积雪变化对高寒草甸钝苞雪莲(Saussurea nigrescens)繁殖分配及功能属性的影响
2021-10-29王长庭宋小艳
王 鑫,王长庭,胡 雷,宋小艳
西南民族大学青藏高原研究院, 成都 610041
植物如何将有限的资源在不同功能器官间进行分配以及这种对策带来的适合度利益与代价,一直都是植物生活史进化领域中的研究热点[1]。而植物生长的最基本活动就是从环境中获取资源、利用资源,并对资源进行再分配[2-3]。在这一过程中,植物不仅受到自身内部资源状况的影响,也受到外界环境因子的影响,且环境因子的影响作用可能更大[2,4]。当外界环境因子,如光照、水分、温度或土壤养分发生变化导致资源受限时,植物不同器官之间会对有限的资源产生竞争,此时植物必须对资源进行权衡分配,进而影响植物的营养生长和繁殖生长[2,5-6]。在植物适应环境变化的过程中,植物的繁殖生长对策发挥了重要作用,通过调整资源分配格局,植物将以最佳的繁殖性状适应外界环境,使个体适合度最大化,其中繁殖分配(Reproductive Allocation, RA)又是植物繁殖策略的关键[7-8]。它不仅受外界环境因子的影响,还与种群特征、个体大小、繁育系统和竞争作用等生物因子有关[9]。因此,植物的繁殖分配一方面是权衡生长、生殖、防卫和贮藏等重要功能的结果,另一方面也是物种生活史和外界环境协同进化的结果[2,10-12]。近些年来,大部分研究都表明在不同生境下随着植株个体增大,繁殖绝对投入会增加,而繁殖相对投入会减小[13-14],但孟丽华等[15]发现海北高寒草甸附近的圆穗蓼(Polygonummacrophyllum)并不存在繁殖分配与个体大小的依赖关系。
在植物生长繁育过程中,开花结果是关键物候期,同一物种在不同环境中的开花数量、种子形态和数量特征差异显著,这是植物自身生物学特性和外界环境条件变化共同作用的结果[2,16]。丁少净等[17]发现干旱胁迫可使花苞直径和长度分别下降21.87%和23.7%,Smith等[18]认为单个种子重量与单株种子数量之间具有负相关关系。另外,叶片作为植物进行光合作用的主要器官,驱动和参与了生态系统的物质循环和能量流动[2,19],既体现了植物对环境的生长适应对策,也反映了植物对资源的获取、利用、保存和分配策略[2,20]。茎作为传导器官,连通了植物地下营养吸收和地上同化组织,为植物的生长发育输送水分和营养元素,同时支撑着叶片和花朵,决定了各器官的空间分布格局[2,21-22]。因此,研究植物花苞、叶片和茎等器官的功能性状,对分析植物的生长、繁殖和进化过程对环境的适应策略具有重要意义。
青藏高原位于我国西南部,有“世界屋脊”和“地球第三极”之称,其对于欧亚大陆气候格局的改变具有重要影响作用[23]。同时,青藏高原对气候变化的响应十分敏感,往往能够更早的对全球气候变化产生预警作用,其独特的环境与生物资源多样性也一直为生态学界所瞩目[24]。近年来,由于青藏高原东部广泛存在着季节性积雪,且融雪的时间不尽相同,这使得季节性雪被对植物的生长繁殖、土壤理化性质及微生物活动等一系列生态因子均产生了深刻影响[25],而植物长期生活在这种异质性环境中,必然会在形态结构等方面产生相应的变化[26]。
钝苞雪莲(Saussureanigrescens)是青藏高原菊科风毛菊属常见种之一,多年生草本,高15—45 cm。茎直立,密被长绒毛。叶片呈线状长圆形,长约8—15 cm,顶端渐坚,边缘有倒生细齿。头状花序1—6个在茎顶伞状排列,密被稀疏长绒毛,梗直立。总苞片4—5层,干后呈黑褐色,顶端稍钝。主要分布在甘肃、青海、青藏高原等西部高海拔地区[5,27]。现有研究指出,钝苞雪莲(Saussureanigrescens)的百粒种子重、个体大小与海拔之间具有负相关关系,而繁殖分配与海拔间具有正相关关系;且雄性器官投入与单株种子数随个体大小的增加而增加,性分配随个体大小的增加而减少[5,28]。
目前在高寒草甸的植物功能属性及繁殖分配方面,研究主要集中在海拔、施肥和放牧的影响[5,11,29],如Zhang等[30]发现氮肥添加促进了禾本科植物种子的成熟、提高了其繁殖分配;Nui等[31]发现放牧显著降低了高寒草甸植物的相对丰度和高度;王一峰等[32]发现随海拔的升高川西风毛菊各器官生物量递减,而种子百粒重与繁殖分配则增加。然而,在影响植物功能性状的诸多环境因子中,有关积雪变化对植物的茎、叶、花苞及繁殖分配影响的研究鲜见报道。近年来,青藏高原降雪格局发生了显著变化,稳定积雪区面积逐渐扩大,尤其是在东部地区,雪被积累量与覆盖时间均呈增加趋势,而积雪格局的改变会进一步影响周边生态系统[33-35]。鉴于此,本研究通过对青藏高原高寒草甸优势物种钝苞雪莲的功能属性及繁殖分配等个体特征在积雪梯度上的变化研究,旨在探讨极端环境对高寒草甸植物的影响[36]。
1 材料与方法
1.1 研究区域概况
图1 研究区位置Fig.1 Location of the study area
研究区位于青藏高原东缘四川省阿坝州红原县境内的西南民族大学青藏高原基地(102°35′E,32°49′N),平均海拔3494 m,年平均气温为1.1℃,最高月均温10.9℃,最低月均温-10.3℃,年均降水量为650—800 mm,年均相对湿度60%—70%,年日照2417.9 h。主要草甸类型为矮嵩草草甸,植被盖度可达80%以上,最高高度达到45—60 cm。主要优势种包括:禾本科垂穗披碱草(Elymusnutans)、剪股颖(Agrostisclavata),莎草科矮嵩草(Kobresiahumilis)、苔草(Carexenervis),豆科异叶米口袋(Tibetiahimalaica)、黄花棘豆(Oxytropisochrocephala),杂草钝苞雪莲(Saussureanigrescens)、条叶银莲花(Anemonetrullifolia)和二裂委陵菜(Potentillabifurca)等,土壤类型为亚高寒草甸土[24,37-39]。采样点地理位置详见图1。
1.2 试验设计
1.2.1样地设置
于2013年非生长季(11月),在研究区内选取30 m×30 m、具有典型代表性的矮嵩草草甸作为试验样地,采用随机区组试验设计,均匀布设25个2 m×2 m 的小样方,各小样方间至少距离1.5 m作为缓冲区。在25个小样方内共设置5个积雪梯度处理(CK、S0、S1、S2和S3),每个处理5个重复。其中,CK为对照处理,即自然积雪量,S0为去除积雪,S1、S2、S3分别为自然积雪量的2倍、3倍、4倍。具体操作方法如下:(1)在生长季结束后,非生长季自然降雪开始前,于试验样地周围建立积雪场,并铺上若干2 m×2 m的防水布用于收集降雪;(2)待每次降雪结束后收集防水布上的积雪,分别堆积到不同积雪处理的小样方中,S1、S2和S3分别堆积1块、2块、3块防水布上的雪;(3)人工去除S0处理样方中的积雪[2]。在2017年冬季至2018年春季的试验处理期间内,CK、S1、S2、S3累计积雪量分别为37 cm、72 cm、110 cm、143 cm,每月降雪量详见表1。2018年8月初地上生物量高峰期时测量每个处理下各植物功能群平均高度,详见表2。积雪实验在每年11月至次年4月进行,期间植物处于枯黄、死亡状态,在5—9月份的植物生长期,无自然降雪,故积雪期与植物生长期相互独立,积雪覆盖不会直接造成植物死亡。
1.2.2样品采集与处理
2018年生长季末期(8月下旬),钝苞雪莲果实成熟后,在每个积雪处理梯度的5个小样方内随机选取5株发育完全的钝苞雪莲植株齐地面刈割并编号,每个积雪处理梯度共采取样品25株,放入冰盒内带回实验室分析。

表1 2017—2018积雪处理期内试验区月降积雪厚度
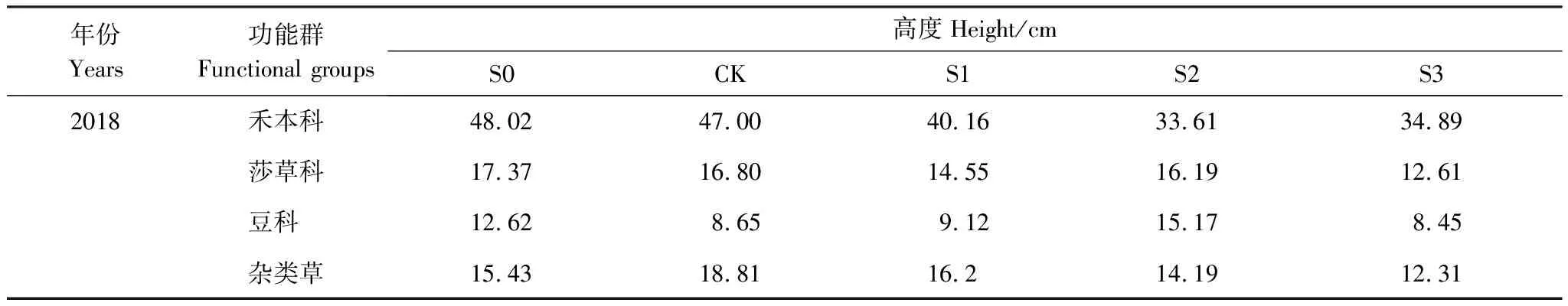
表2 积雪梯度下各功能群平均高度
完整摘取每株样品的所有叶片,记录叶片数量,用多功能扫描仪(Canon, Lide110, Japan)对每株样品进行叶片扫描获取叶面积数据。记录每株样品的花苞数,并利用精度为0.01 mm的游标卡尺测量每颗花苞直径,以及每株样品茎部直径(测量3次);利用精度为0.1 cm的卷尺测量株高、茎高,茎高为茎上第一颗花苞与茎末端之间的长度,株高为整株植物长度,根据株高和茎高的差值计算总状花苞长度,即近基端第一颗花苞到顶端的距离。
将每株样品不同器官分别装在信封袋中于65℃下烘干至恒重,用万分之一天平测量每株植物茎、叶、花苞的生物量,后将所有样品粉碎、研细,以备养分分析。
1.3 样品分析
采用杜马斯燃烧法测量每株植物样品的花苞、叶片和茎的全碳(Carbon,C)和全氮(Nitrogen,N)含量,采用钼锑抗比色法测定全磷(Phosphorus,P)含量。
1.4 数据处理与统计分析
由于完整植物根系获取难度较大,且挖掘采样严重破坏试验样地,因此以地上部分生物量代表植株个体大小(Individual Biomass, IB),以茎、叶生物量代表营养器官生物量(Vegetative Organ Biomass, VB),以植株花苞生物量代表繁殖器官生物量(Reproductive Organ Biomass, RB),另外繁殖分配(RA)以繁殖器官生物量占地上部分总生物量的百分比来表示[1]。采用ImageJ软件(National Institutes of Health, USA)分析每株植物总叶面积,以每株植物总叶面积/总叶干质量计算得出比叶面积。
采用单因素方差分析法(One-way ANOVA)比较同一器官、不同积雪梯度间的生物量、养分含量、生态化学计量学的差异,若方差为齐性,用LSD法进行显著性多重比较,若方差为非齐性,则用Tamhane′s 2法进行多重比较。显著性水平均为α=0.05,图表中数据为平均值±标准误。所有数据采用Origin2018、SPSS 20.0、Excel 2010进行计算与统计分析。
2 结果与分析
2.1 不同积雪梯度对钝苞雪莲各器官生物量的影响
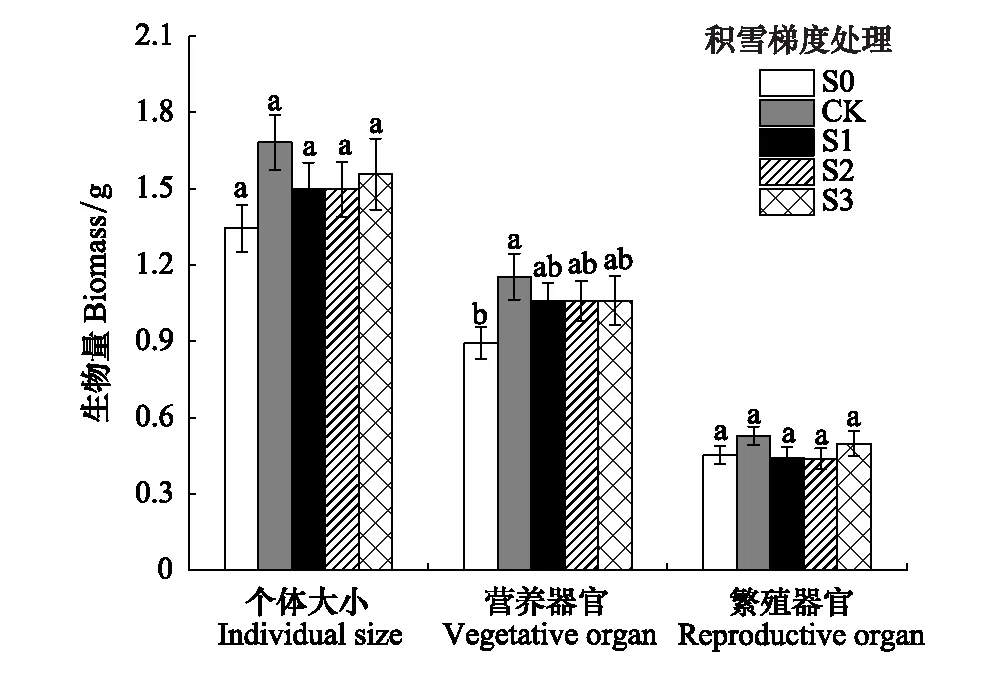
图2 不同积雪梯度间钝苞雪莲个体大小(地上部分)、营养器官和繁殖器官生物量差异Fig.2 Differences in individual size (aboveground biomass), vegetative organ biomass and reproductive organ biomass of Saussurea nigrescens under different levels of snowpackS0为去除积雪,CK为自然积雪量,S1、S2和S3分别为自然积雪量的2倍、3倍和4倍
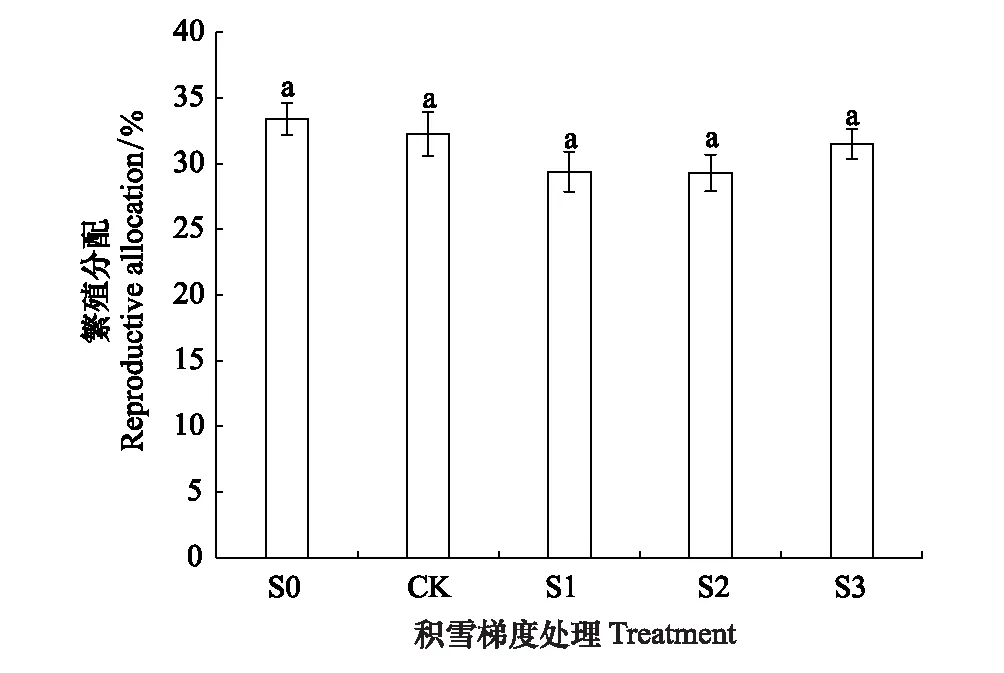
图3 不同积雪梯度间钝苞雪莲繁殖分配的差异 Fig.3 Differences in reproductive allocation of Saussurea nigrescens under different levels of snowpack
由图2可知,钝苞雪莲个体大小、繁殖器官生物量、营养器官生物量最大值均出现在CK(自然积雪量)处理下,另外个体大小、营养器官生物量在S0(去除积雪)处理下最低,但各处理间差异基本不显著,仅在CK处理下营养器官生物量显著高于S0处理(P<0.05)。由图3可知,繁殖分配随积雪梯度的增加先降低后上升,其中S0最高为33.4%,S2最低为29.3%,但各处理间无显著差异(P>0.05)。
2.2 繁殖器官生物量与个体大小及营养器官间的相互关系
不同积雪梯度处理下,随着个体大小(地上部分生物量)的增加,钝苞雪莲繁殖器官生物量显著增加,二者呈现极显著正相关关系(P<0.01),且去除积雪(S0)和增雪(S1—S3)处理的个体大小与繁殖器官生物量间的斜率大于CK。但繁殖分配与个体大小无显著相关性(图4)。
不同积雪梯度处理下(CK除外),钝苞雪莲营养器官和繁殖器官生物量同样表现出显著正相关,尤其是在4倍积雪时(S3处理),斜率达到0.45,高于其它积雪处理,表明更高积雪量可能导致更多资源分配至繁殖器官(图5)。

图4 钝苞雪莲个体大小(地上部分生物量)与繁殖器官生物量、繁殖分配的线性回归关系Fig.4 Linear regression relationships between individual size (aboveground biomass) and reproductive organ biomass, reproductive allocation of Saussurea nigrescens
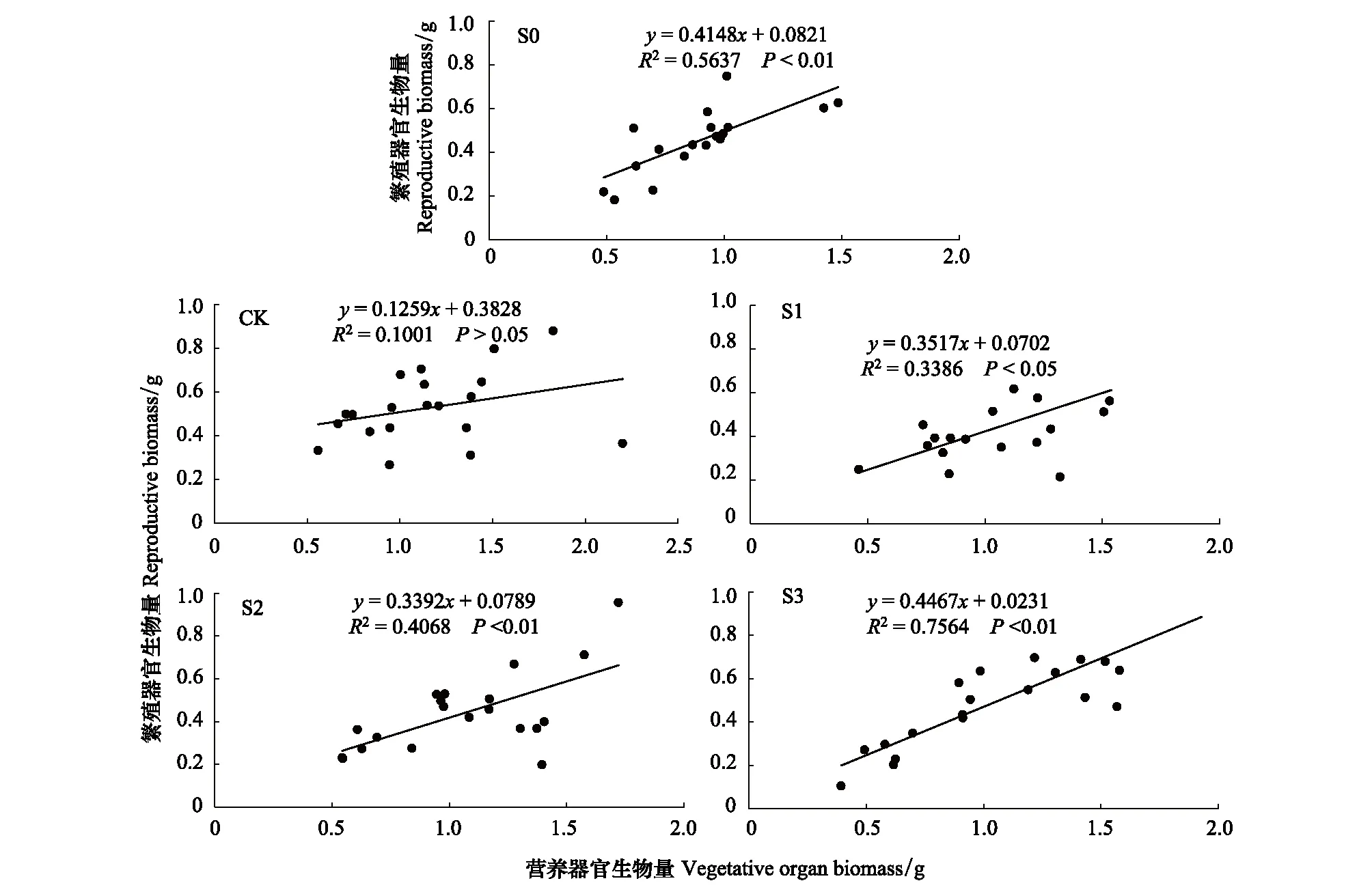
图5 钝苞雪莲营养器官生物量与繁殖器官生物量间的线性回归关系Fig.5 Linear regression relationship between vegetative organ biomass and reproductive organ biomass ofSaussurea nigrescens
2.3 不同积雪处理下各器官C、N、P化学计量特征
不同积雪梯度下的钝苞雪莲茎、叶、果实等器官养分含量存在显著差异。其中,茎部C含量在S3处理下显著高于S0(P<0.05),N含量在S0、S3处理下显著高于CK和S1,P含量随积雪梯度的增加显著升高(P<0.05)。叶片P含量随积雪梯度呈现单峰型变化,在S2处理达到峰值,且显著高于其它积雪处理(P<0.05);C、N含量则在各处理间无显著差异(P>0.05)。CK、S3处理下花苞C含量显著高于S0、S2,CK、S2、S3处理下花苞N含量显著高于S0、S1,花苞P含量在S1最高、在S2最低,且组间存在显著差异(表3)。
对照(CK)样方内钝苞雪莲茎部C∶N显著高于S0、S2、S3,茎部C∶P和N∶P表现为随积雪量的增加而显著降低(P<0.05)。对照(CK)样方内钝苞雪莲叶片C∶N显著高于S1、S2、S3(P<0.05),叶片C∶P和N∶P则表现为随积雪量的增加先下降后上升的趋势,即:S0>S3>CK>S1>S2。花苞C∶N在S0处理下最大且显著高于其它积雪梯度处理,C∶P和N∶P则在S2处理下最大且显著高于其它积雪梯度处理(表4)。
2.4 不同积雪处理下各器官功能属性特征
对照处理(CK)下,茎部干重最高且显著高于S0(P<0.05),但茎粗在不同积雪梯度下无显著差异;株高、茎高表现为随积雪量的增加先上升后下降,在CK处理下达到最大值且显著高于S0和S3(P<0.05);茎分配中S0处理最低且显著低于CK、S1、S2(P<0.05)(表5)。
S2处理下比叶面积最高且显著高于S0,单株叶片数中CK处理下最高且显著高于S2,叶干重与叶分配则在不同处理下均无显著差异(表6)。
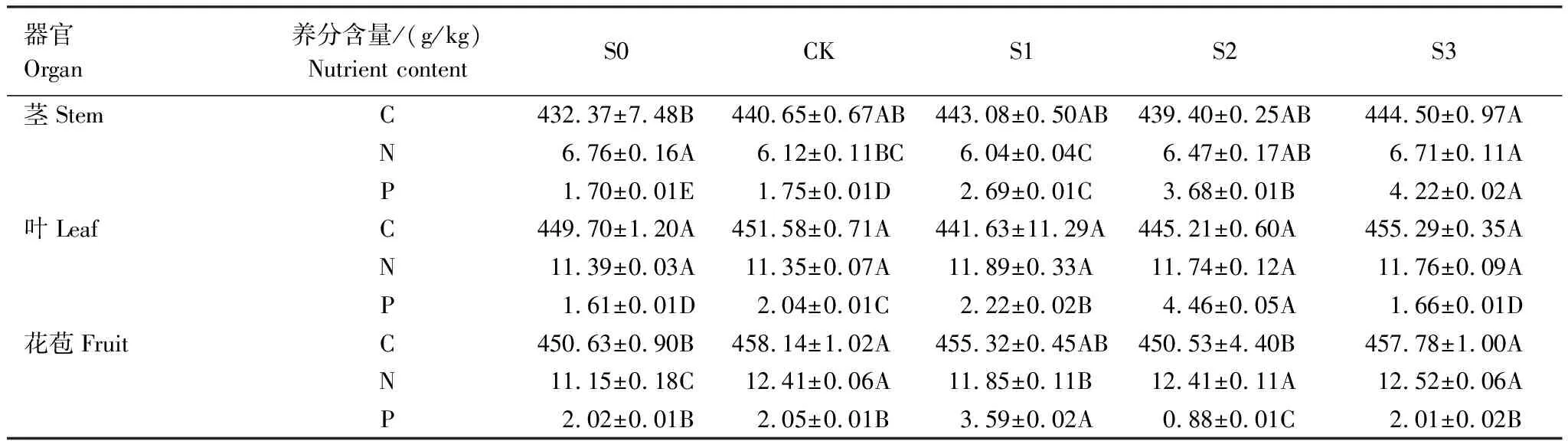
表3 不同积雪梯度下钝苞雪莲各器官养分含量

表4 不同积雪梯度下钝苞雪莲各器官生态化学计量特征
花苞直径和单株花苞重在组间无显著差异,花苞长度中CK最高且显著高于S1、S3,单株花苞数中CK最高且显著高于S0,单颗花苞重中S0最高且显著高于其余所有处理(表7)。
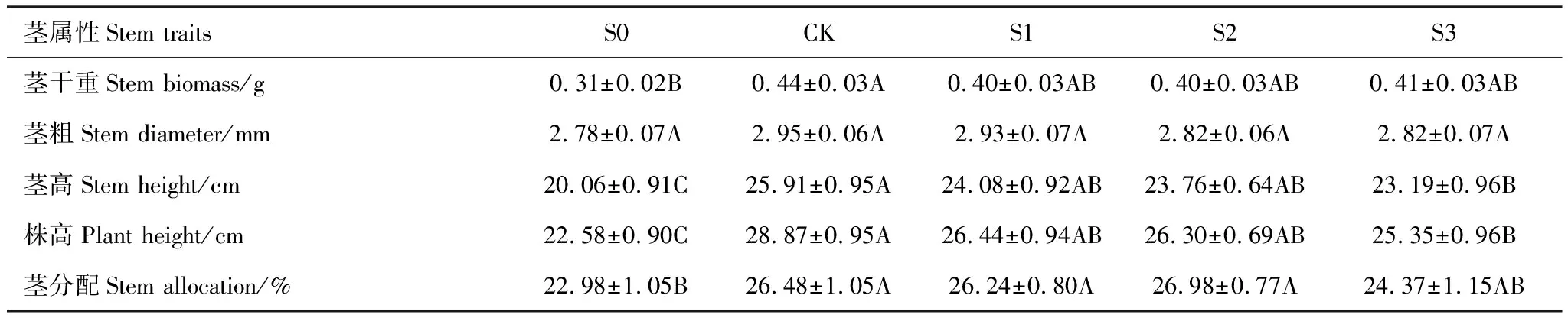
表5 不同积雪梯度下钝苞雪莲茎部属性
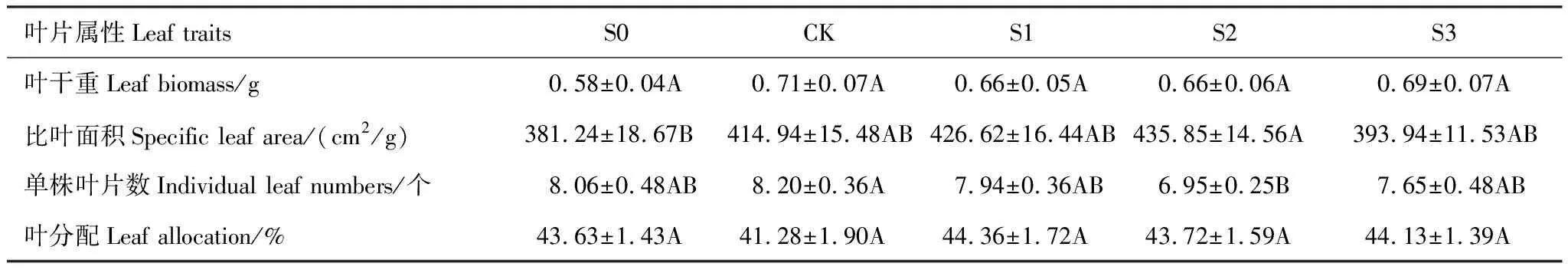
表6 不同积雪梯度下钝苞雪莲叶片属性
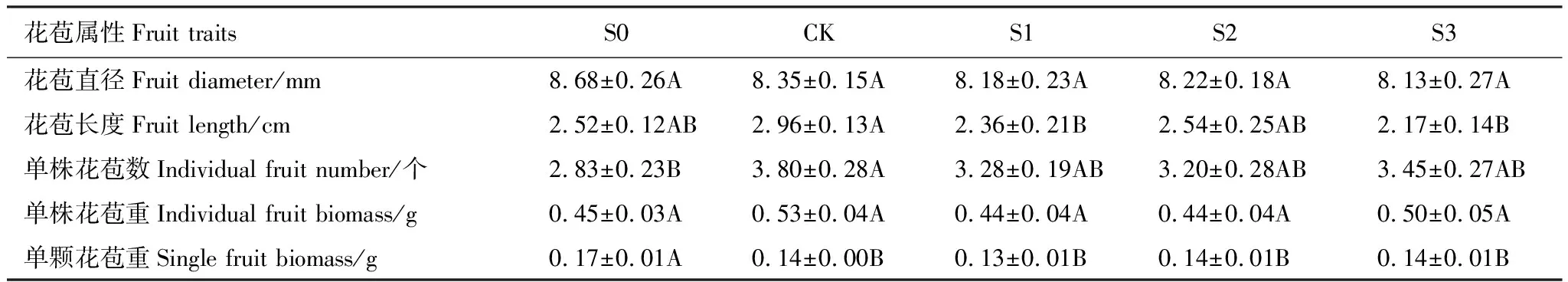
表7 不同积雪梯度下钝苞雪莲花苞属性
3 讨论
3.1 钝苞雪莲各器官生物量及繁殖分配特征
环境是植物生长发育的基础和必要条件,在青藏高原高寒草甸这样一个独特的生态系统中,植物往往要经受住降雪、霜冻、冰雹、昼夜温差大、太阳辐射强、气压低等极端环境的考验[1,40]。因此,生长在该环境下的钝苞雪莲也有着特殊的组织结构和生理特征,并对不同积雪厚度变化产生生态适应性[2]。本研究中,积雪处理对钝苞雪莲各器官生物量无显著影响,仅在CK处理下营养器官生物量显著高于S0(P<0.05)。造成这种差异不显著的原因可能是研究区降雪每年仅持续到4月底,而植物则在5月初返青、7月和8月达到生长旺盛期,故环境胁迫无法长期直接作用于植物生长,且冬季降雪对土壤温湿度的影响在生长季也有所降低,而夏季频繁的自然降雨更是削弱了这种影响[41],导致钝苞雪莲生物量对积雪变化的响应较弱。
但各器官的变化规律却相同,即积雪增加或减少处理下,钝苞雪莲地上部分生物量、繁殖器官和营养器官生物量均小于自然降雪,表明积雪量的改变对植物的生长发育起到了消极作用。这是因为冬季青藏高原具有昼夜温差大、热辐射水平高等特点[42],使得地下冰层反复冻融并在该过程中逐渐下移,且不同梯度下的地下冰储量对应的反复冻融所持续的时间和冰层下移的速率均不同[24]。因此,过多的积雪量(S1、S2、S3)导致地下冻土层反复冻融[24],这种强烈反复的冻融现象对植被根系的生长和存活产生消极的影响[43-44],这进一步负反馈到植物体地上部分[45]。而积雪去除(S0)直接导致土壤水分严重缺失,增加了水分胁迫,抑制植物地上部分的生长[46],尤其降低了营养器官生物量与茎干重(P<0.05),同时王辰阳等[47]研究也指出,土壤缺水可能会导致植物各茎节间的活动受阻,生长迟缓、株高降低,因此S0处理下各器官生物量低于CK处理。
不同处理间繁殖分配差异不显著(P>0.05),可能是研究样地内的积雪量变化对整个区域的传粉昆虫丰富度、活动能力并无影响,因此植物无需增加对繁殖器官的资源投入也可保障受粉结实的成功率[1]。但繁殖分配比例最高的为S0(图3),这是由于移除积雪导致土壤水分的大幅度减少,更加严峻的环境和繁殖压力迫使钝苞雪莲加大了对繁殖器官,如花苞的资源投入[1-2,48]。
3.2 钝苞雪莲各部位生物量间线性关系
有研究表明,植物个体生物量与繁殖器官生物量呈正比,与繁殖分配呈反比[2-3,8,49-50]。在本研究中,不同积雪处理下钝苞雪莲繁殖器官生物量随植物个体生物量的增加而增加(P<0.05),但繁殖分配与个体大小未呈现显著相关性(图4),这表明钝苞雪莲应对积雪量变化时并不依赖繁殖分配,可能是因为:1)钝苞雪莲的繁殖期为高寒草甸生长季,外界环境资源状况良好,积雪量变化的影响逐渐减弱,因此钝苞雪莲的繁殖代价较低,或在采样期间植物个体的物候差异较小导致[2-3];2)钝苞雪莲在面临环境胁迫时可通过性器官变异、提高种子成熟率来保证后代的成功繁衍[51],另外较低的繁殖分配投入可提高植物的资源吸收能力[52],因此繁殖分配可表现为不随个体大小的变化而显著改变;3)对于一个有限的资源库来说,对繁殖活动投入的增加意味着对营养活动投入的减少,将导致资源获取能力下降影响个体的生长,而植物为避免这种不利影响也会在自身个体增大时维持繁殖分配相对恒定[3]。
同时,图4显示在个体大小与繁殖器官生物量的关系中,CK处理下的斜率是0.2,而S0—S3的斜率在0.31—0.34之间(P<0.05),图5显示CK处理下的营养器官与繁殖器官间的斜率是0.1(P>0.05),S0—S3的斜率为0.33—0.44(P<0.05),这表明积雪处理在一定程度上改变了钝苞雪莲对生物量的分配,使繁殖器官获得比叶和茎更多的体内资源,提高了繁殖的绝对投入以支持种子成熟,这可能是环境变化改变了植物体内原有的生物量分配格局[53],当积雪增加时,土壤含水量上升,植物获取的有效水资源与自身组织的含水量也提高,可在一定程度上改变体内的光合生理特征与水分生理特征,此时植物可能具有更高的光合速率并能加大对繁殖器官的绝对投入[54],因此植物通过生物量投资及分配格局调节形成与水分资源供应水平相适应的生理特性,尤其是通过生长与繁殖之间的权衡维持自身发育的持久性与适应环境的稳定性[55]。而图3则表明增雪处理下的繁殖相对投入有下降趋势,可能因为随着繁殖器官的增多,植物相应需要产生更多的茎和叶才能支撑不断增大的繁殖结构[56]。
3.3 钝苞雪莲养分含量特征
生态化学计量学的动态平衡理论指出,有机体能够控制其自身的养分平衡使其在外界自然环境发生剧烈变化时仍能维持相对稳定[2,57],而碳、氮、磷作为生物有机体生长发育和繁殖过程中所必需的结构与营养元素,尽管其含量在同一物种的不同生长阶段表现出一定的波动性和变异性,但三者之间的化学计量特征可保持相对稳定,并能在一定程度上反映出植物碳的累积和氮、磷养分的限制格局,成为影响植物生长繁殖阶段的内在驱动因素之一[2,58-59]。在本研究中,不同积雪处理下钝苞雪莲茎、叶和花苞部位的碳、氮、磷含量及生态化学计量特征尽管出现了不同程度的差异,但无明显变化规律。其中,叶片N含量普遍低于我国植物叶片N含量平均值20.2g/kg[60],且除S2处理下花苞N∶P为14.16外,各处理下的茎、叶和花苞部位N∶P均位于1.59—7.08之间,表明钝苞雪莲受到N元素的限制[57],这也与宾振钧等[61]得出的青藏高原高寒草甸更易受到N元素的限制这一结论相符合。
3.4 钝苞雪莲各器官属性特征
植物不同器官的功能性状往往存在着相互作用关系,植物通常会根据外界环境变化和资源状况来权衡自身不同器官之间的资源分配,以达到对环境的最佳适应[2,62],短时期内的这种权衡关系可以通过植物的表型可塑性直接体现出来[63]。在本研究中,积雪量变化导致钝苞雪莲的茎、叶和花苞属性均表现出不同程度的差异。如去除积雪后,钝苞雪莲叶片生物量和比叶面积最低,这是由于积雪当量的水分减少导致干旱胁迫加剧,钝苞雪莲需要通过减少叶片生物量和降低蒸腾作用以维持体内水分[2,64],同时低的比叶面积意味着叶片中大部分物质被投入到增加叶肉细胞和构建保卫结构中去,使叶边缘更厚、叶组织密度更大[2,65];S2处理下植物的比叶面积最大,在一定程度上表明植物叶片同化二氧化碳和累积有机物质的能力更强[66-67],因此具有更高的生长速率。5个处理下,茎干重仅在CK下显著高于S0(P<0.05),叶干重在不同处理间无显著差异(P>0.05),但两者变化趋势与个体大小相同,表明茎、叶质量随个体大小的增大而增大,主要是因为较大的个体受到的外界干扰更强(如:动物践踏和采食、强风袭击等),因此个体大的植株其繁殖代价也增大,进而需要加大对繁殖支持结构的资源分配所致[15]。另外,单株花苞重在处理间无差异,可能钝苞雪莲对环境具有较强的适应性[51],并在研究区属优势种[38],因此在面临不同环境压力时具备一定抗性从而使其繁殖成本维持相对恒定。同时,花大小及数目一般与个体大小呈正相关关系[68],这与本研究中的CK处理下个体大小、单株花苞量、单株花苞重、花苞长度最大,S0处理下个体大小、单株花苞量最小相符合。此外,花苞数量在很大程度上表征着植物的繁殖适合度,花苞数目越多,植物产生种子数量越多,繁殖成功率越高,也就具有更高的繁殖适合度[2,68]。本研究中S0处理下花苞数目显著低于CK,故去除积雪可降低植物的繁殖适合度,也表明环境变化可能影响该物种的生存能力或分布,因此钝苞雪莲的生活史特征看起来已经适应了本地原有的生境,未来的积雪改变有可能对其繁殖带来胁迫。
川西北高寒草甸优势物种钝苞雪莲通过改变自身生物量、养分分配以及各器官功能性状,共同反映了其对于不同积雪梯度生境下的适应策略。但整体来看积雪变化对植物影响较弱,可能是因为积雪期与植物生长期独立,因此积雪对于正在生长的植物其直接作用较弱。已有研究表明,在功能群水平,同一功能群内的植物往往会体现出相似的属性集合,因此这些植物对于干扰的响应会趋于一致[69-70];而在群落水平,不同功能群植物的功能属性差异较大,这使得群落在面对外界环境变化或干扰时,不同功能群的生态位互补效应具有一定的缓冲作用[2,71-73],最终导致群落水平无显著变化。因此,本研究更加注重物种水平,尤其是优势种不同器官功能属性对外界环境变化的响应,但本研究仅初步探讨了一个优势物种的生态适应性,且只选取了容易快速测量的性状,未涉及更能准确代表植物对外界环境变化响应的性状。未来的研究需要综合更多植物物种的其它功能属性,如地下根系生物量和分泌物、土壤生物酶活性、叶片光合特征和水分利用效率、种子百粒重和繁殖成功率等指标,进一步研究植物个体乃至群落水平在不同积雪量变化下的生长繁殖分配特征、养分利用策略和生理生态适应机制,为区域及全球尺度下植物功能属性的模型研究提供基础数据。
4 结论
(1)积雪变化对钝苞雪莲的繁殖分配、个体大小以及繁殖器官生物量无显著影响。(2)钝苞雪莲不存在繁殖分配与个体大小间的依赖关系。(3)总体上繁殖器官生物量与个体大小、营养器官生物量之间呈显著或极显著的正线性关系。(4)积雪的增加显著提高了钝苞雪莲茎部P含量;而叶部P含量则具有先增后减的趋势(P<0.05)。(5)不同积雪处理对钝苞雪莲各器官的功能属性产生了不同影响,其中去除积雪(S0)降低了营养器官生物量,以及茎干重、株高、茎高、茎分配和单株花苞量,过度积雪(S3)则降低了株高、茎高和花苞长度。因此,积雪变化可改变钝苞雪莲不同器官的功能属性与养分含量,但这种变化对该物种的生存策略有何影响尚需要进一步的研究。
