舟山渔场大小鱼山附近海域鱼类群落结构及生物多样性
2021-10-27唐嘉威于南京许永久俞存根
唐嘉威,于南京,郑 基,许永久,俞存根
(浙江海洋大学水产学院,浙江舟山 316022)
舟山渔场是我国著名的四大渔场之一,它是多种渔业生物索饵、繁殖、洄游、生长的重要场所,是我国优质海水鱼虾蟹类的高产区。大小鱼山位于舟山渔场灰鳖洋海域,受台湾暖流以及长江、钱塘江径流的影响,大小鱼山附近海域营养盐丰富,渔业资源种类繁多。过去,曾有许多学者对舟山渔场的渔业资源做过研究,如俞存根等[1]报道了舟山渔场及邻近海域鱼类种类组成和数量分布情况;邓小艳等[2]报道了舟山近岸海域春秋季鱼类种类组成及其数量时空分布;陈小庆等[3-4]报道了舟山渔场及邻近海域浮游动物数量分布特征;而对舟山渔场范围内某些局部海域的鱼类资源研究报道也有不少[5-8]。但是,针对大小鱼山附近海域的鱼类群落结构研究尚未见报道。本文根据2019 年4 月(春季)、10 月(秋季)在舟山渔场大小鱼山附近海域开展渔业资源底拖网定点调查所获得的鱼类资料,对该海域的鱼类种类组成、优势种、数量时间分布、生物多样性、ABC 曲线等群落结构特征进行分析,以期进一步提高对舟山渔场鱼类群落生态学及渔业资源状况的认知能力,并为该海域鱼类资源动态变化及合理利用和保护提供科学依据。
1 材料与方法
1.1 数据来源
本文所用数据为2019 年4 月(春季)和10 月(秋季)在舟山渔场大小鱼山附近海域(30.00°-30.59°N,121.67°-122.18°E)开展渔业资源底拖网定点调查获得的鱼类资料,调查共设置8 个站位,具体站位设置如图1 所示。调查船租用群众捕捞生产单拖网渔船,船号为浙普渔32207 号,主机功率为257 kW,网具规格为860 目×80 mm。调查时每站拖曳约1 h,各站位的平均拖速为3.5 kn。调查采集方法按照《海洋渔业资源调查规范》(SC/9403-2012)执行。对拖曳所获得的渔获物采用全部取样,即将每站拖网所获得的渔获物全部装入样品袋,并进行编号、记录后,冰鲜保存,带回实验室进行分析、鉴定,并对主要渔获种类进行生物学测定。种类名录以刘瑞玉(2008 版本)的《中国海洋生物名录》[9]为主要参考依据。
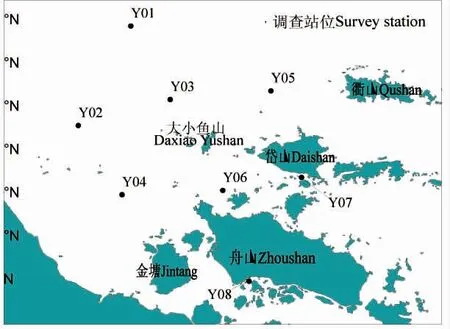
图1 渔业资源调查站位图Fig.1 The surwey stations of fishery resources
1.2 数据处理及分析方法
1.2.1 鱼类资源密度
鱼类的资源密度估算采用扫海面积法,计算公式如下:

式中:ρi为第i 站的鱼类资源密度(重量:kg·km-2;尾数:ind·km-2);Ci为第i 站的每小时拖网渔获物中鱼类数量(重量:kg·h-1;尾数:ind·h-1);ai为第i 站的拖网每小时扫海面积(km2·h-1),即网口水平扩张宽度(km)×每小时拖曳距离(km·h-1);q 为网具捕获率(本文q 取0.5)。
1.2.2 优势种计算公式
优势种的计算采用相对重要性指数,计算公式如下:

式中:N 是该站位占总位数的百分比(%),W 是该种占总质量的百分比(%),F 为该生物在调查站位中出现的频率,即出现站次数与总调查站次数的百分比。
1.2.3 生物多样性指数计算公式
生物多样性采用Shannon-Weaver 指数、均匀度指数、丰富度指数3 个公式计算,计算公式如下:

式中:H′为生物多样性指数;Pi是第种鱼类生物量占总生物量的比例;S 为鱼类总种数。

式中:J′为均匀度指数;H′为生物多样性指数;S 为鱼类总种数。
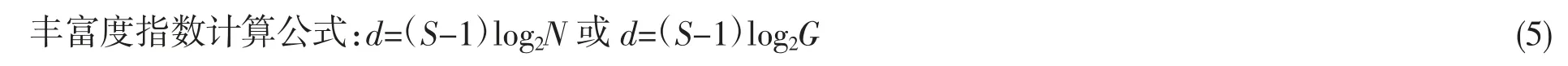
式中:d 为丰富度指数;N 为鱼类总尾数;G 为鱼类总生物量;S 为鱼类总种数。
1.2.4 群落结构和相似性分析
利用聚类分析法(CLUSTER)分析该海域2 个季节群落结构的相似度。利用非参数多维标序技术(nonmetric multidimensional scaling,NMDS)经过四次方转换,以使优势种在群落中的作用平稳,平衡稀有物种的效度,分析鱼类群落结构的变化。用胁迫系数(Stress)评价NMDS 二维点阵分布图效果,一般认为Stress<0.05 时,结果具有很好的代表性;0.05≤Stress<0.1 时,分析结果基本可信;0.1≤Stress<0.2 时,结果具有一定解释意义[10]。
利用SIMPER 分析方法研究群落贡献率,通过对季节间物种相异性和特征种以及分歧种的贡献率进行研究得出结论。利用单因子相似性分析(ANOSIM)检验不同季节鱼类群落结构的差异性。
1.2.5 ABC 曲线(丰度/生物量曲线)
丰度生物量比较曲线是在同一坐标系中比较生物量优势度曲线和丰度优势度曲线,通过2 条曲线的分布情况分析群落处于不同干扰状况下的特征[11],统计量用W 表示[12],计算公式如下:

式中:S 为出现的物种数,Bi、Ai分别为曲线中种类序号对应的生物量和丰度的累计百分比,群落处于未受到干扰的状态,W 值为正,其主要以生长慢、性成熟晚的个体较大的种类为主,生物量优势度曲线位于丰度优势度曲线之上;当处于中等干扰的状态时,2 条曲线处于相交状态;当群落处于严重干扰的状态,群落组成逐渐变为由生长快、个体较小的种类为主,此时生物量曲线位于丰度优势度曲线下方。
2 结果
2.1 种类组成
根据2019 年春秋两季在大小鱼山附近海域开展渔业资源底拖网调查获得的渔获物,经分析共鉴定出鱼类33 种,隶属于8 目、15 科、26 属,其中,以鲈形目的种类为最多,共有17 种,隶属于6 科、15 属;其次是鲱形目的种类,共有6 种,隶属于2 科、4 属;而仙女鱼目、刺鱼目、灯笼鱼目的鱼类种类均只有1 种。从站位出现率来看,春季,出现率最高的是凤鲚Coilia mystus,为75%,仅在1 个站位出现的鱼类数占42.10%;秋季,出现率最高的是龙头鱼Harpodon nehereus,为100%,仅在1 个站位出现的鱼类数占56.52%。具体鱼类种类组成及优势种如表1 所示。
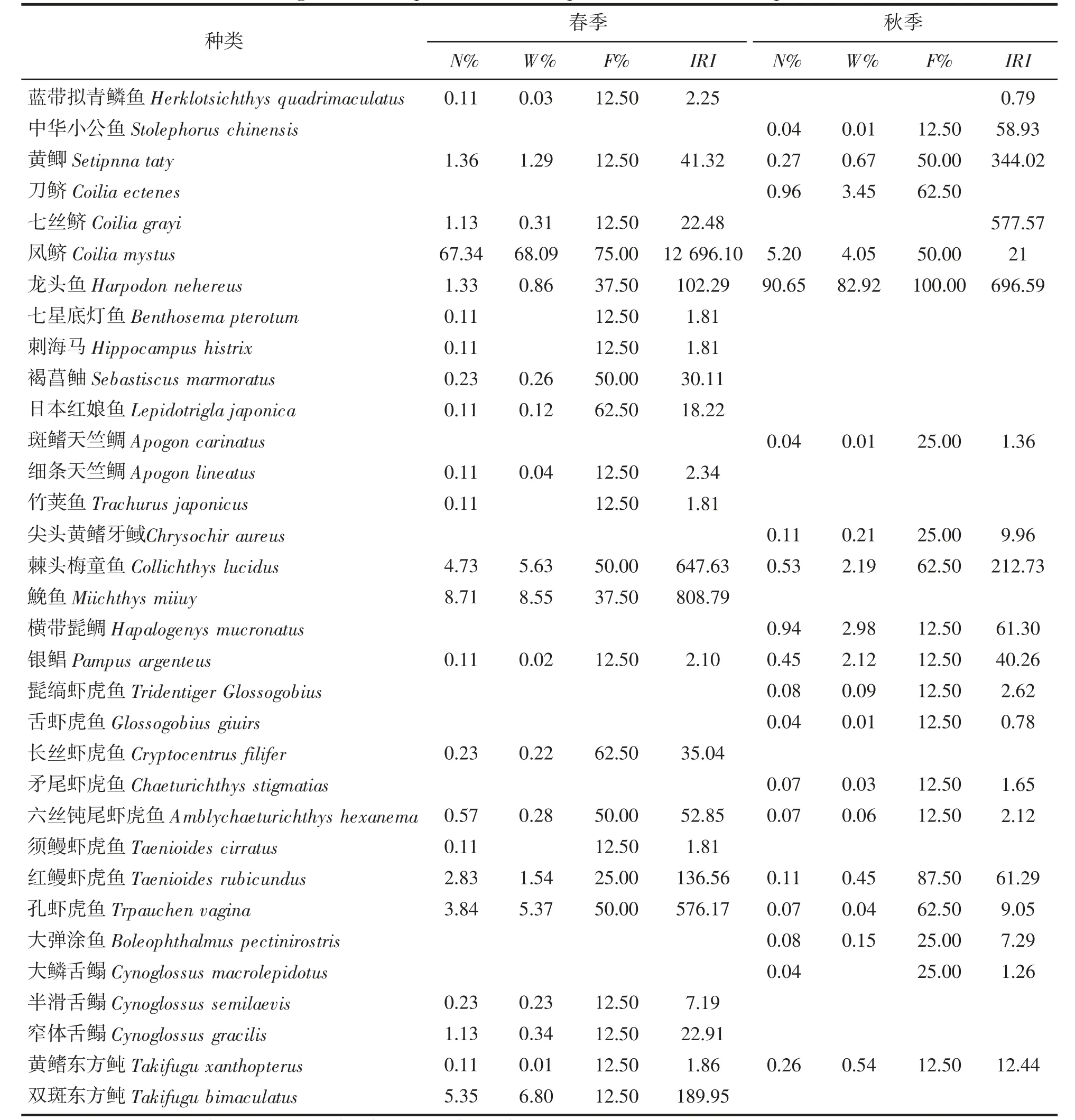
表1 调查海域鱼类种类组成及优势种Tab.1 Investigate the composition of fish species and dominant species in the sea
2.2 资源密度分布
图2、图3 分别为调查海域鱼类重量密度和鱼类尾数密度分布。从图中可以看出,在春季,大小鱼山附近海域鱼类重量平均密度为10.86 kg·km-2,其中,重量密度最高的是Y03 号站位,为52.10 kg·km-2,最低的是Y06 号站位,仅为0.05 kg·km-2;尾数平均密度为2.96×103ind·km-2,其中,尾数密度最高的是Y03 号站位,为8.19×103ind·km-2,最低的是Y06 号站位,为0.24×103ind·km-2,从空间分布来看,重量密度和尾数密度均以调查海域的西北部较高,南部海域较低。
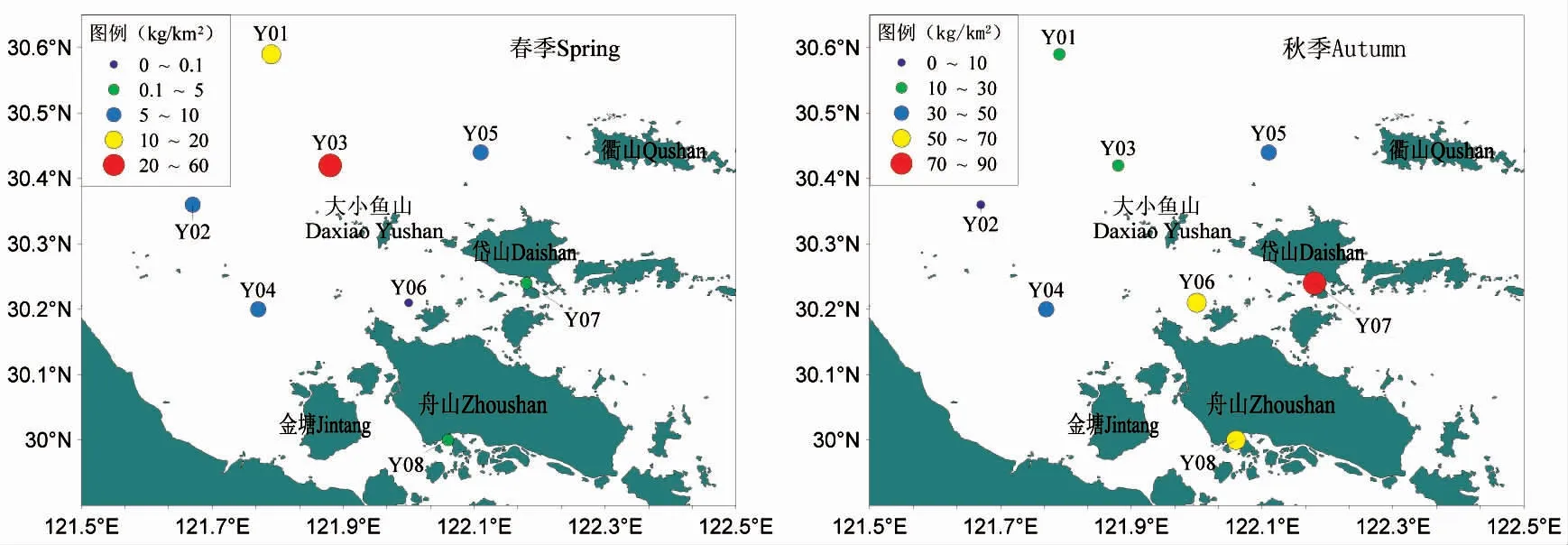
图2 调查海域鱼类重量密度分布Fig.2 Investigate the distribution of fish weight density in the sea
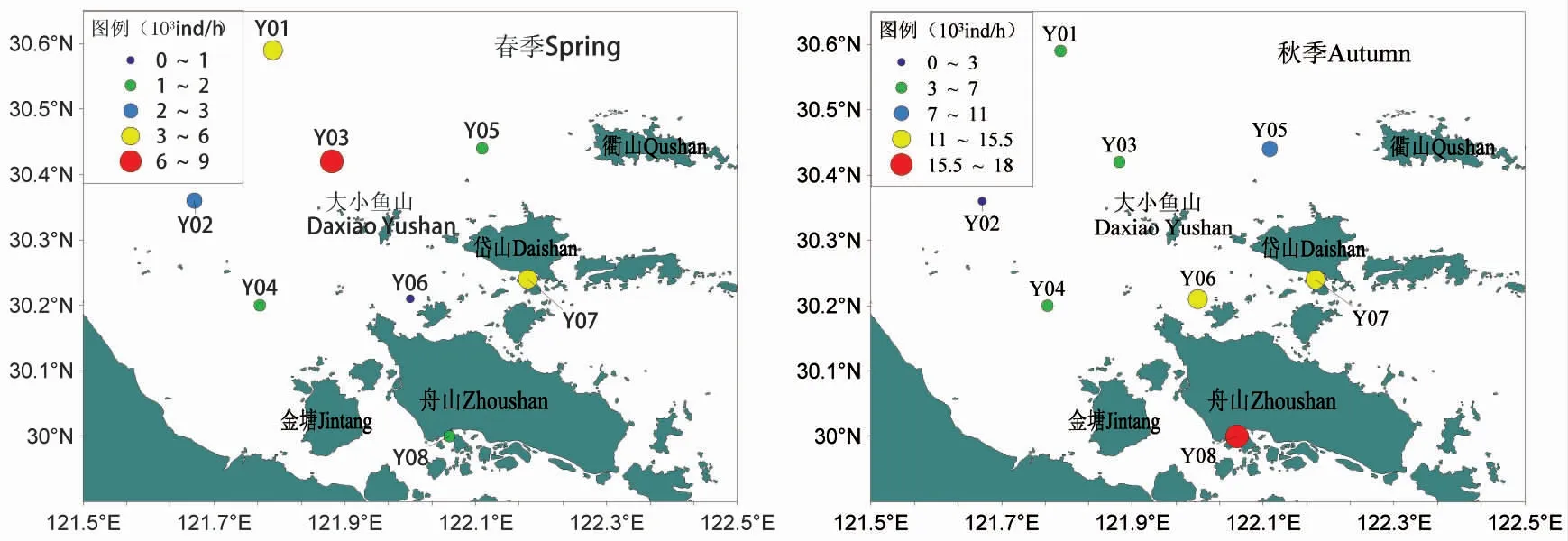
图3 调查海域鱼类尾数密度分布Fig.3 Investigation of fish tail density distribution in the sea area
秋季,鱼类重量平均密度为40.68 kg·km-2,重量密度最高的是Y07 号站位,为73.14 kg·km-2,最低的是Y02 号站位,为8.41 kg·km-2;尾数平均密度为9.18×103ind·km-2,尾数密度最高的是Y08 号站位,为15.56×103ind·km-2,最低的是Y02 号站位,为2.33×103ind·km-2,从空间分布来看,重量密度以调查海域东南部较高,西北部海域较低,尾数密度则以调查海域南部较高,西北部海域较低。
从季节分布来看,春季鱼类重量密度和尾数密度均低于秋季,并且数量密度较高和较低的站位各不同。
2.3 优势种和常见种
将相对重要性指数(IRI)大于1 000 者定为优势种,在100~1 000 之间者定为常见种[13],从表1 中看出,春季,调查海域的优势种仅有凤鲚1 种,常见种有红鳗虾虎鱼Taenioides rubicundus、棘头梅童鱼Collichthys lucidus、孔虾虎鱼Trpauchen vagina、龙头鱼、鮸鱼Miichthys miiuy、双斑东方鲀Takifugu bimaculatus 共6 种;从渔获量上来看,凤鲫占鱼类总重量的68.1%,其次是鮸鱼和双斑东方鲀,分别占7.0%、6.8%。秋季,优势种仅有龙头鱼1 种,常见种有刀鲚Coilia ectenes、凤鲚、棘头梅童鱼共3 种;龙头鱼占鱼类总重量的82.92%,其次是刀鲚和凤鲚,分别占4.05%、3.45%。
2.4 鱼类种类数的空间分布
表2 和图4 为调查海域不同水深区的鱼类种类数分布。春季,各站位鱼类种类数分布在3~14 种之间,秋季,各站位鱼类种类数分布在4~10 种之间。对比春秋季不同站位的鱼类种类数得知,春季以3 号和7 号站位鱼类种类数为较多,分别与12、11 种,以5、8 号站位种类数最少,均为3 种;秋季则以5 号和7 号站位鱼类种类数为较多,分别有10、9 种,以8 号站位鱼类种类数较少,仅有4 种。按水深将调查海域分为5~10 m、10~15 m、15~20 m 3 个水深带(表2)来观察鱼类种类数分布状况,春季,随着水深的增加鱼类种类数逐渐减少,即以5~10 m 水深区鱼类种类数最多,为16 种,以10~15 m 水深区鱼类种类数次之,为15种,以15~20 m 水深区鱼类种类数最少,仅有3 种;秋季,鱼类种类数随着水深的增加先增加后降低,以10~15 m 水深区鱼类种类数最多,为13 种,以5~10 m 水深区鱼类种类数次之,为11 种,以15~20 m 水深区鱼类种类数最少,为10 种,与春季相比,该水深区间鱼类种类数增加显著。总的来看,春、秋季鱼类种类数在10~15 m 水深区间最多,5~10 m 次之,15~20 m 水深区间最少。
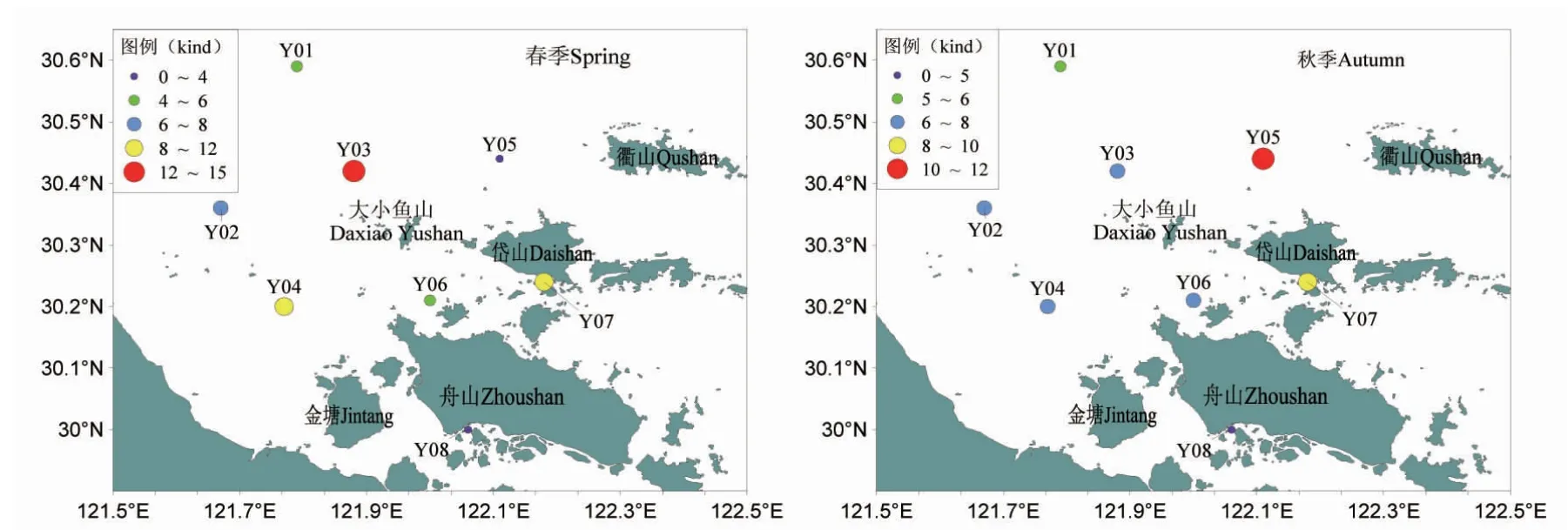
图4 调查海域鱼类种类数空间分布Fig.4 Survey the spatial distribution of fish species in the sea area
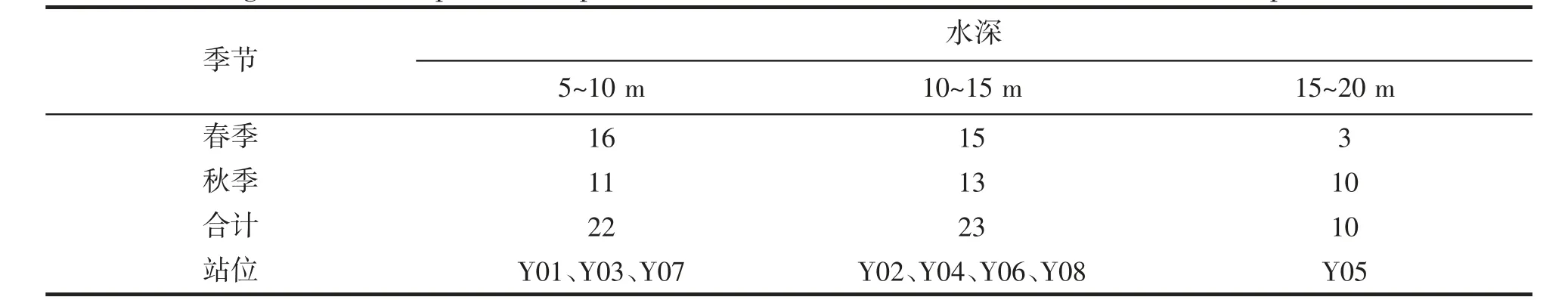
表2 调查海域不同水深鱼类种类组成和站点分布Tab.2 Investigation of fish species composition and station distribution in different water depths in the sea area
2.5 生物多样性
调查海域各站位春秋季鱼类生物多样性指数如表3 所示。

表3 调查海域各站位鱼类多样性指数Tab.3 The fish diversity index of each station in the surveyed sea area
从表3 可看出,春季,生物多样性指数(H')分布在0.08~2.89 之间,平均值为1.45,最高值位于7 号站,最低值位于5 号站位;均匀性指数(J')分布在0.05~4.33 之间,平均值为1.3,丰富度指数(d)分布在0.38~3.08 之间,平均值为1.64,二者最高值都位于6 号站位,最低值都位于5 号站位。秋季鱼类生物多样性指数(H')分布在0.09~1.51 之间,平均值为0.91;均匀性指数(J')分布在0.04~0.66 之间,平均为0.34;丰富度指数值(d)分布在0.40~1.21 之间,平均为0.81,生物多样性指数(H')和均匀度指数(J')最高值均位于1 号站位,丰富度指数(d)最高值位于2 号站位,3 个指数最低值均出现在8 号站位;总的来看,春、秋季大小鱼山附近海域不同站位鱼类多样性差异较大,且两个季节的多样性最高值和最低值分别出现在不同站位。
2.6 群落聚类分析
根据图5 鱼类群落CLUSTER 聚类分析和鱼类群落NMDS 二维点图得知,聚类分析和NMDS 分析的结果一致。且从春季鱼类群落CLUSTER 聚类分析可看出,在35%相似性水平上,调查站位可以被分为3个群组,第一群组有Y01、Y02、Y03、Y04、Y05 共5 个站位,代表性鱼类是凤鲚,第二群组只有Y06 一个站位,代表性鱼类是细条天竺鲷Apogon lineatus,第三群组有Y07、Y08 共2 个站位,代表性鱼类是凤鲚和六丝钝尾虾虎鱼Amblychaeturichthys hexanema。秋季,在55%相似性水平上,调查站位可以被分为4 个群组,第一群组有Y01、Y03、Y05 共3 个站位,代表性鱼类是龙头鱼和凤鲚,第二群组有Y07、Y08 共2 个站位,代表性鱼类是龙头鱼,第三群组仅有Y02 一个站位,代表性鱼类是龙头鱼和刀鲚,第四群组有Y04、Y06 共2 个站位组成,代表性鱼类有龙头鱼和棘头梅童鱼。春季鱼类群落相似性低于秋季,且各群落代表性鱼类均不同。由于大小鱼山春季NMDS的胁强系数为0.01,Stress<0.05,因此具有很好的代表性;秋季,胁强系数为0.08 位于0.05~0.1 之间,因此其结果基本可信[10]。经ANOSIM 检验可得,春季和秋季(R=0.566,P=0.001<0.05)呈现极显著差异。
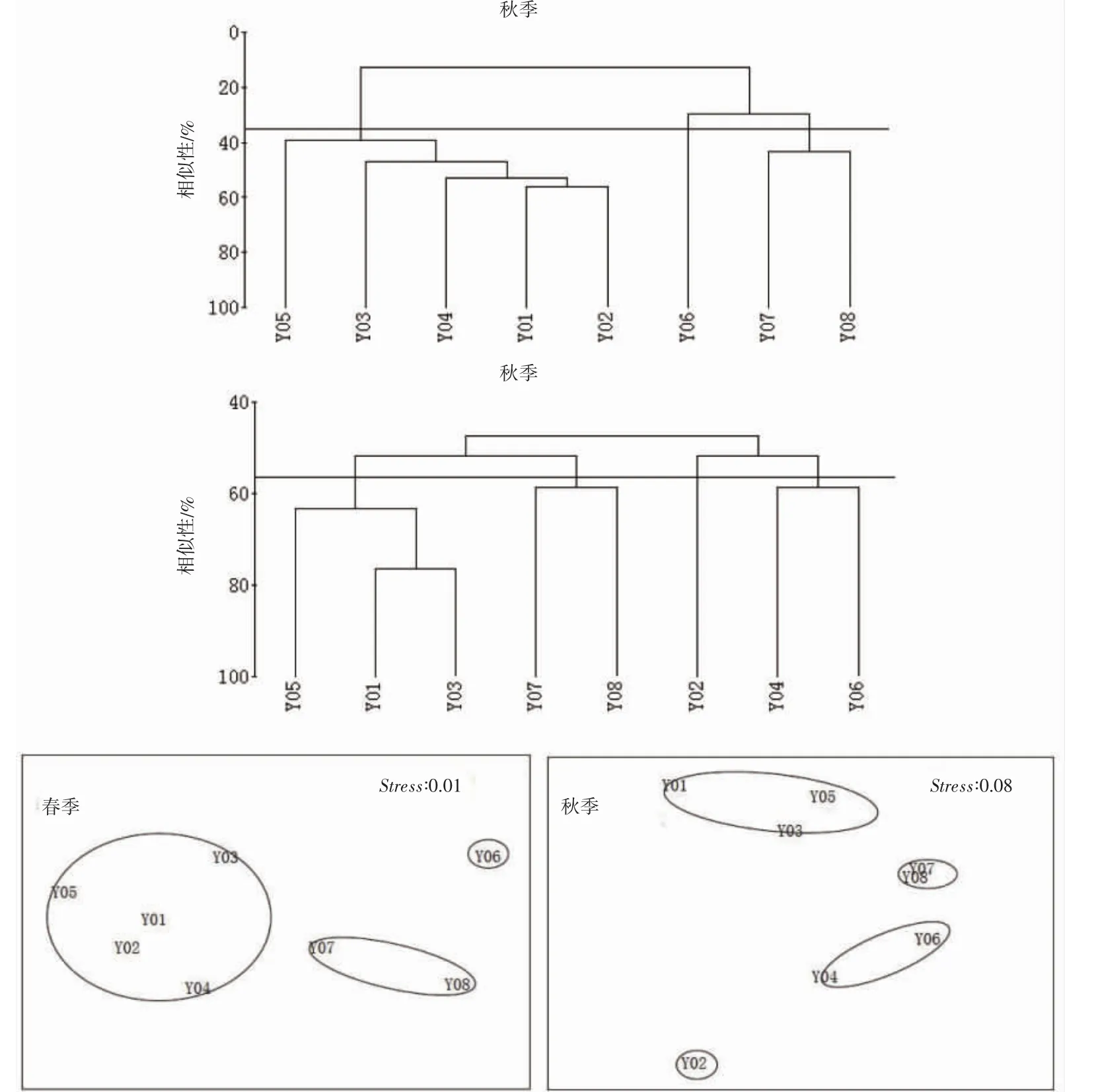
图5 鱼类群落CLUSTER 聚类图和NMDS 排序图Fig.5 CLUSTER analysis and NMDS analysis for fish assemblagein
表4 为春、秋季平均相似性、平均相似性累积贡献率之和大于90%的特征种和平均相异性、平均相异性的贡献率之和大于90%的鱼类分歧种。春季鱼类平均相似性为15.97%,特征种有凤鲚、褐鲳鲉Sebastiscus marmoratus、六丝钝尾虾虎鱼和孔虾虎鱼,它们对组内平均相似性贡献率之和达91.43%,其中以凤鲚的贡献率最高,为74.55%;秋季鱼类平均相似性为48.71%,特征种仅有龙头鱼一种,对组内平均相似性贡献率高达94.82%。春、秋季的平均相异性为69.80%,主要分歧种有龙头鱼、凤鲚、横带髭鲷Hapalogenys mucronatus 和刀鲚,它们对季节间平均相异性贡献率之和达91.37%,以龙头鱼贡献率最高,为68.23%,其次是凤鲚,为14.52%,其它鱼类贡献率均相对较低。

表4 调查海域鱼类季节内相似性及季节间相异性贡献率(%)Tab.4 Intra-season similarity and inter-season dissimilarity contribution rate of fishes
2.7 ABC 曲线
调查海域春、秋季ABC 曲线如图6 所示。春季,大小鱼山附近海域丰富度曲线先是与生物量曲线重合,然后位于生物量曲线的下方,最后逐渐重合,W 统计值为正值(0.027);秋季,大小鱼山附近海域丰度曲线一直位于生物量曲线上方,最后逐渐靠近直至重合,W 统计值为负(-0.034)。春季生物量曲线和丰度曲线的起点均比秋季生物量曲线和丰度曲线的起点低,表明该海域春季鱼类优势度低于秋季。

图6 调查海域鱼类ABC 曲线以及W 统计值Fig.6 Investigate the ABC curve of fish in the sea and the W statistical value
3 讨论
3.1 种类组成
根据2019 年春秋两季的调查渔获物,共鉴定出大小鱼山附近海域鱼类有33 种,其中春季有23 种,秋季有19 种,春季鱼类种类数多于秋季。该结果与邓小艳等[2]报道的2015 年舟山近岸渔场春季鱼类种类数(58 种)大于秋季(53 种)的结论一致,出现这种结果的原因可能是该调查海域属于河口性区域,营养盐充足,饵料生物丰富,是很多生物的繁殖、索饵场所,春季随着水温升高,鱼类性腺发育成熟,很多鱼类洄游到此进行产卵繁殖、索饵生长。同时,由于调查海域地处河口区,到这里生栖的鱼类多数属于季节性洄游鱼类,秋季,随着水温下降,一些鱼类逐渐向外侧深水区移动,也有一些鱼类逐渐开展季节性的越冬洄游,离开调查海域,从而导致秋季的鱼类种类数少于春季。
3.2 优势种分析
2019 年春季优势种仅有凤鲚1 种,秋季仅有龙头鱼1 种,优势种季节更替显著。其中,凤鲚多生活在沿岸浅水区或者近海,调查海域是凤鲚的产卵场和重要的索饵、栖息地[15],因此,春季,调查海域的凤鲚鱼类数量较高。龙头鱼属于杂食、广温广盐鱼类,对环境的适应能力较强,大多栖息于浅海底层水域,经常至河口水域觅食[16],也可能是因为该调查采用底拖网方式,渔获物中底层鱼类较多,所以秋季龙头鱼生物量较高。与俞存根等[2]、刘惠等[17]、何贤保等[18]对舟山渔场相关海域的鱼类优势种报道结果相比,优势种更替变化明显,呈现由大个体、高值向小个体、低值鱼类演替,这与其他学者研究结果相一致[19-20],出现这种现象的主要原因可能是过度捕捞引起的,其次也可能与环境污染导致海洋生境发生变化有关。
3.3 数量分布
根据调查结果,大小鱼山附近海域各站位鱼类种类数差异较大,春季,以秀山北部海域和大鱼山岛北部海域鱼类种类数较多,以调查海域南部和盘峙岛附近海域鱼类种类数较少;秋季,以调查海域东北部鱼类种类数较高,以调查海域南部鱼类种类数较低。从鱼类生物量来看,春季鱼类重量密度较高的站位(Y01、Y03)和秋季生物量密度较低的站位(Y01、Y02、Y03)主要分布在调查海域西北部的开阔海域,春季重量密度较低的站位(Y06、Y07、Y08)和秋季生物量密度较高的站位(Y06、Y07)主要分布在近岛屿侧的海域。这可能是由于调查海域北部以及西北部属于相对开阔海域,距离岛屿相对较远,污染、养殖活动以及跨海大桥等人为干扰相对较弱,因此,该海域的鱼类种类数比南部海域多,也可能是与海底地形、水深不同有关。具体原因尚有待进一步分析研究。
3.4 生物多样性分析
对比春秋季各站位的生物多样性指数得知,大小鱼山附近海域不同站位鱼类多样性差异较大且均匀度相对较低,梅春等[21]认为出现这种情况是优势种种类较少,导致最大优势种比例较大,其他物种占比较少,进而引起均匀度偏低。同时,春季鱼类生物多样性指数(H')、均匀性指数(J')、丰富度指数(d)均高于秋季。
3.5 群落聚类分析
舟山渔场大小鱼山附近海域秋季物种平均相似性高春季,春季在生物量和种类水平上均比秋季低,这两个季节相似性贡献率最高的鱼类特征种分别为凤鲚和龙头鱼,而春、秋季物种相异性贡献率最高的鱼类分歧种是孔虾虎鱼,这可能是因为春秋季捕获的鱼类数量主要集中于个别种,该种类数量占比较高。调查海域春秋季鱼类群落分别可分为4、3 个群组,不同组群或者说鱼类群落结构空间差异主要是由群组间鱼类种类组成不同和重要种及优势种不同造成的。
春季,调查海域丰富度曲线先是与生物量曲线重合,然后位于生物量曲线的下方,并且W 值为正数,表明春季调查海域鱼类群落结构尚未受到干扰,也显示调查海域大个体的鱼类占主体[23],秋季,丰度曲线一直位于生物量曲线的上方,其W 值为负数,表明秋季该海域鱼类群落结构受到严重干扰,该海域小个体鱼类占主体。春、秋季丰度和生物量曲线的起点都高于60%,表明某一种鱼类优势较为显著,这个特征与春季唯一的优势种凤鲚和秋季唯一的龙头鱼相吻合;春秋季丰度和生物量曲线均相距很近,表明一些机会鱼类的数量开始增加,这也预示了调查海域污染或者干扰的程度会加大[24],这是否可能反映了调查海域因人类活动造成海洋环境恶化的趋势问题。尚有待于进一步深入探讨。
