闽北青冈林迹地演替林木群落物种多样性特征
2021-10-27陈杰
陈杰
(福建省建阳范桥国有林场, 福建 南平 354200)
福建省的壳斗科植物主要为青冈属(Cyclobalanopsis)、锥属(Castanopsis)、石栎属(Lithocarpus)及栗属(Castanea),为中国壳斗科植物密集的地区之一[1],在天然环境中由于内外在的持续扰动,顶级群落往往难以长期维持,而在次级演替到顶级间循环,适度的扰动可维持林分的稳定性,此类规律又称为中度干扰假说(Intermediate Disturbance Hypothesis)[2]。而强大的人为干扰和自然干扰是导致林分逆向演替,形成低质低效次生林的原因[3]。林分干扰后自我恢复和自我调控的研究是森林生态系统的一个热点[4]。本研究试验地位于福建省南平市延平区葫芦山村,原生优势群落以壳斗科乔木为主,如福建青冈(Cyclobalanopsischungii)、青冈(Cyclobalanopsisglauca)、甜槠(Castanopsiseyrei)、栲树(Castanopsisfargesii)等,周边有马尾松(Pinusmassoniana)林及木荷防火林带。20世纪50年代以来,经人工干扰后天然形成不同时期的青冈次生林。实验地选择天然更新30、35、40、50、65、75年的不同演替阶段林地6个,调查并分析林木群落物种多样性特征,根据其自然演替规律,探索青冈次生林自然恢复方式,为解决中国长期传统经营方式造成森林生态系统结构简单、病虫害严重、多样性降低、生态功能和稳定性下降等问题,构建健康稳定优质高效的森林生态系统[5]提供参考。
1 研究区域概况
研究区位于福建省南平市延平区葫芦山村,海拔在60~700 m之间,土壤以红壤为主,年均温为19.8℃,最热月(7月)均温为25.5℃,最冷月(1月)均温为9.1℃,年降雨量为1 700 mm。本研究区前茬是青冈林,早期因民生所需,采用择优采伐的“拔大毛”采伐方式,进行阶段性采伐利用,随着经济发展及产业结构调整,于20世纪90年代停止采伐,采伐后的林地经天然更新逐渐发育为不同阶段的青冈林,并有少数马尾松及木荷(Schimasuperba)镶嵌其中。
2 研究方法
2.1 样地设置及林木调查
选取采伐后不同天然更新时间的林地作为试验样地,其优势树种为福建青冈、青冈、栲树、木荷等,根据资源档案资料,分别在沙基洋(30、35年)、黄梨园(40、50年)及双坑口(65、75年)各设置2个50 m×50 m方形样区,最终依据天然更新时间编定为A区(30年)、B区(35年)、C区(40年)、D区(50年)、E区(65年)、F区(75年),每个样区内再划分25个10 m×10 m小样区。2019年进行样地调查,选择胸高直径1 cm以上乔木,调查树种、胸径、树高等。
2.2 数据处理与分析
本研究采用重要值(Important Value;IV)、α多样性测度、β多样性测度来比较不同演替阶段林木群落物种多样性特征变化,其中重要值可以用来表现物种在群落中的相对重要性,α多样性测度与β多样性测度一起构成了总体多样性(Overall Diversity)与生物异质性(Biotic Heterogeneity),可用来表现一定地段下的物种多样性梯度变化情形[6-8],α多样性测度可表现出不同演替阶段下各群落的总体多样性。本研究采用Margalef丰富度指数、Simpson 优势度指数、Shannon-Wiener 多样性指数及Pielou 均匀度指数。β多样性测度可用来表现演替阶段间的梯度变化。本研究采用Jaccard相似性指数、Sorenson相似性指数、Whittaker 相异性指数及Cody相异性指数。计算公式如下。
重要值=(相对密度+相对频度+相对显著度)/3
(1)
Margalef 丰富度:R=(S- 1) /lnN
(2)
Simpson 优势度:λ=∑[Ni(Ni-1)/N(N-1) ]
(3)
Shannon-Wiener 多样性:H=-∑PilnPi
(4)
Pielou 均匀度:Jsw=-∑(PilnPi)/lnS
(5)
式(2~5)中:S为样区中的物种总数;N为所有物种总个体数;Ni为第i种的个体数;Pi为第i种的个体比例。
Jaccard相似性指数:Cj=j/(a+b-j)
(6)
Sorenson相似性指数:Cs= 2j/(a+b)
(7)
Whittaker 相异性指数:βw=S/ma-1
(8)
Cody相异性指数:βc= [g(H) +I(H) ]/2
(9)
式(6~9)中:S为所研究系统中记录的物种总数;ma为各样方或样本的平均物种数;g(H)是沿生境梯度H增加的物种数目;I(H)是沿生境梯度H失去的物种数目;j为两个样地共有的物种数;a和b分别为样区A和样区B的物种数。
调查的数据均录入Excel中进行初步处理,并采用SPSS 17.0统计分析软件对数据进行相关性分析。
3 结果与分析
3.1 群落结构及物种组成特征
如表1、图1所示,采伐迹地经30年天然更新的A区以中小径木为主,尚未有明显的优势树种,重要值依序为绒毛润楠(Machilusvelutina)、青冈、福建青冈,树种间差异较小。采伐后35年的B区,林分密度下降,蓄积量上升,优势种逐渐明显,重要值受大型乔木影响,依序为青冈、木荷、栲树。采伐后40年的C区呈现明显的复层林结构,冠层优势林木为青冈,树高可达20 m以上,优势木的重要值与其他树种的差异也达到最大,下层树种开始有二代的青冈出现。采伐后50年的D区,蓄积量达到最高峰,林分密度亦有所上升,冠层优势树种青冈、木荷与其他次优势树种的重要值逐渐降低。采伐后65、75年的E、F区,蓄积量有所下降,林分密度变化不大。

表 1 闽北地区青冈林不同演替阶段物种数量变化及主要树种相对重要值(IV)Table 1 Changes of species number and relative importance of main tree species in different succession stages of Cyclobalanopsis glauca forest in Northern Fujian (IV)
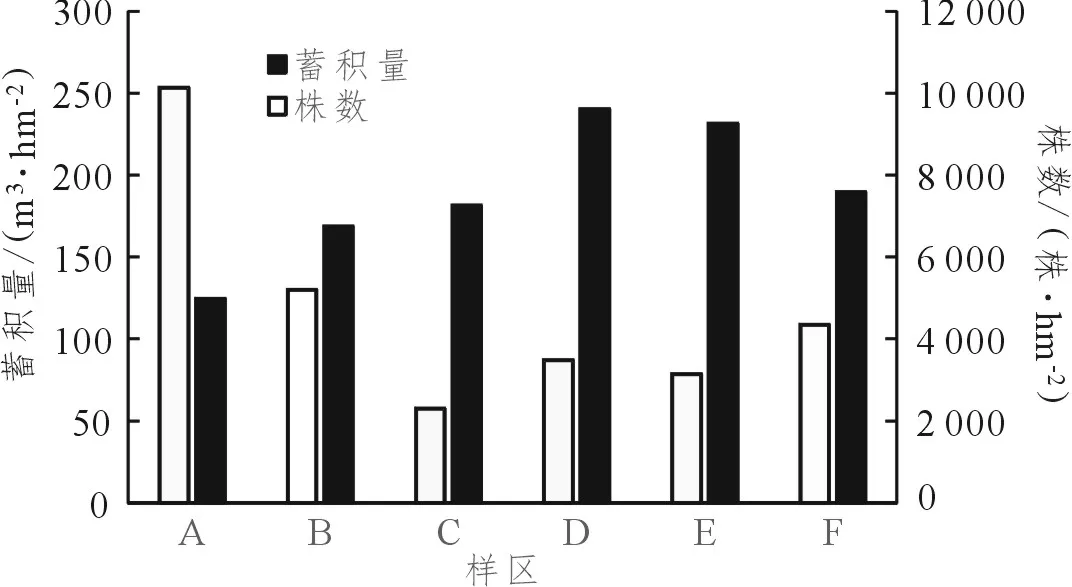
图 1 闽北地区青冈林不同演替阶段蓄积量与株数变化 Figure 1 Changes of volume and plant number of Cyclobalanopsis glauca forest in different succession stages in Northern Fujian
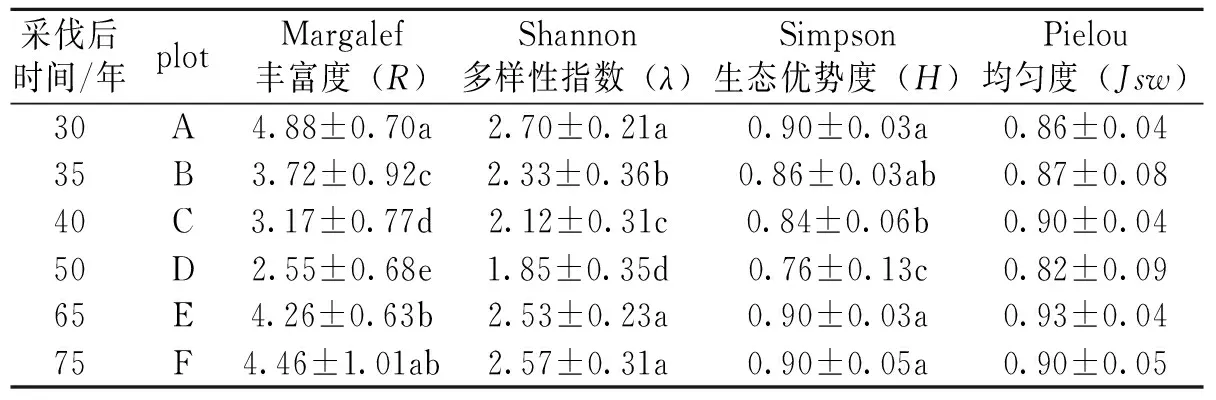
表 2 闽北地区青冈林不同演替阶段α多样性测度变化 Table 2 Change of α Diversity measure during different succession stages of Cyclobalanopsis glauca forest in Northern Fujian

表 3 闽北地区青冈林不同演替阶段β多样性测度变化 Table 3 Change of β diversity measure during different succession stages of Cyclobalanopsis glauca forest in Northern Fujian
3.2 α多样性特征
Margalef丰富度可用来表现物种个数与个体总数间的关系,Simpson优势度可反映物种集中性,Shannon多样性则可反映出物种的不定性,这3项指数虽然概念不同,但指数的高低主要受到物种数与个体总数的影响。调查结果显示这3项指数随着更新时间的增加,呈现先下降再上升的趋势,以A区最高, D区最低,到E、F区后3项指数又上升。Pielou均匀度指数则可以表现群落中不同物种多度分布的均匀程度。本研究的调查分析结果显示,均匀度指数在样区间并无显著的差异(表2)。
3.3 β多样性特征
β多样性为沿着环境梯度的变化物种替代的程度,与α多样性无直接的相对关系,主要受物种更替速率的影响,其中Jaccard相似性指数、Sorenson相似性指数可表现共同物种的相对比例,Whittaker 相异性指数及Cody相异性指数则可表现出新增/减少物种与物种总数量之间的关系。调查结果显示,相似性指数呈现先下降再上升的趋势,相异性指数则反之。演替阶段A-B林分密度下降最多,而物种增减变动量较小,相似性指数较高,演替阶段B-C开始有明显的冠层优势树种并逐渐郁闭,部分阳性树种被耐阴的树种所取代,到E-F阶段时相似性指数上升,林分的种类变化相对较低,有较高的相似性(表3)。
4 讨论与结论
4.1 青冈林采伐后初期的更新情形
调查显示,本试验地块状择伐天然更新后,林地优势树种为福建青冈、青冈、栲树、马尾松及木荷等。主要是周边及防火林带成熟林分中,具有果翅的种子如马尾松、木荷等,借助风力进入采伐林地,此类传播形态的树种通常为阳性树种,另一类为壳斗科为代表的坚果类种子,仰赖动物的传播移动或是二次传播,此类树种的发芽形态通常为子叶留土型(hypogealgermination),以耐阴树种为主[9]。采伐迹地在演替初期产生幼苗并发育为小型乔木,具有林分密度最高、蓄积量最低、物种丰富度、多样性指数及生态优势度最高的林分特征。
4.2 林木的竞争与替代
随着林木的发育,冠层逐渐郁闭,在B、C区出现大量中小径木死亡,但蓄积量维持稳定上升,林分逐渐呈现明显的复层林结构,上层以木荷、青冈为主要冠层优势树种,与少部分的马尾松、栲树、石栎等混生,其中马尾松在林缘生长较好,形成冠层的优势林木,林内则因强烈的竞争而生长不良或干枯死亡,这主要是由于群落内光照减弱及相应的马尾松物质代谢能力的变化[10]。下层的阳性树种如野漆树(Toxicodendronsuccedaneum)、赤杨叶(Alniphyllumfortunei)等,逐渐被耐阴的茶科(Theaceae)、山矾科(Symplocaceae)等树种取代,物种替代速率较高,这与陈世品研究结果[11]一致。林下有较多的青冈、福建青冈萌发种子及少量实生苗,这可能主要是由于林分密度大,种子成苗过程中获取光照等营养空间受阻所致,这与黄雍容等学者研究结果[12]一致。
4.3 老熟林木死亡与幼苗填补
调查显示,从D区到F区株数呈现小幅度的波动,蓄积量下降,主要是由于天然更新发育约65年后,冠层部分优势林木老化或受雷击、风害等外力破坏后,发生真菌、虫蛀等感染导致林木生长衰退死亡,产生的林窗增加林内光照时间及光量[13],林下以壳斗科为主的幼苗获得足够的光照而加速生长,同时也有少量其他物种种子传播至林窗内更新生长,提升了林分密度及生物多样性指数,此外,福建青冈的老熟林木在生长衰退后易发生根萌,这与前人的结果[11-12]相符,形成丛生的中小径木,青冈则根萌情况并不明显。
4.4 青冈次生林培育方式探讨
壳斗科植物为福建省的天然林、次生林的主要优势树种,因其适应性良好而被广泛运用在用材林、薪炭林或作为经济果树[14]。由于省内各地的立地条件不一,适宜的树种也会产生差异[15],而在退化生态系统恢复过程中,物种多样性变化剧烈,天然更新过程漫长[3,11]。本试验地以青冈、福建青冈为主要优势林木择伐后,探讨如何尽快达到稳定群落的近自然经营方式。首先,在林木更新初期,萌生与实生呈镶嵌状并存,密度较低或过密,立木的年龄与径级分布不规则等[3],采取人工促进方法,对丛生的目标树种萌条,每丛保留2~3株主干通直、冠型圆满的,其余伐除,并在林缘和林窗进行播种或植苗作业,补充目标乡土树种[16-17]。其次,在天然更新约30年后,林内发生竞争,根据天然次生林的特点与经营目标,因地制宜地构建林分间伐指标体系[18],围绕目标树伐除干扰树,释放空间促进目标树种生长。最后,在天然更新约50年后,林分生长缓慢、蓄积量最大和生态稳定性较高,可逐步择伐利用达到目标胸径的目标树,在森林出现衰退前获取其自然会损失的一部分而维持森林生物的总量[19],并保留部分生长良好的母树,保障丰富种子来源[16,20],作为珍稀乡土树种的培育方式。
志谢:
本次调查得到福建林业职业技术学院巫智斌、黄华明、俞群老师的帮助,谨此致谢!
