亚热带森林不同恢复模式土壤与林下植被特征的动态研究
2021-10-28王振宇傅彦榕邹秉章苏香萍王思荣万晓华
王振宇 傅彦榕 邹秉章 苏香萍 王思荣 万晓华

萬晓华,1987年生,高级实验师,主要从事森林土壤碳氮循环研究,熟练操作总有机碳分析仪、连续流动分析仪、土壤粒径分析系统、全自动微生物鉴定系统和气相色谱质谱联用仪等多个大型仪器。2017-2019年主持国家自然科学基金青年项目(31600495)“根系在凋落物层生长对凋落物分解的影响及机制”,参与国家自然科学基金委重点项目(31930077)、国家自然科学基金委杰青项目(31232212)等课题,以第一作者发表论文或通讯作者发表论文11篇,其中Global Ecology and Biogeography、Soil Biology & Biochemistry、Plant and Soil等SCI一区期刊论文4篇、《生态学报》、《土壤学报》和《应用生态学报》等中文期刊5篇。
摘 要:选择亚热带自然恢复(次生林)和人工种植杉木两种恢复模式下不同林龄(5、8、21、27、40年)的林分为研究对象,通过测定0~10 cm土层土壤性质以及林下植被特征,探讨不同恢复模式下森林生态系统林下植被与土壤性质的恢复趋势以及相互作用过程。结果表明:随着林分发育,杉木林和次生林土壤碳氮储量均未发生显著变化,土壤C∶N保持不变。林型对土壤碳氮储量有显著影响,在林龄5年和8年,次生林土壤碳氮储量显著高于杉木林。随着林分发育,杉木林土壤磷储量呈增加-降低-增加的变化趋势,在杉木成熟期27年达到最低,C∶P和N∶P最大。而次生林在发育过程中,土壤磷储量先下降后增加,在21年达到最低。100年次生林表层土壤碳氮储量与40年次生林相当,而磷储量却处于较低水平。相关分析表明,杉木林中灌木层盖度与土壤碳氮储量呈显著正相关。在林分发育5年和8年,杉木林灌木层盖度和丰富度均显著低于次生林,但随着时间的推移在27年时趋于一致。研究表明,杉木林在短期恢复中更有利于地上生物量的快速恢复,其经营措施增加了草本层的丰富度,但以牺牲林分发育前期灌木层的盖度和丰富度为代价;自然恢复能更快地形成复杂结构的森林群落。杉木营林过程中,可减少对林下灌木层植被的砍伐,保持一定的林下植被发育水平有利于土壤地力条件的改善和恢复。
关键词:森林恢复;次生林;杉木人工林;土壤理化性质;林下植被
中图分类号:S 714.2 文献标志码:A 文章编号:0253-2301(2021)08-0007-10
DOI: 10.13651/j.cnki.fjnykj.2021.08.002
Dynamics Study on Soil and the Characteristics of Understory Vegetation UnderDifferent Restoration Modes in Subtropical Forests
WANG Zhen-yu1,2, FU Yan-rong1,2, ZOU Bing-zhang3, SU Xiang-ping1,2, WANG Si-rong3, WAN Xiao-hua1,2*
(1. School of Geographical Science, Fujian Normal University, Fuzhou, Fujian 350007, China;
2. Cultivation Base of State Key Laboratory of Humid Subtropical Mountain Ecology, Fuzhou, Fujian 350007, China;
3. Fujian Shanghang Baisha State-owned Forest Farm, Longyan, Fujian 364205, China)
Abstract: The standing forests with different ages (5, 8, 21, 27, and 40 years) under two restoration modes of subtropical natural restoration (secondary forest) and artificial plantation of Cunninghamia lanceolata were selected as the research objects. By measuring the soil properties and the characteristics of understory vegetation in the soil layer of 0-10 cm, the restoration trend and interaction process of the understory vegetation and the soil properties in different restoration modes were discussed. The results showed that with the development of the stand, the carbon and nitrogen storage in soil of Cunninghamia lanceolata forest and secondary forest did not change significantly, and the soil carbon-nitrogen ratio remained unchanged. Forest type had a significant effect on the soil carbon and nitrogen storage, and the soil carbon and nitrogen storage in the secondary forest was significantly higher than that in Cunninghamia lanceolata forest at 5 and 8 years of stand age. With the development of the stand, the soil phosphorus storage of Cunninghamia lanceolata forest showed a trend of increasing-decreasing-increasing, and reached the lowest at 27 years of Cunninghamia lanceolata mature period, while the carbon-phosphorus ratio and nitrogen-phosphorus ratio reached the highest. However, during the development of the secondary forest, the soil phosphorus storage decreased first and then increased, and reached the lowest at 21 years of stand age. The carbon and nitrogen storage in the surface soil of the 100-year secondary forest was similar to that of the 40-year secondary forest, but the phosphorus storage was at a lower level. The correlation analysis showed that the coverage of shrub layer was significantly positively correlated with the soil carbon and nitrogen storage in Cunninghamia lanceolata forest. At 5 and 8 years of stand development, the coverage and abundance of shrub layer in Cunninghamia lanceolata forest were significantly lower than those in the secondary forest, but as time went on they tended to be the same at 27 years of stand age. The results showed that Cunninghamia lanceolata forest was more conducive to the rapid recovery of above-ground biomass in the short-term recovery, and its management measures increased the abundance of herbaceous layer. However, the coverage and abundance of shrub layer were sacrificed at the early stage of stand development. The natural restoration could form the complex forest communities more quickly. In the culture and management process of Cunninghamia lanceolata forest, the felling of vegetation in the understory shrub layer could be reduced, and maintaining a certain level of understory vegetation was beneficial to the improvement and restoration of the soil fertility conditions.
Key words: Forest restoration; Secondary forest; Cunninghamia lanceolata plantation; Soil physical and chemical properties; Understory vegetation
自森林面积减少造成严重的环境危机以来,森林恢复日益受到各国政府和生态学家的关注[1]。自然恢复与人工造林是全球森林恢复的两种主要模式[2-3]。有研究指出,单一树种人工林大面积栽培可能会造成土壤退化、生物多样性减少以及生态服务功能丧失等负面问题[4-5]。而自然恢复的森林往往被认为具有较高的生态系统服务功能[6]。因此,对集约化栽植人工林和自然恢复次生林进行比较研究是理解人工林生态系统服务功能丧失原因的重要途径。
不同恢复模式对土壤特性和林下植被特征影响较大[7]。最近的综述研究表明,全球范围内,土壤碳氮储量在森林停止干扰后随时间的推移而增加,但恢复30年后仍低于参照水平[1];Feng等[8]在中国亚热带对恢复了31年的退化森林研究发现,人为干扰停止后,表层土壤碳、氮以及有效養分含量随着恢复时间呈现升高的趋势,而恢复时间对总磷含量无显著影响。Hume等[9]在北方森林研究发现,火灾干扰后的恢复过程中表层土壤氮含量呈先上升后下降的驼峰型趋势。Chen等[10]对亚热带杉木人工林的研究发现,土壤有机碳储量在森林皆伐再植后迅速下降,到16年时下降了38.9%。而Bautista-crus等
[11]在热带森林发现,农田退耕还林后的恢复过程中,表层土壤氮含量无明显变化,土壤磷含量随恢复时间增加。可见,不同地区和不同扰动类型下,土壤性质恢复模式差异较大。植被组成、气候条件、土地利用方式等对森林土壤性质均有重要影响[12]。此外,林下植被作为森林生态系统的重要组分,在维持森林生态系统结构和功能方面起着不可忽视的作用[13]。而在森林恢复过程中,一方面,灌木和草本植被需要不断调整分布格局以适应林分微环境(光照条件、养分资源等)在时空上的变化[14]。另一方面,林下植被的变化也必然反馈于土壤,改变土壤的生物化学过程进而影响着森林生态系统长期的稳定性和生产力
[15]。因此,比较人工经营和自然恢复下森林土壤和林下植被性质的恢复动态,有利于及时把握亚热带森林恢复现状,对重建和管理森林生态系统具有重要意义。
为缓解生态系统退化造成的环境危机,中国开展了世界上最大的生态恢复工程[3,16]。截至2010年,已将3.2×107 hm2的耕地或荒地转化为了森林和草地[17]。然而随着生态恢复的进行,不同恢复模式下土壤性质与林下植被的发育如何?地上地下的恢复速度以及能否恢复到干扰前的水平也是目前极为关心的问题[18]。鉴于此,本研究选取亚热带森林皆伐后自然恢复和人工种植杉木两种恢复模式不同林龄(5、8、21、27、40年)的林地作为研究平台,并以当地林龄大于100年的次生林作为参照,以期揭示两种恢复模式下森林土壤理化性质和林下植被的变化规律。为该地区森林恢复和重建提供依据。
1 研究地区与研究方法
1.1 研究区概况
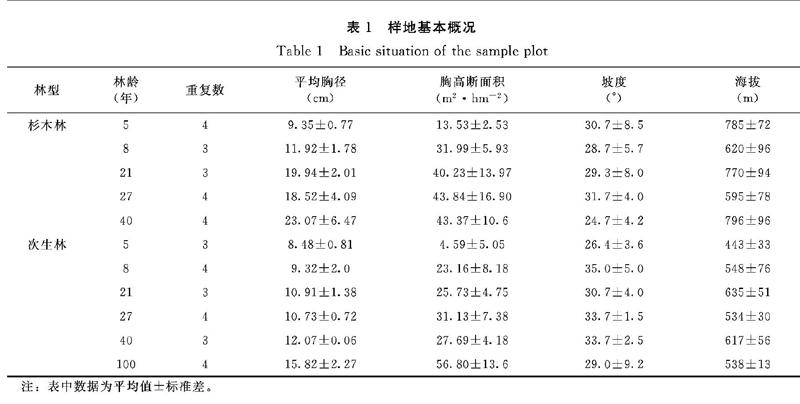
研究区地处武夷山脉南段低山丘陵地带(25°04′ N,116°30′ E),属亚热带季风气候,海拔400~800 m,山地坡度10°~40°,年均温度20.1℃,年降雨量1600 mm,无霜期270 d。土壤主要由粗粒花岗岩发育而成的红壤组成,土层厚度60 cm以上。该地区同时具有皆伐后自然恢复的次生林和集约化种植杉木的人工林,恢复时间均超过40年,且存在多种不同时间段的植被类型[19]。研究样地均经历了相同程度的皆伐,前茬均为成熟的常绿阔叶林。根据查看林业历史资料,定位了100年来未受人为干扰的次生林作为参照。
杉木林下灌木层植物主要包括弯蒴杜鹃Rhododendron henryi、黄瑞木Adinandramillettii、日本五月茶Antidesmajaponicum、密花山矾Symplocoscongesta等;草本层植物主要包括狗脊蕨Woodwardiajaponica、算盘子Glochidionpuberum、扇叶铁线蕨Adiantumflabellulatum、石斑木Rhaphiolepisindica等。次生林林下灌木层植物主要包括黄瑞木Adinandramillettii、狗骨柴Diplosporadubia、榕叶冬青Ilex ficoidea、变叶榕Ficus variolosa等;草本层植物主要包括粉背菝葜Smilax hypoglauca、狗脊蕨Woodwardia japonica、算盘子Glochidionpuberum。
1.2 试验设计
2018年7月,选取5个林龄(5、8、21、27、40年)的杉木人工林和天然次生林,并以林龄大于100年的次生林作为当地森林演替的参照值。每个林龄设置4个重复,但由于样地的局限性和实际条件,部分年龄阶段设置3个重复,共39个样地,每一个样地的面积为20 m×30 m。样地之间间隔大于500 m。距离农田、道路或其他类型林地的边缘大于100 m,所有样地均处中坡位。样地基本特征见表1。
于2019年7月份,用内径为3.5 cm的土钻在每个样方内沿“S”形选取12个取样点,去除凋落物层,取0~10 cm土壤样品。按照四分法,在每个样方内不同点取得的土样汇集成复合样品,挑除石砾和植物细根残体并分为两份,一份过2 mm筛网,用于测定土壤pH值以及有效养分含量。另一份在自然状态下风干后研磨过0.15 mm筛网,用于测定总碳、总氮、总磷指标。在样方的中心区域挖取土壤剖面,用环刀取0~10 cm原状土,用于测定土壤容重及含水量。
在样方内,对胸径≥ 5 cm的所有活树进行鉴定记录并计算胸高断面积。灌木层被定义为高度在1.3~4.0 m的所有木本物种,草本层为高度<1.3 m的维管和非维管植物[19]。在每个样地中,从样地中心以放射状向不同的方向和距离(不重叠)随机选取3个5 m×5 m和10个1 m×1 m的样方,分别调查灌木层和草本层物种数量,并按照Mueller-Dombois等[20]的方法,测定灌木层和草本层的盖度。
1.3 测定项目与方法
土壤容重和含水量用烘干法测定;土壤pH采用电位法(土水比为1∶2.5)测定;土壤总碳、总氮采用元素分析仪(Elemental EL MAX CNS analyzer,德国)测定;土壤总磷的测定参考Carter等[21]的方法,样品经H2SO4-HClO4(4∶1)高温消解,再用连续流动分析仪(Skalar San++,荷兰)测定磷酸根离子含量。土壤有效磷和矿质氮采用连续流动分析仪(Skalar San++,荷兰)测定。
1.4 数据处理
用一般线性模型和方差分析(ANOVA)检验林龄和林型对土壤理化性质和林下植被特征的影响,用shapiro-wilk′s test对残差值进行正态性检验(α=0.05)。采用皮尔逊相关分析(Pearson)方法检验林下植被特征与土壤理化性质的关系。用R进行分析作图。
2 结果与分析
2.1 不同林型土壤理化性质的变化趋势
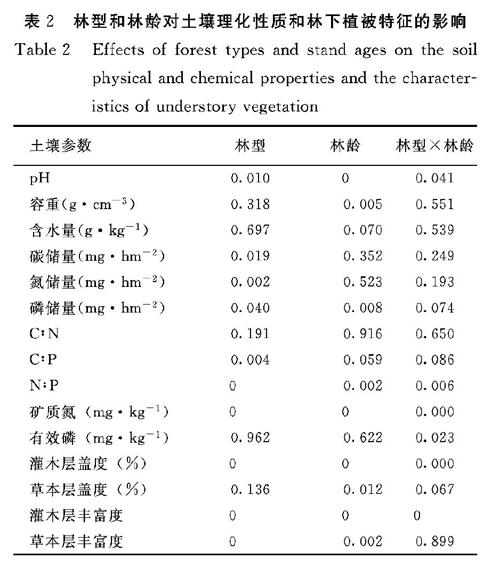
从表2可知,林龄、林型以及交互作用对0~10 cm土层土壤pH均有显著影响。从图1可知,杉木人工林土壤pH在27年之前呈下降趋势。与恢复了40年的杉木人工林和天然次生林相比,100年的次生林表土层土壤pH值最低。林龄和林型对表层土壤含水量均无显著影响,杉木林和次生林在发育过程中,土壤含水量呈升高的趋势。林龄对土壤容重有显著影响,100年次生林表层土壤容重处于较低水平。
从表2可知,林龄对土壤碳氮储量均无显著影响,而林型对土壤碳氮储量有显著影响。从图2可知,在林分发育5和8年,次生林表层土壤碳氮储量均显著高于杉木人工林,在杉木林的速生阶段8~27年,土壤磷储量呈下降的趋势。而次生林在发育过程中,土壤磷储量先下降后增加在21年达到最低。与发育40年时的杉木林和次生林相比,100年次生林表层土壤碳氮储量与之相当,而磷储量却处于较低水平。
从表2可知,林型和林龄对土壤有效磷含量均无显著影响,林型和林龄以及交互作用对土壤矿质氮均有显著影响。从图2可知,林分发育40年,杉木人工林在各个林龄土壤矿质氮含量均高于天然次生林。但杉木人工林矿质氮含量无明显变化趋势,而天然次生林土壤矿质氮含量呈增加的趋势。恢复了40年的次生林土壤矿质氮含量仍显著低于100年的水平。
从表2可知,林型和林龄对土壤C∶N均无显著影响,而林型对土壤C∶P和N∶P有显著影响。从图3可知,与发育了40年的杉木林和次生林相比,100年次生林的C∶P和N∶P均处于较高水平。在杉木林的生长过程中,土壤C∶P和N∶P在8~27年速生阶段呈增加的趋势,而次生林呈先增加后降低的趋势,均在21年达到最高水平。
2.2 不同林型林下植被的变化趋势
从表2可知,林型和林龄对灌木层丰富度和盖度均有显著影响。从图4可知,在林分发育的5和8年时间段,杉木林灌木层植被盖度和丰富度显著小于天然次生林。但在40年的恢复过程中,杉木人工林灌木层植被盖度随林龄的增加而增加,在27年阶段与次生林趋同,并达到与100年次生林同样水平;杉木人工林的草本层植被的丰富度在林分发育的各个阶段均显著高于天然次生林。
2.3 不同林型林下植被特征与土壤理化性质的关系
从表3可知,杉木人工林中,灌木层植被盖度与土壤碳氮储量均呈显著正相关关系(P<0.05),而草本层丰富度与土壤pH和土壤容重呈显著正相关关系,与土壤含水量呈显著负相关关系。而在次生林中,灌木层植被盖度与土壤氮储量呈显著正相关(P<0.05),而灌木层丰富度与土壤有效磷呈显著负相关关系(P<0.05)。
3 讨论
3.1 杉木林和次生林土壤理化性質的恢复趋势
在森林生态系统中,林龄通过改变植被凋落物质量[22]、细根生物量与周转速率[23],以及林分结构等因素,进而影响着土壤养分分配格局。本研究发现,经过40年的发育,不同恢复模式表层土壤pH、容重随林龄发生了显著变化。杉木人工林中,土壤pH呈先降后增的趋势,在杉木成熟期27年龄段达到最低水平。这与张芸等[24]在不同林龄杉木林的研究结果一致。土壤pH在很大程度上取决于土壤酸性离子(H+、Al3+)和碱性阳离子(Ca2+、Mg2+)的分布状况[25]。在生态系统速生阶段,一方面,植物由于快速吸收养分,从根系中不断释放出H+,而土壤中基本的碱性阳离子如钙镁(Ca2+、Mg2+)等逐渐固定到植被生物量中,从而导致土壤溶液H+浓度升高,造成土壤酸化[26]。另一方面,凋落物分解过程中产生的有机酸也是土壤酸化的重要原因,杉木凋落物木质素含量较高,形成的腐殖质具有较低分子质量的有机酸,而这种有机酸容易被水浸出,导致土壤酸度增加[27],使人工林在速生阶段土壤pH显著降低。
森林生态系统中的碳大约有2/3储存在土壤,土壤碳动态可能对生态系统碳平衡产生重大影响[12],然而,土壤碳储量变化的驱动机制是非常复杂的。本研究表明,林龄对土壤碳储量无显著影响。尽管在一些研究中发现了土壤碳储量随着森林的发育而增加[8-9],但大量的研究也呈现出不同的结果。例如,一项关于热带次生林恢复的综述分析表明,在牧场或者农田上进行次生演替的森林土壤碳储量与林龄无显著相关关系[28]。而在亚热带杉木人工林中研究发现,土壤有机碳于森林皆伐再植后迅速下降,到16年时下降了38.9%,但在16~21年开始重新积累碳,而21年之后达到了比皆伐前低30%的土壤有机碳库并保持稳定[10]。这些研究所呈现出的不同结果表明,评估森林恢复过程中土壤碳储量的变化要综合考虑气候、土壤类型、管理措施以及植被组成等重要因素。本研究杉木生长在粗粒花岗岩发育的土壤上,土壤质地以砂质壤土为主,吸附和稳定有机碳的作用较差,不利于土壤有机碳的固持[24]。本研究发现,杉木人工林中土壤磷储量从8年中幼林阶段到杉木成熟期27年下降,并在成熟期达到最低水平,此外C∶P和N∶P在该阶段有上升趋势。这主要是由于磷素属于地球化学循环驱动的养分,其含量主要由土壤母质类型和风化程度所决定[29]。而杉木为低养分归还的针叶树种[30],养分循环周期较长。而且从生长周期来看,速生期和成熟期的杉木对磷等养分的需求量达到最大[31],导致土壤中的磷在杉木速生阶段逐渐被植被木质器官所存留。此外,本研究与亚热带森林演替结论一致[32],即顶级群落100年次生林胸高断面积达到最高水平,土壤容蓄能力较好(高含水量、低容重),矿质氮含量较高,但C∶P和N∶P均达到较高水平,表明该区域森林生态系统长期的发育使得磷限制加剧[8]。本研究发现,杉木人工林在幼龄林阶段表层土壤碳氮储量显著低于次生林,这可能是人工林栽植前的整地措施加速了有机残留物与土壤的混合,导致部分有机质被快速分解[33]。
3.2 杉木林和次生林土壤理化性質与林下植被的关系
本研究发现,杉木林在5年和8年幼龄林阶段,灌木层丰富度和盖度显著低于天然次生林,而草本层丰富度在各个阶段均高于次生林。这种差异可能与人工林频繁的人为干扰(如疏伐和施肥)增加了光照和土壤养分的有效性,从而促进了草本植物的生长有关[19]。一方面,疏伐往往会破坏灌木层盖度和丰富度,但疏伐后林下资源的可用性提高,为快速生长的草本植物(如一年生植被)提供有利的生长条件[34]。另一方面,天然次生林灌木盖度和丰富度较高,通过对光和养分的竞争,抑制了草本层的生长[14]。而随着林分的发育,在杉木林轮作期27年,灌木层盖度逐渐与次生林趋同,可能由于本研究杉木人都是一代人工林,仍然保留着前茬天然阔叶林的大量种子库,因此在人为干扰停止后,灌木层植被可快速恢复并达到次生林的水平[19]。但值得注意的是,经过多代轮作后,种子库可能会耗尽,灌木层再生速度也会减慢甚至停止。而灌木层作为林下植被的重要组分,其存在对生态系统功能的维持有着重要的作用。本研究中,杉木林下灌木层盖度与土壤碳氮储量均呈显著正相关关系。此外,也有研究表明,林下灌木植被去除显著降低了土壤有机碳、总氮、总磷、矿质氮等养分含量,同时也抑制了纤维素酶和亮氨酸基肽酶活性[13]。林下植被可以通过根系代谢活动,直接增加土壤中无机和有机养分含量[10]。同时,杉木凋落物与林下植被凋落物有明显的混合分解效应,当混合了30%的林下植被凋落物后,其分解速率可以提高1倍[35]。因此,林下植被的完整对森林的养分循环有重要的影响。在杉木林营林过程中,保持一定的林下植被发育水平,减少对灌木层植被的砍伐,或适当延长杉木人工林轮作期以促进灌木层植被的定植可能有利于土壤肥力的保持与恢复。
4 结论
研究表明,40年的恢复过程中,杉木林和次生林土壤碳氮储量均无显著变化。杉木林在人工抚育的短期恢复中胸高断面积快速增加,可能更有利于地上生物量的快速恢复;而自然恢复次生林能更快地形成草灌乔多层结构的森林群落。此外,灌木层盖度是影响杉木林土壤理化性质的重要因素,人工林经营措施增加了草本层的丰富度,但以牺牲林分发育前期灌木层的盖度和丰富度为代价。
参考文献:
[1]MORENO-MATEOS D,BARBIER E B,JONES P C,et al.Anthropogenic ecosystem disturbance and the recovery debt[J].Nature Communications,2017,8:14163.
[2]王芸,欧阳志云,郑华,等.不同森林恢复方式对我国南方红壤区土壤质量的影响[J].应用生态学报,2013,24(5):1335-1340.
[3]CHEN C,PARK T,WANG X,et al.China and India lead in greening of the world through land-use management[J].Nature Sustainability,2019,2(2):122-129.
[4]NEWBOLD T,HUDSON L N,HILL SLL,et al.Global effects of land use on local terrestrial biodiversity[J].Nature,2015,520(7545):45-50.
[5]SELVARAJ S,DURAISAMY V,HUANG Z,et al.Influence of long-term successive rotations and stand age of Chinese fir (Cunninghamia lanceolata) plantations on soil properties[J].Geoderma,2017,306:127-134.
[6]SEGURA C,NAVARRO F B,JIMNEZ M N,et al.Implications of afforestation vs.secondary succession for soil properties under a semiarid climate[J].Science of The Total Environment,2020,704:135393.
[7]ZHANG K,DANG H,TAN S,et al.Vegetation community and soil characteristics of abandoned agricultural land and pine plantation in the Qinling Mountains,China[J].Forest Ecology and Management,2010,259(10):2036-2047.
[8]FENG C,YAN M,FU S,et al.Soil carbon and nutrient dynamics following cessation of anthropogenic disturbances in degraded subtropical forests[J].Land Degradation & Development,2017,28:2457-2467.
[9]HUME A,CHEN HYH,TAYLOR AR,et al.Soil C∶N∶P dynamics during secondary succession following fire in the boreal forest of central Canada[J].Forest Ecology and Management,2016,369:1-9.
[10]CHEN GS,YANG ZJ,GAO R,et al.Carbon storage in a chronosequence of Chinese fir plantations in southern China[J].Forest Ecology and Management,2013,300:68-76.
[11]BAUTISTA-CRUZ A,DELCASTILLO R.Soil changes during secondary succession in a tropical montane cloud forest area[J].Soil Science Society of America Journal,2005,69:906-914.
[12]POWERS JS,MARN-SPIOTTA E.Ecosystem processes and biogeochemical cycles in secondary tropical forest succession[J].Annual Review of Ecology,Evolution, and Systematics,2017,48(1):497-519.
[13]刘仁,袁小兰,刘俏,等.林下植被去除对杉木人工林土壤酶活性及其化学计量比的影响[J].林业科学研究,2020,33(5):121-128.
[14]KUMAR P,CHEN HYH,THOMASSC,et al.Linking resource availability and heterogeneity to understorey species diversity through succession in boreal forest of Canada[J].Journal of Ecology,2018,106(3):1266-1276.
[15]POWERS R,BUSSE M,MCFARLANE K,et al.Long-term effects of silviculture on soil carbon storage:Does vegetation control make a difference?[J].Forestry,2012,86:133-139.
[16]ZHANG P,SHAO G,ZHAO G,et al.China′s forest policy for the 21st century[J].Science,2000,288(5474):2135.
[17]ZHANG KR,DANG H,TAN S,et al.Change in soil organic carbon following the′Grain-for-Green′programme in China[J].Land Degradation & Development,2010,21(1):13-23.
[18]刘斌,张参参,汪金松,等.江西九连山不同恢复模式林分的物种多样性特征[J].林业科学研究,2020,33(4):42-52.
[19]SU X P,LI S J,WAN X H,et al.Understory vegetation dynamics of Chinese fir plantations and natural secondary forests in subtropical China[J].Forest Ecology and Management,2020,483:118750.
[20]MUELLER-DOMBOIS D,ELLENBERGH.Aims and methods of vegetation ecology[M].Network:Wiley,1974.
[21]CARTER MR,GREGORICH EC.Soil sampling and methods of analysis[M].Florida:CRC Press,1993.
[22]FENG C,WANG Z,MA Y,et al.Increased litterfall contributes to carbon and nitrogen accumulation following cessation of anthropogenic disturbances in degraded forests[J].Forest Ecology and Management,2019,432:832-839.
[23]FENG C,WANG Z,QI Z,et al.Rapid increases in fine root biomass and production following cessation of anthropogenic disturbances in degraded forests[J].Land Degradation & Development,2018,29:461-470.
[24]張芸,李惠通,张辉,等.不同林龄杉木人工林土壤C∶N∶P化学计量特征及其与土壤理化性质的关系[J].生态学报,2019,39(7):1-11.
[25]TIAN D,NIU S.A global analysis of soil acidification caused by nitrogen addition[J].Environmental Research Letters,2015,10:179-186.
[26]YU ZP,SEARLE E,SARDANS J,et al.Whole soil acidification and base cation reduction across subtropical China[J].Geoderma,2019,361:667-676.
[27]KOVP,TEJNECKY V,ASH C,et al.Low-Molecular-Mass organic acids in the forest soil Environment[J].Mini-Reviews in Organic Chemistry,2017,14:75-84.
[28]MARIN-SPIOTTA E,SHARMA S.Carbon storage in successional and plantation forest soils:A tropical analysis[J].Global Ecology and Biogeography,2013,22:105-117.
[29]VITOUSEK PM,STEPHEN P,HOULTON BZ,et al.Terrestrial phosphorus limitation:mechanisms,implications,and nitrogen-phosphorus interactions[J].Ecological Applications,2010,20(1):5-15.
[30]盛煒彤,杨承栋,范少辉.杉木人工林的土壤性质变化[J].林业科学研究,2003,16(4):377-385.
[31]MA X Q,HEAL KV,LIU A,et al.Nutrient cycling and distribution in different-aged plantations of Chinese fir in southern China[J].Forest Ecology and Management,2007,243(1):61-74.
[32]LI Y,YANG F,OU Y,et al.Changes in forest soil properties in different successional stages in lower tropical China[J].PLOS ONE,2013,8(11):1359.
[33]ZHENG H,OUYANG ZY,WANG XK,et al.How different reforestation approaches affect red soil properties in southern China[J].Land Degradation & Development,2005,16:387-396.
[34]ROSSMAN A,HALPERN C,HARROD R,et al.Benefits of thinning and burning for understory diversity vary with spatial scale and time since treatment[J].Forest Ecology and Management,2018,419:58-78.
[35]盛炜彤.关于我国人工林长期生产力的保持[J].林业科学研究,2018,31(1):1-14.
(责任编辑:柯文辉)
收稿日期:2021-07-02
作者简介:王振宇,男,1996年生,硕士,主要从事森林土壤碳氮循环研究。
通信作者:万晓华,女,1987年生,博士,高级实验师,主要从事森林土壤碳氮循环研究(E-mail:xiaohuawan2012@foxmail.com)。
基金项目:国家自然科学基金项目(31570604、31600495)。
