舟山长白海域春秋季鱼类群落结构特征研究
2021-10-27求锦津王咏雪胡成业聂振林水柏年
求锦津 ,王咏雪,胡成业,孙 鹏,聂振林,田 阔,水柏年
(1.浙江海洋大学水产学院,浙江舟山 316022;2.台州市海洋环境检测中心,浙江台州 318000;3.中国海洋大学水产学院,山东青岛 266100)
舟山长白海域位于舟山本岛西北部的近岸,既毗邻杭州湾口又位于岱衢洋海域。该海域是著名的岱衢族大黄鱼Larimichthys crocea,小黄鱼L.polyactis、鮸Collichthys lucidus 等重要经济鱼类产卵、索饵、育肥的重要场所,因此舟山长白海域具有重要的生态价值。因其位于钱塘江口,且受长江冲淡水系、台湾暖流、中国沿岸流等复杂的流系影响,该海域为重要的生态敏感区。近年来,随着对海洋生物多样性保护及可持续利用的日益重视,多样性以及群落结构特征等领域成了研究热点。国外一些学者开展了生物群落组成、时空变化、结构特征及其与水深、底质等非生物环境因子相关关系等方面的研究[1-3]。国内一些学者对近岸海洋鱼类的研究,也大多涉及群落结构特征、变化及影响因素等,如刘勇等[4],王理想[5]均从鱼类群落的种类、数量、优势种、物种多样性等方面入手,结合聚类分析等方法探究了鱼类群落结构及时空变化;王小林等[6],董静瑞等[7]运用典范对应分析(CCA)等方法分析了鱼类群落结构与非生物环境因子(水深、水温、盐度、pH 等)的相关性。杭州湾口的金塘岛、岱衢洋等长白邻近海域的鱼类群落研究仅涉及鱼类资源密度及物种多样性等方面[8-9],未见有关长白海域鱼类群落结构及其与环境因子相关关系方面的研究及报道。因此,本文试图通过春、秋2 季长白海域调查数据,研究该海域鱼类的种类组成、优势种、生态位、群落分布与环境因子的相关关系等,以期为长白海域鱼类资源的保护、管理及可持续利用的应用研究提供科学理论参考,同时也为鱼类生物多样性的研究积累资料。
1 材料与方法
1.1 研究区域
2016 年10 月(秋季)和2017 年4 月(春季)在舟山长白海域 (121°49′54.55″-122°18′33.98″ E,30°01′44.46″-30°18′58.27″ N)开展了渔业资源调查以及同步调查了6 个主要环境因子(水温、盐度、悬浮物、溶解氧、化学需氧量及pH),2 季调查分别设置12 个调查站位(图1)。渔业资源调查底拖网规格、拖速以及生物学测定等均按《海洋调查规范》[10]要求执行,实际拖网时间为0.5~1 h,在数据分析前进行拖网时间(1 h)的标准化处理。鱼类种类鉴定参照相关文献[11-14]。
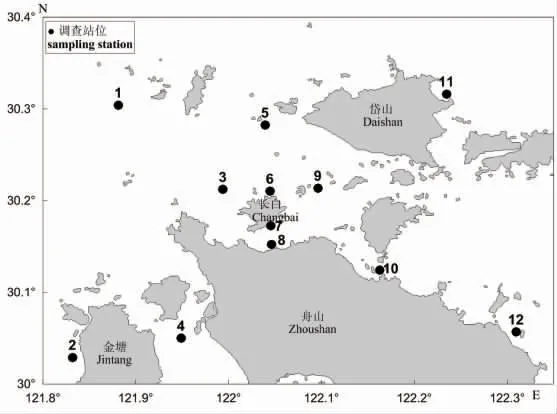
图1 舟山长白海域渔业资源调查站位Fig.1 Survey stations of fisheries resource in Changbai
1.2 数据处理
1.2.1 优势种
优势种判断采用PINKAS,et al[15]的相对重要性指数IRI,计算公式如下:

式(1)中,IRI 为相对重要性指数,N 为某鱼类尾数占总尾数的百分比,W 为该物种重量占总重量的百分比,F 为出现频率。优势种的判定采用王雪辉等[16]的划分标准,将IRI>1 000的种类定为优势种,1 000~100 为重要种,100~10 为常见种,10~1 为一般种,<1 为少有种。
1.2.2 生态位宽度
生态位宽度采用Shannon 指数[17],计算公式如下:

式(2)中,Pij为种i 在j 个站位中的个体数占站位总个体数的比例;R 为总站位数;Bi为生态位宽度,取值范围为[0,R],Bi值越大,生态位宽度就越大。
1.2.3 长度谱
借鉴有关学者的研究方法[18-19],根据各站位不同种类体长(肛长或叉长)(L,cm),设置长度谱变化组距为5 cm,然后将相应种类的单位网次单位时间渔获尾数ind·h-1分配到不同长度组中,以各长度组渔获总尾数的自然对数(lnN)为纵坐标,长度组中值(L)为横坐标作两者的关系图,并对两者进行线性回归,得到长度谱斜率和截距。斜率与鱼类群落的营养传递效率有关,直线越陡,营养循环效率越低;截距与生产力水平有关,截距越大,生产力水平越高。
1.2.4 冗余分析
运用Canoco 4.5 软件对鱼类与环境因子之间的关系进行分析及排序图绘制。先对单位时间渔获质量(g·h-1)数据进行除趋势对应分析(DCA),根据每个轴的梯度长度(LGA)选择适宜的排序方法。当LGA<3 时,选择冗余分析(RDA);当LGA>4 时,则可运用典范对应分析(CCA);当3<LGA<4 时,2 种分析均可[20]。DCA分析结果显示,LGA 均<3,故本文采用线性模型的冗余分析。
2 结果与分析
2.1 种类及优势种组成
春、秋2 季共鉴定出鱼类24 种,其中秋季17 种,隶属于7 目10 科14 属,春季18 种,隶属于6 目10科16 属。2 个季节共同种为11 种,以不作长距离洄游的种类为主,如龙头鱼Harpodon nehereus、棘头梅童鱼Collichthys lucidus、虾虎鱼类Gobiidae sp.和舌鳎类Cynoglossus sp.等。其余种类以长距离洄游性鱼类为主,如海鳗Muraenesox cinereus、黄姑鱼Nibea albiflora、灰鲳Panpus cinereus、蓝点马鲛Scomberomorus niphonius、四指马鲅Eleutheronema tetradactylum 等。
如表1 所示,秋季,优势种为龙头鱼和棘头梅童鱼,重要种为鮸和孔虾虎鱼Trypauchen vagina,龙头鱼的重量和尾数百分比均有绝对优势,为绝对优势种。春季,优势种有孔虾虎鱼和刀鲚Coilia nasus,尾数分别占春季鱼类总尾数的46.02%和12.80%,重要种有龙头鱼、凤鲚Coilia mystus、棘头梅童鱼、鮸、蓝点马鲛、褐斑三线舌鳎Cynoglossus trigrammus 等6 种。2 个季节无共同优势种,仅鮸为共同重要种。
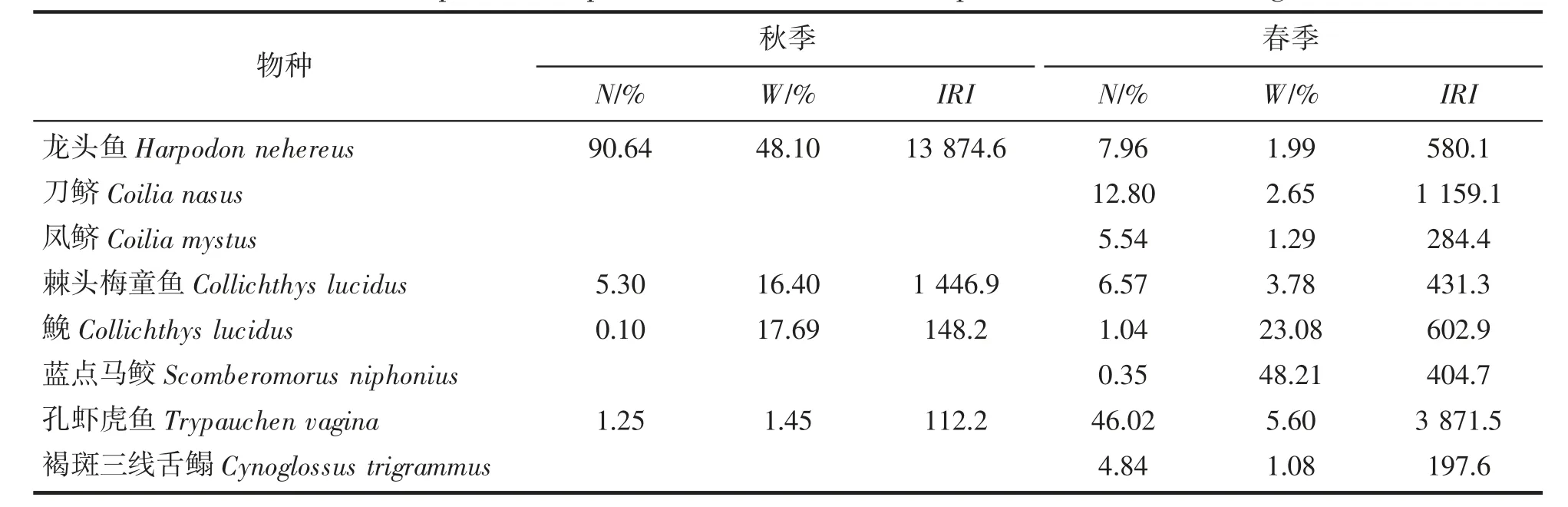
表1 长白海域鱼类优势种和重要种相对重要性指数值(IRI)Tab.1 Fish species composition and the relative importance index in Changbai
2.2 生态位宽度
春、秋2 季鱼类生态位宽度值变化大,且总体来说,春季生态位宽度值高于秋季(表2)。秋季,生态位宽度值变化范围为0.03~1.52,其中生态位宽度最大的是棘头梅童鱼(1.52),四指马鲅的生态位宽度最小(0.03);春季,鱼类生态位宽度值变化范围为0.09~3.09,孔虾虎鱼的生态位宽度值最大(3.09),灰鲳和矛尾虾虎鱼的生态位宽度较窄,仅为0.09。比较春、秋2 季共同种生态位宽度值可得,同一物种的生态位宽度值,春季均高于秋季(图2),不同种的生态位宽度总体上也是春季高于秋季,均值分别为0.38(0.09~1.05)和0.08(0.03~0.23)。
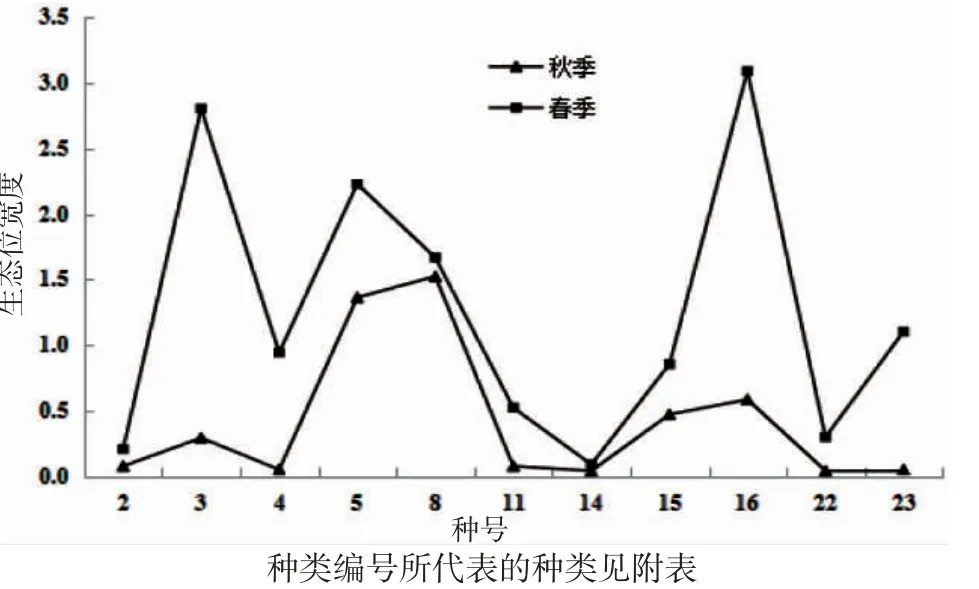
图2 春、秋2 季共同种生态位宽度比较Fig.2 Comparison of niche breadth of common species in the spring and autumn
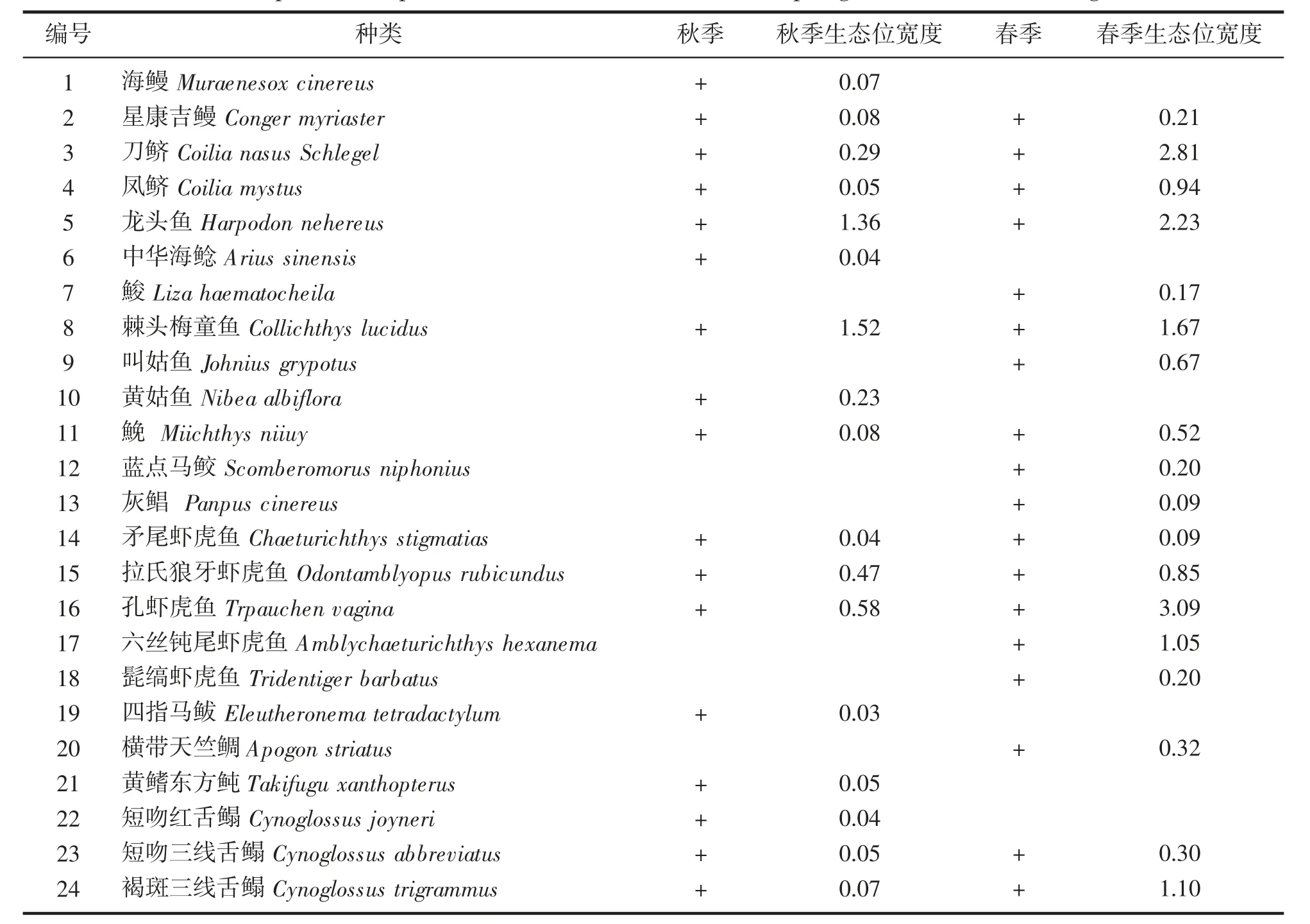
表2 长白海域春、秋2 季物种组成及生态位宽度Tab.2 Species composition and niche breadth in the spring and autumn of Changbai
2.3 春、秋季长度谱比较
由春、秋季鱼类长度谱分析可知,春、秋季鱼类群落均以小型个体为主,鱼类尾数的自然对数(lnN)和体长呈负相关,大个体鱼类尾数比例远小于小个体尾数比例(图3)。秋季,鱼类个体体长主要集中在5~10 cm,鱼类尾数占总尾数的65.59%。春季,鱼类个体体长主要集中在5~10 cm 和10~15 cm,其尾数分别占总尾数的35.29%和43.25%。体长20 cm 以上的个体均很少,即秋季和春季鱼类尾数分别仅占当季鱼类总尾数的0.83%和2.77%。线性回归趋势线还得出,秋季鱼类群落长度谱直线斜率的绝对值和截距均大于春季。
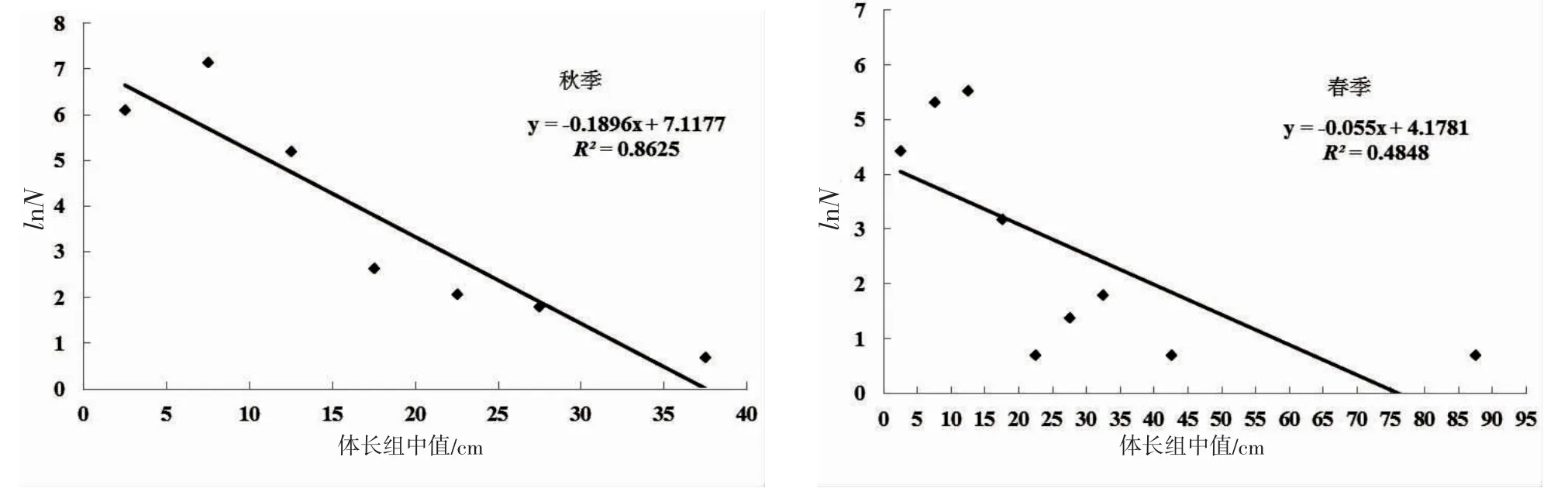
图3 长白海域鱼类春、秋季长度谱Fig.3 The size spectrum of fish in the spring and autumn of Changbai
2.4 鱼类群落与环境因子的关系
如图4 所示,RDA 分析得出,秋季,第一轴特征值为0.208,第二轴为0.129,2 个轴共解释了物种与环境因子累计方差的66.2%,第一轴和第二轴物种与环境因子的相关系数分别达0.855 和0.876,表明RDA排序能较好地反应鱼类群落与环境因子的关系。第一排序轴与pH 呈最强正相关(0.473),与温度呈最大负相关(-0.565),从左到右pH 逐渐升高,温度则逐渐下降;第二排序轴与溶解氧和悬浮物呈较强正相关(分别为0.578 和0.436),与化学需氧量呈最大负相关(-0.583),从上至下溶解氧和悬浮物逐渐降低,化学需氧量则逐渐升高。排序图显示,龙头鱼、拉氏狼牙虾虎鱼Odontamblyopus rubicundus、四指马鲅等种类分布均与pH 呈负相关性,而刀鲚等与pH 则呈正相关性;孔虾虎鱼、髭缟虾虎鱼Tridentiger barbatus 则主要分布在盐度较高的区域。
春季,第一轴特征值为0.221,第二轴为0.159,2 个轴共解释了物种与环境因子累计方差的59.1%,第一轴和第二轴物种与环境因子的相关系数分别达0.973 和0.896,表明RDA 排序也能较好地反应春季鱼类群落与环境因子的关系。第一排序轴与pH 呈最强正相关(0.261),与化学需氧量呈最强负相关(-0.475),从左到右pH 逐渐升高,化学需氧量则逐渐下降。第二排序轴与盐度、化学需氧量呈较强正相关(分别为0.701 和0.647),与溶解氧呈最强负相关(-0.641),从上至下盐度、化学需氧量逐渐降低,溶解氧则逐渐升高。排序图显示,龙头鱼、拉氏狼牙虾虎鱼、叫姑鱼Johnius grypotus 等分布在化学需氧量较高的区域,而与pH 呈负相关性;六丝钝尾虾虎鱼Amblychaeturichthys hexanema、褐斑三线舌鳎与溶解氧呈正相关,主要分布在溶解氧较高的区域;而孔虾虎鱼、鮻Liza haematocheila、棘头梅童鱼等物种与盐度、悬浮物呈负相关(图4)。
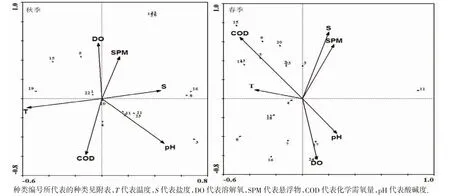
图4 长白海域鱼类群落与环境因子的RDA 分析Fig.4 RDA analysis of fish community and environmental factors in Changbai
3 讨论
3.1 鱼类群落组成特征
种类组成是决定群落性质最重要的因素,也是鉴别不同群落类型的基本特征[21]。长白海域春、秋2 季鱼类种类较少,分别为18 种和17 种,这可能与近些年渔业资源捕捞过度、附近临港工业造成污染等给鱼类栖息环境带来负面影响有关。长白海域春、秋2 季鱼类种类数相近,春、秋季定居性或短距离洄游种类均为11 种,其中8 种为2 季的共同种,因此,总体而言该海域定居性或短距离洄游种类比例较高,这表明该海域鱼类群落具有显著的定居性或短距离洄游特征。这可能与该海域所处的地理位置、气候特征、复杂的流系配置等特征有关。共同的重要经济种类龙头鱼、棘头梅童鱼在秋季均为优势种,而在春季却仅为重要种,这可能是春季一些长距离洄游的大个体种类随台湾暖流向北向岸延伸而分布在长白海域,占据了一部分的资源和空间,秋季随着台湾退缩南下离岸而去,导致了群落组成的变化。另外,春、秋2 季长距离洄游性种类则分别为7 种和6 种,且均为重要经济种类。2 季共同的长距离洄游种类有3 种,依次为刀鲚、凤鲚和鮸,其中刀鲚和凤鲚分别为春季的优势种和重要种。这符合庄平等[14]的研究,即刀鲚和凤鲚等成熟大个体在春季繁殖季节经过杭州湾及毗邻海域进行产卵或生殖洄游,秋季则顺流进入海水里育肥、越冬。鮸为春、秋2 季共同的重要种,这可能与鮸分布于包括长白海域在内的岱衢洋,为其传统上天然的重要产卵、索饵场有关。综上所述,春、秋2 季龙头鱼、鮸、蓝点马鲛和棘头梅童鱼等优势种或重要种以重要经济种类为主,且多为营养级较高的肉食性种类,这表明该海域群落一定程度上仍然体现群落组成种类经济价值较高,渔场价值较高的特征。
3.2 生态位
生态位宽度可用于反映物种的分布范围、分布格局以及对环境的适应能力或对资源的利用程度[22-23]。春、秋2 季鱼类生态位宽度值呈现显著的季节变化,即同一物种的生态位宽度值,春季均高于秋季,不同种类仅秋季黄姑鱼的生态位宽度值(0.23)较大,其余物种也均小于春季其他物种的生态位宽度值。究其原因,可能是春季台湾暖流进入带来的高温高盐,使得定居性或短距离洄游种类群体由深水区向浅水区扩散,站位出现频率提高,分布范围较秋季扩大;长距离洄游性种类群体则随台湾暖流进退进行洄游,4 月随台湾暖流进入调查海域,鱼类群落种数显著增多,各站位出现频率高,分布广泛。因此,长距离洄游性和定居性种类春季生态位总体较秋季宽,这也说明了秋季鱼类分布较为集中,可能是鱼类集群为越冬洄游作准备的状态。
求锦津等[24]在研究舟山长白海域主要游泳动物生态位宽度和相对重要性指数之间相关性时得出,在物种均衡分布的情况下,IRI 值越大,Bi 值也越大,本文研究结果也类似。秋季第一优势种孔虾虎鱼与第二优势种刀鲚的IRI 值大,Bi 值也大;秋季鮸的IRI 值(602.9)远大于凤鲚(284.4)、棘头梅童鱼(431.3)等种类,但其生态位宽度值很小(0.52)。这可能是鮸的尾数占鱼类总尾数的百分比较小(1.04%),而重量百分比较大(23.08%),导致了IRI 指数值较大,而生态位宽度却包含了物种尾数、出现频率和分布均匀性等信息,使得生态宽度仍然较小。因此,IRI 值与Bi 值之间无绝对的正向相关关系。
3.3 鱼类长度谱
长度谱分析能够直接反应物种的长度组成及分布,能够在一定程度上反映生物群落的能量流动和群落结构组成,衡量群落结构的变化[25]。春、秋2 季鱼类体长主要集中在5~15 cm的范围,表明2 个季节渔获物小个体种类数比例较高,总体呈现小型化与低龄化。秋季鱼类长度谱斜率的绝对值和截距均大于春季,即秋季,营养循环效率低,生产效率较高,该季鱼类群落中的大型个体少,以小型个体为主导;而春季的营养循环效率较高,生产效率低,春季成熟大个体鱼类到沿岸产卵或经过沿岸进行产卵洄游,大型个体增加。这与春、秋季底拖网所获鱼类重量、尾数大小趋势相一致,即秋季共捕获鱼类1 922 尾,共计重量8 526 g,重量均值为(11.85±52.14)g;春季捕获鱼类尾数578 尾,总重却高达20 160 g,重量均值可达(39.56±320.40)g。这与王晶等[19]研究得出的调查海域生产力水平在夏、秋季较高,冬、春季较低的结论几乎一致。
3.4 鱼类分布与环境因子的关系
RDA 排序图的第一排序轴显示,春、秋2 季鱼类群落分布均受pH 影响,且龙头鱼、拉氏狼牙虾虎鱼等多数物种与pH 呈负相关。pH 是水域中一些化学性的总和,在水体中相对稳定,与生物的生长、新陈代谢以及生殖发育均有密切联系[26],因此2 季的物种分布受pH 影响具有相似性。而刀鲚、凤鲚、星康吉鳗等种类受pH的影响因季节不同而不同,即秋季,3 个物种主要在pH 较高的区域分布,春季则相反。这可能是物种本身生理、生态行为的季节性变化引起的,如刀鲚、凤鲚为溯河性产卵鱼类,于春季在江河口进行产卵或产卵洄游,远离高pH 区域。另外,其他环境因子的季节性变动也会对pH的作用力产生影响。RDA 排序图的第二排序轴显示,溶解氧、化学需氧量等因子也是影响春、秋2 季鱼类群落分布的重要因子。MAES,et al[27]研究认为,鱼类更适于溶解氧含量高的海域生活。张迎秋等[28]认为,溶解氧含量可以影响鱼类群落的分布及结构。调查海域2 季溶解氧含量均较高但变化小(秋季为(8.31±0.09)mg·L-1,春季为(9.67±0.05)mg·L-1),因此溶解氧虽为影响2 季物种分布的因子,但多数物种分布未表现出明显的高溶解氧分布趋势。舟山长白及附近海岛船舶修造业、化工业等发达,带来有机污染导致了化学需氧量较高,从而影响物种的分布。众多研究[6-7,29]还表明,水温是影响海洋生物群落分布、群落结构的最重要因素。4 月的春季调查,台湾暖流尚未完全到达调查海域,水温(15.52±0.56) ℃较低,而10 月的秋季调查台湾暖流尚未完全撤退,水温较高(23.52±0.38) ℃。RDA 分析显示,温度是影响秋季物种分布的重要因子,如孔虾虎鱼与温度呈负相关,因此该物种在低温的春季分布数量明显多于秋季。
4 结论
舟山长白海域毗邻杭州湾口,又位于岱衢洋,为著名的大黄鱼、鮸等石首鱼科鱼类天然产卵场。调查研究得出,鮸、凤鲚、刀鲚和棘头梅童鱼等为春、秋2 季共同出现的优势种或重要种,也是典型特色种类,更是重要的经济种类,显示出长白海域鱼类仍然以重要经济种类为主。生态位分析得出,秋季鱼类分布较为集中,伏季休渔刚结束,鱼类群体即承受着高强度、无序的捕捞,导致其承受强大的捕捞压力,必然面临严重的资源衰退风险,这也可能是长期伏季休渔未能有效扭转重要渔业资源衰退的重要原因。此外,渔获物中鱼类群落的优势种相对较少,且渔获物小个体种类数比例较高,呈现小型化、低龄化。冗余分析可知,春、秋2 季鱼类群落除了受温度、盐度、pH 等环境因素影响外,还受到化学需氧量的重要影响,其对鱼类的生长、发育及繁衍均存在较大的负面影响。因此,鉴于海洋开发的强大压力与海洋生态面临被破坏的严峻态势,加强该海域鱼类资源的监测、保护与管理,为棘头梅童鱼、鮸鱼等重要经济鱼类提供良好的产卵场、索饵场及洄游通道是当务之急,也可为今后长白海域鱼类资源修复、管理及可持续利用提供良好的基础资料。
