2),光照强度(PPFD)变化范围为400—800 μmol m-2s-1,温度(T)变化范围为9—30℃,水汽压差(VPD)变化范围为0.9—3.2 kPa,最高值大都出现在7、8月份。
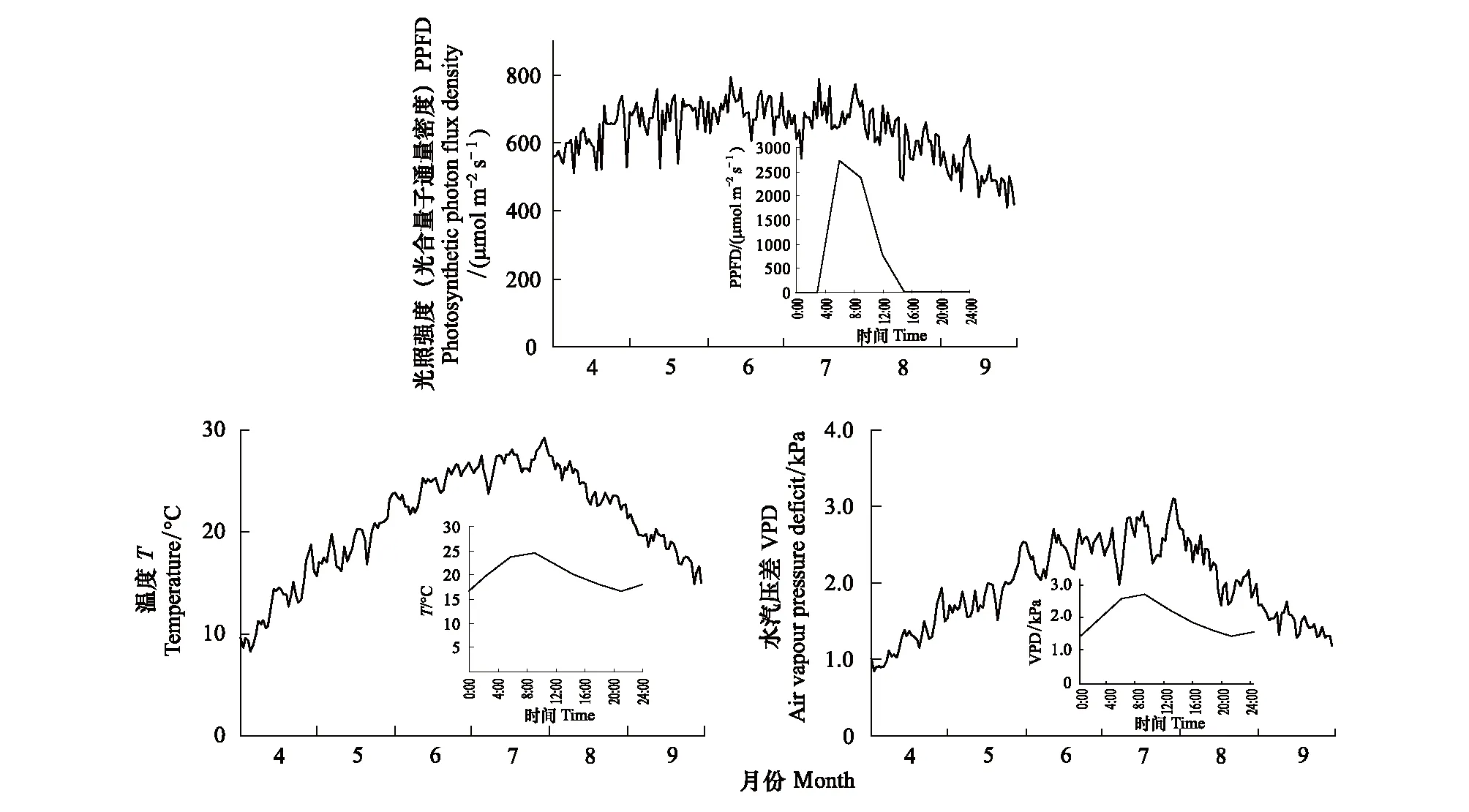
图2 PPFD、T、VPD生长季逐日变化(小图为5月19日逐时变化)Fig.2 Daily variations of PPFD, T, and VPD during growing season(The small figure shows hourly PPFD, T, and VPD on May 19)
根据前期研究结果[38],得出上述典型植物的适宜、极限生态地下水埋深,以及根据土壤水分特征曲线(V-G模型)确定的不同土壤类型和质地根系含水率(田间持水量的70%和67.5%),即表2中土壤含水率(θ1、θ2)。参照国内外文献得出典型植物生理特征参数,即植物根系半径(r)、根系密度(Lv)和α数据(表2)。植物其它生理特征及耗水计算公式参数均采用统一值[39](表3)。
3 结果
3.1 植物蒸腾耗水量计算结果与验证
将研究区气象数据和模型参数输入Tardieu-Davies模型,计算适宜、极限生态地下水埋深下7种典型植物生长期多年平均日蒸腾过程及生长季耗水量,在适宜生态地下水埋深下生长季蒸腾耗水量介于279—1292 mm,平均为793 mm;极限生态地下水埋深下生长季耗水量为130—1050 mm,平均为602 mm。
生长季内不同植物蒸腾耗水量存在较大差异。水生植物芦苇和河岸带植物柽柳的蒸腾量较大,旱生植物梭梭蒸腾量最小,盐节木、罗布麻、白刺和胡杨蒸腾量依次减少(表4)。在适宜生态地下水埋深条件下,生长季芦苇、柽柳的蒸腾量分别为1292、1147 mm,梭梭蒸腾量为279 mm,盐节木、罗布麻、白刺和胡杨蒸腾量分别为940、913、534 mm和448 mm;在极限生态地下水埋深下,芦苇、柽柳的蒸腾量分别为1050、999 mm,梭梭蒸腾量为130 mm,盐节木、罗布麻、白刺和胡杨蒸腾量分别为706、686、345 mm和298 mm。
绘制7种植物生长季蒸腾耗水量变化范围,与表1文献中得出的内陆干旱区典型植物生长季耗水量统计结果相比较。整体来看,Tardieu-Davies模型计算得到的不同供水条件下植物生长季耗水量在现有研究成果的范围内,适宜、极限生态地下水埋深下本文模拟结果与已有研究成果的配对t检验,P值分别为0.12、0.94,均大于0.05,即模拟值与已有文献成果的差异性不显著,说明本文模拟结果可靠。已有文献中植物蒸腾量来源于石羊河、塔里木河、黑河,存在气象条件、植物基径(树龄)差异,因此植物蒸腾量统计值变化范围大。

表2 土壤水力特征和植物生理特征参数

表3 模型参数取值
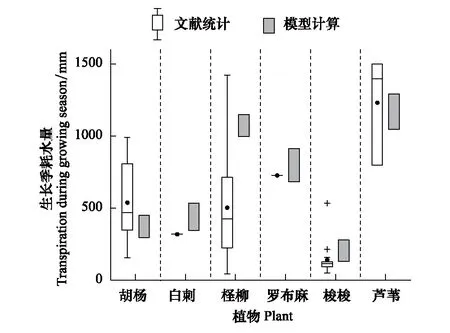
图3 模型计算植物蒸腾耗水与已有成果对比 Fig.3 Comparison of plant transpirations between simulation results and existing statistical results
3.2 生长季植物蒸腾耗水量变化特征
植物蒸腾耗水量变化如图4,与光照和温度逐月变化(图2)相似,呈现先增加后减少的变化形态,最大值、最小值分别出现在7月、4月。在7月,适宜、极限生态地下水位埋深下蒸腾量平均值(表5中Tp1、Tp2)分别为161、122 mm;远大于4月蒸腾耗水量,平均值分别为99、74 mm。
在适宜生态地下水位埋深下,耗水量大的芦苇、柽柳7月蒸腾耗水量分别为256、232 mm,4月蒸腾耗水量的平均值分别为166、142 mm;耗水量最小的梭梭7月蒸腾耗水量为58 mm,4月蒸腾耗水量为34 mm。在极限生态地下水位埋深下,耗水量大的芦苇、柽柳7月蒸腾耗水量分别为210、203 mm,4月蒸腾耗水量的平均值分别为133、123 mm;耗水量最小的梭梭7月蒸腾耗水量为29 mm,4月蒸腾耗水量为14 mm。
日时间尺度上(图4中5月19日)蒸腾耗水量变化大。植物蒸腾耗水量变化趋势与光照和温度变化趋势相似:在光照和温度较大的时刻植物蒸腾耗水量多,光照和温度较小时植物蒸腾耗水量少。不同植物蒸腾耗水量在日尺度上也存在极大的差异。以5月19日为例,适宜生态地下水埋深条件下,芦苇、柽柳的日蒸腾量分别为7.1、6.3 mm,而梭梭日蒸腾量仅为1.5 mm,其它植物如盐节木、罗布麻、白刺和胡杨蒸腾量分别为5.1、5.0、2.9、2.5 mm;在极限生态地下水埋深下,芦苇、柽柳的蒸腾量分别为5.7、5.5 mm,梭梭蒸腾量为0.7 mm,盐节木、罗布麻、白刺和胡杨蒸腾量分别为3.9、3.7、1.9 mm和1.6 mm。

图4 典型植物生长季逐日蒸腾量(小图为5月19日日时间尺度蒸腾耗水量变化过程)Fig.4 Daily transpiration of typical plants during growing season (The small figure shows diurnal change of transpiration on May 19)
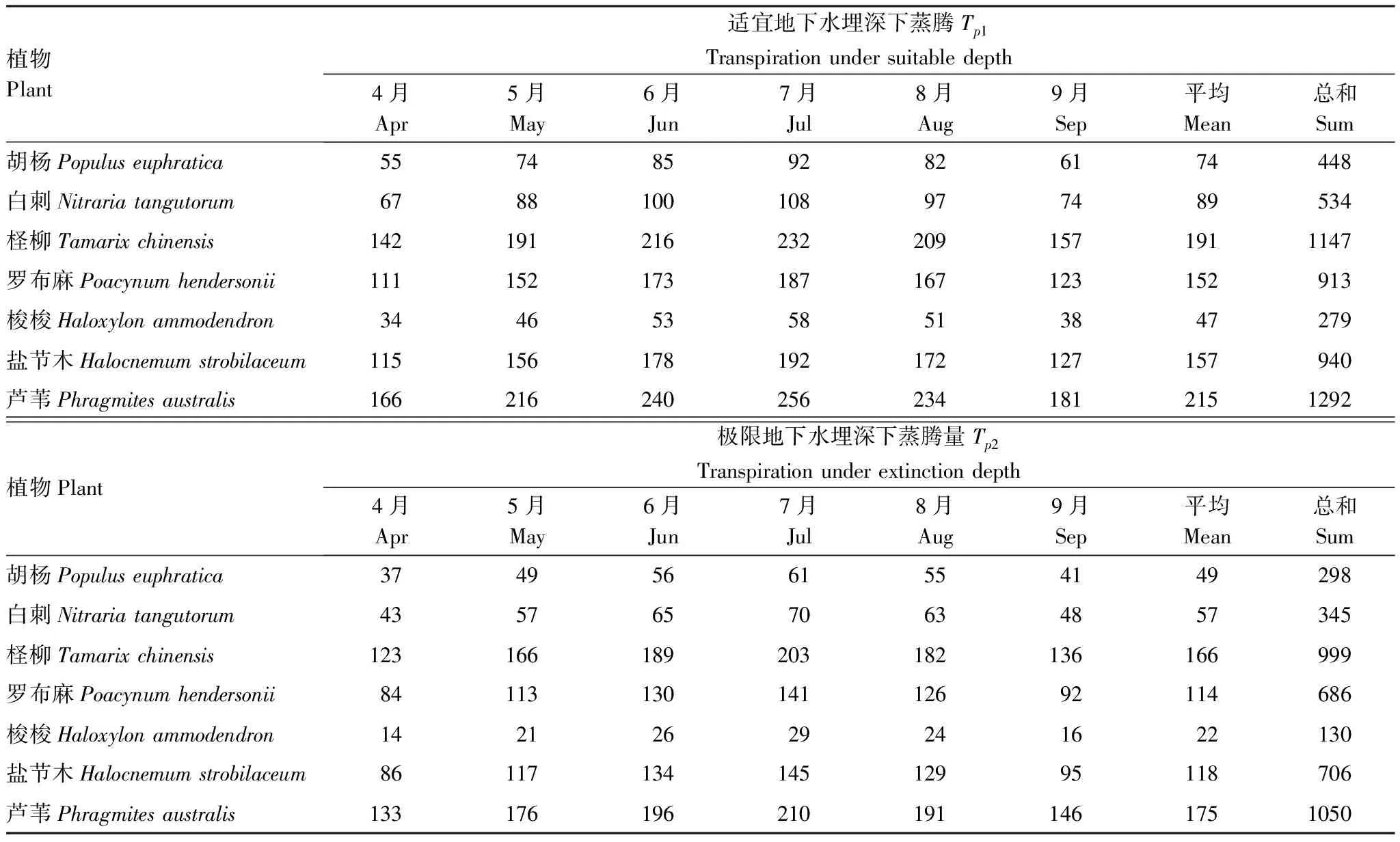
表4 典型植物蒸腾量/mm
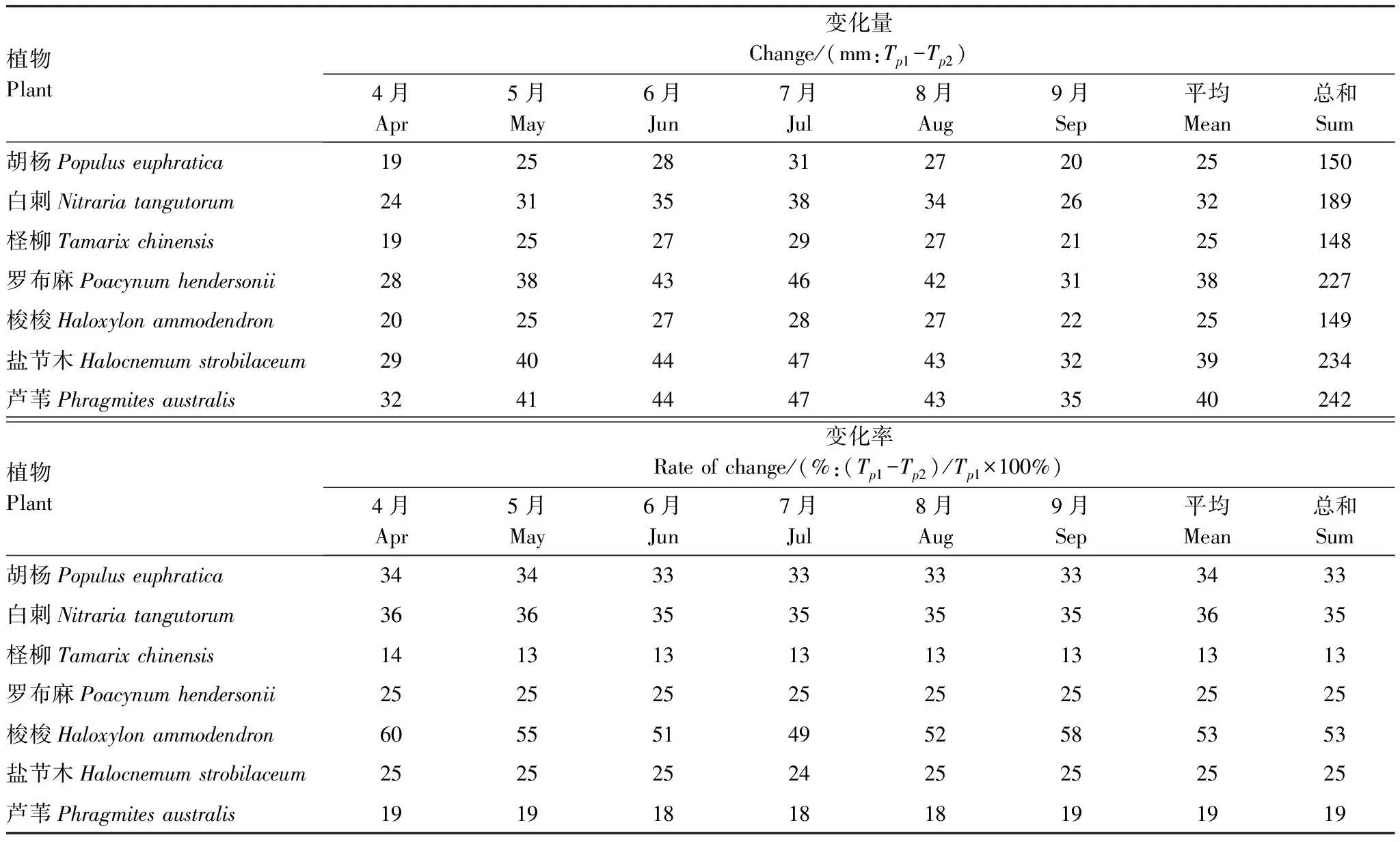
表5 适宜和极限生态地下水位埋深下植物蒸腾耗水量变化
3.3 供水对植物蒸腾耗水量影响
随着地下水位埋深加大,荒漠植物根系所能利用的水分降低,植物可通过调节生理特征(如气孔导度)以减少蒸腾耗水。对比适宜生态地下水埋深增大到极限生态地下水埋深下植物蒸腾耗水量(表5),7种植物月平均蒸腾耗水量减少13%—53%,平均减少29%。不同植物蒸腾量减少程度有所差异,耗水量大的植物蒸腾量减少较小,如柽柳减少量最小(13%),芦苇次之(19%);耗水量小的植物蒸腾量减少较大,如梭梭减少量最大(53%);白刺和胡杨次之(35、33%)。
4 讨论与结论
4.1 讨论
本文计算结果表明,不同植物在生长季内和日时间尺度上的蒸腾耗水量均存在显著差异性。芦苇和柽柳的蒸腾耗水量较大,梭梭、白刺和胡杨蒸腾耗水量小。这与不同类型植物根系分布及供水状态有关,植物根系影响水的利用效率以及植物耐旱性[45-46]。芦苇通常生长在河湖湿地,对水依赖性强、消耗量大[47]。胡杨、柽柳为河岸带植物,但两者耗水量差异明显[11,48],胡杨根系延伸范围大(如最深可达地下7.0 m),对极端环境的忍耐力强,在有限水源条件下,会降低蒸腾耗水量以维持自身生长。梭梭和白刺耐旱性最强,耗水量最小[24,27],这与本研究中白刺、梭梭生长季耗水量较小的结果一致。
当地下水位埋深增大时,植物蒸腾耗水减少,如从适宜增大至极限生态地下水位埋深,本文计算的植物蒸腾耗水量平均减少24%,且不同植物的减少量差异明显。说明当地下水位降低时,植物减少蒸腾耗水量以维持自身的生长[13]。植物蒸腾量减少程度与植物水分利用效率有关,在干旱条件下耐旱性强的植物水分利用效率调节程度大,蒸腾耗水降低幅度大[49]。Farid等[15]采用液流计法对常年输水和间歇性输水河段胡杨蒸腾耗水量监测发现,胡杨在间歇性输水河段蒸腾耗水量(396 mm)较常年输水河段的(936 mm)减少57.8%;Dong[24]对柴达木盆地不同供水条件下白刺的蒸腾量研究,得出当土壤含水量由16.8%降低为6.0%,白刺耗水量减少67%。但对水生植物芦苇而言,植物耗水量随水分条件和地下水位埋深变化不大,如Xu等[50]通过监测不同降水年份芦苇耗水量,得出当降水量减少21%,芦苇耗水量无明显降低趋势。
植物在生长季内和日时间尺度上的蒸腾耗水量变化过程与气象因子的变化过程相似。植物蒸腾耗水量变化特征与Dang等[51]采用液流计法得出的变化过程一致,气象因子对蒸腾耗水量变化的影响与Yu等[52]运用涡度相关法分析结果一致,即植物蒸腾耗水量受光照强度、温度和水气压差的影响。
本文运用Tardieu-Davies模型计算的是特定供水条件下(适宜、极限生态地下水位埋深对应的根系水分状态)单株植物生长季蒸腾耗水量,即单位叶面积植物叶片蒸腾流(式(1)中Jout)。应用于区域荒漠植被实际蒸腾耗水量估算,还需要根据不同干旱区气象条件、水分状态和地下水埋深,并结合遥感等方法推求植物覆盖度,扩展式(1)计算功能,推求流域生态系统的蒸腾耗水总量。并可根据适宜、极限地下水位埋深下植被覆盖度等推求的蒸腾耗水量与实际地下水位埋深下植被状态及蒸腾耗水量之间的差值,探讨干旱区地下水位控制对覆被状态及生态耗水量的影响程度。
4.2 结论
本文选择我国西北内陆干旱区7种典型旱生植物,采用Tardieu-Davies模型计算不同地下水埋深(适宜和极限)以及不同植物生长季耗水量,分析其耗水量的差异及其成因,得出以下结论:
(1)建立的Tardieu-Davies模型综合考虑了气象条件(如光照、温度和水汽压差)、植被生理特征(如气孔导度)和水分状态(如根系层土壤含水率)对植物蒸腾耗水的影响。计算的典型旱生植物生长季蒸腾量与已有研究成果对应的植物蒸腾耗水量对比表明,Tardieu-Davies模型计算结果合理,体现出“SPAC”系统中不同植物蒸腾耗水及其对水分的适应性。
(2)在适宜、极限生态地下水埋深下7种植物生长季蒸腾量平均值分别为793、602 mm。不同植物耗水量差异大,芦苇生长季蒸腾量最大,梭梭的最小。适宜、极限生态地下水埋深下,生长季芦苇蒸腾量分别为1292、1050 mm,远大于梭梭的蒸腾量(分别为279、130 mm)。年内植物蒸腾量4月最小、7月最大。
(3)供水条件影响植物蒸腾耗水量,由适宜生态地下水位埋深增大为极限生态地下水位埋深时,植物生长季耗水量平均减少24%。耐旱性强的梭梭、白刺蒸腾量减幅大(53、35%),耐旱性弱的芦苇、柽柳蒸腾量减幅小(19、13%)。
