不同埋土深度杉木无性系萌蘖及内源激素含量的差异
2021-10-21张吉玲叶义全林思祖曹光球
张吉玲 叶义全,2 黄 樱 李 勇 郑 宏 林思祖,2 曹光球,2
(1. 福建农林大学林学院,福建 福州 350002;2. 国家林业和草原杉木工程技术研究中心,福建 福州 350002;3. 福建省洋口国有林场,福建 南平 353200)
杉木(Cunninghamia lanceolata)是我国南方最重要的速生用材树种之一,也是萌芽能力较强的一个树种[1]。自20世纪90年代以来,杉木无性繁殖技术日趋成熟,杉木无性系造林面积日益扩大。营林生产实践表明,杉木组培苗萌蘖能力普遍高于扦插苗和实生苗。杉木组培苗萌蘖不仅会提高幼林抚育成本,而且还会影响幼林生长,这在一定程度上限制了杉木组培苗的推广应用。迄今为止,人们对杉木组培苗的研究更注重于组培配方的优化,而忽略了组培苗在造林过程中出现的各种问题[2]。
内源激素是由植物自身代谢产生的一类有机物质,是植物细胞接受特定环境信号诱导产生的、低浓度时可调节植物生理反应的活性物质。内源激素对植物细胞的分裂[3]、分化[4-5].、器官建成以及植物的休眠[6]和形态反应[7]等均有直接或间接的调节控制作用。近些年来,随着植物内源激素研究越来越广泛,植物内源激素的合成机理以及作用机理已成为植物生理学研究的热点之一。诸多研究结果表明,植物内源激素与萌蘖能力密切相关[8-10]。
埋土是农业生产中常用的一种措施。埋土深度不同,不仅影响实验对象生长微环境的温度,而且还会影响实验对象生长微环境的湿度。目前,不同埋土浓度研究主要集中在种子发芽能力的研究[11]。林业生产实践表明,不同埋土深度会影响杉木无性系的萌蘖能力。然而,不同埋土深度影响杉木无性系萌蘖能力的机理至今尚未清楚。有鉴于此,本研究以萌蘖能力较强的杉木‘洋020’组培苗及萌蘖能力较弱的杉木‘洋020’扦插苗为研究对象,通过室内盆栽实验,分析不同埋土深度处理对无性系萌蘖能力及不同器官内源激素含量的影响,研究结果为揭示杉木萌蘖机理以及杉木人工林高效培育提供参考依据。
1 材料与方法
1.1 试验材料
供试材料为福建省洋口国有林场(117°92′ E,26°87 N)提供的1年生杉木‘洋020’扦插苗和‘洋020’组培苗。挑选生长健壮、长势一致、无病虫害的苗木为试验材料。‘洋020’扦插苗平均苗高30 cm,地径为5 mm;‘洋020’组培苗平均苗高18 cm,地径为2.6 mm。培养基质为黄心土,黄心土有机质含量49.00 g/kg,全氮1.09 g/kg,水解性氮l.32 mg/kg,速效磷2.30 mg/kg,速效钾151.30 mg/kg。
1.2 试验方法
1.2.1 试验设置
本试验在福建农林大学国家林业和草原杉木工程技术研究中心田间实验室(26°5′N,119°13′E)内盆栽。于2018年4月13日将苗木种植于花盆中,每盆种植3株,花盆规格为21 cm×26 cm×27 cm。埋土深度分3个处理,分别为埋土深度3、6、9 cm。每个处理种植30盆,共90盆。盆栽后每月中旬调查不同处理杉木基部的萌芽数。在萌蘖高峰期8月份中旬,分别采集已萌芽扦插苗及组培苗第2轮健康新生枝叶、萌芽处韧皮部(萌芽处切0.5 cm×0.2 cm小块韧皮部)及根尖样品。每个处理采集9株,每3株混成1个待测样,每个器官待测样3个重复。不同器官样品称量后迅速放入液氮中,置于-80 ℃超低温冰箱中保存待测。培养期间定期给苗木浇自来水,保持土壤含水率在65%左右,定期除草;9月17日结束实验。
1.2.2 萌蘖数调查
2018年4月份至8月份期间,每月中旬调查不同处理的萌蘖数,以芽露出培养基质表面为萌芽条。
1.2.3 内源激素含量测定
采用酶联免疫吸附法(ELISA)测定杉木枝叶、萌芽韧皮部和根尖内源激素吲哚乙酸(IAA)、赤霉素(GA3)、脱落酸(ABA)和细胞分裂素(CTK)含量[12-13],各激素试剂盒由上海科兴生物有限公司提供。
1.3 数据处理
试验数据用Excel 2010和SPSS 22.0软件进行数据和相关性分析。
2 结果与分析
2.1 不同埋土深度杉木无性系萌蘖差异
由表1可知,随着培养时间的延长,扦插苗及组培苗萌蘖能力呈现先增强后降低的趋势,在8月时,2种无性系的萌蘖能力最强。随着埋土深度的增加,同一杉木无性系萌蘖能力均呈逐渐下降的趋势。就不同类型无性系萌蘖能力差异而言,在相同条件下,组培苗萌蘖能力均强于扦插苗;在萌蘖高峰期8月,埋土深度3、6、9 cm组培苗的萌蘖数与同一埋土深度扦插苗相比分别提高11.15%、4.76%及3.45%。在萌蘖高峰期8月,同埋土深度同一类型无性系萌蘖能力差异如下,培苗埋土深度9 cm处理的萌蘖数分别比埋土深度6 cm及3 cm处理的降低4.54%和27.34%,扦插苗埋土深度9 cm处理的萌蘖数分别比埋土深度6 cm及3 cm处理的降低3.33%和21.92%。方差分析结果表明,扦插苗及组培苗萌蘖能力差异均为显著(P<0.05)。在萌蘖6月份,组培苗埋土深度3 cm处理平均萌蘖数与埋土深度6 cm及9 cm处理的差异达到显著水平(P<0.05);在萌蘖7月份,组培苗不同埋土深度处理的平均萌蘖数两两差异达显著水平(P<0.05);在萌蘖8月份,组培苗埋土深度9 cm处理的平均萌蘖数与埋土深度3 cm及6 cm处理的差异达到显著水平(P<0.05)。

表1 不同埋土深度、不同繁殖方式、不同时期杉木无性系萌蘖差异Table 1 Tillering differences of cuttings and plantlets of C. lanceolata clones at different time in different soil depths
2.2 不同埋土深度杉木无性系不同器官内源激素含量
2.2.1 不同埋土深度扦插苗不同器官内源激素含量
由图1可知,随着埋土深度增加,杉木枝叶及韧皮部IAA含量呈逐渐上升趋势,枝叶及基部韧皮部GA3含量、基部韧皮部ABA含量呈逐渐下降趋势,根尖IAA含量呈先下降后上升的趋势,根尖GA3含量、枝叶及根尖ABA含量则表现为先上升后下降的趋势。随着埋土深度的增加,枝叶的CTK含量表现为先下降后上升的趋势,基部韧皮部含量表现为先上升后下降的趋势,而根尖含量则表现为逐渐上升的趋势。随着埋土深度的增加,各器官内源激素含量呈现出较复杂的变化规律且差异性显著。同一埋土深度不同器官IAA含量体现为枝叶>基部韧皮部>根尖;埋土深度3 cm处理的不同器官GA3含量表现为基部韧皮部>枝叶>根尖,ABA含量表现为基部韧皮部>根尖>枝叶,CTK含量表现为枝叶>基部韧皮部>根尖。埋土深度6 cm处理的不同器官IAA含量表现为枝叶>根尖>基部韧皮部,GA3含量表现为根尖>基部韧皮部>枝叶,ABA含量表现为基部韧皮部>枝叶>根尖。埋土深度9 cm处理的不同器官ABA含量表现为基部韧皮部>枝叶>根尖,ABA含量则表现为枝叶>根尖>基部韧皮部。
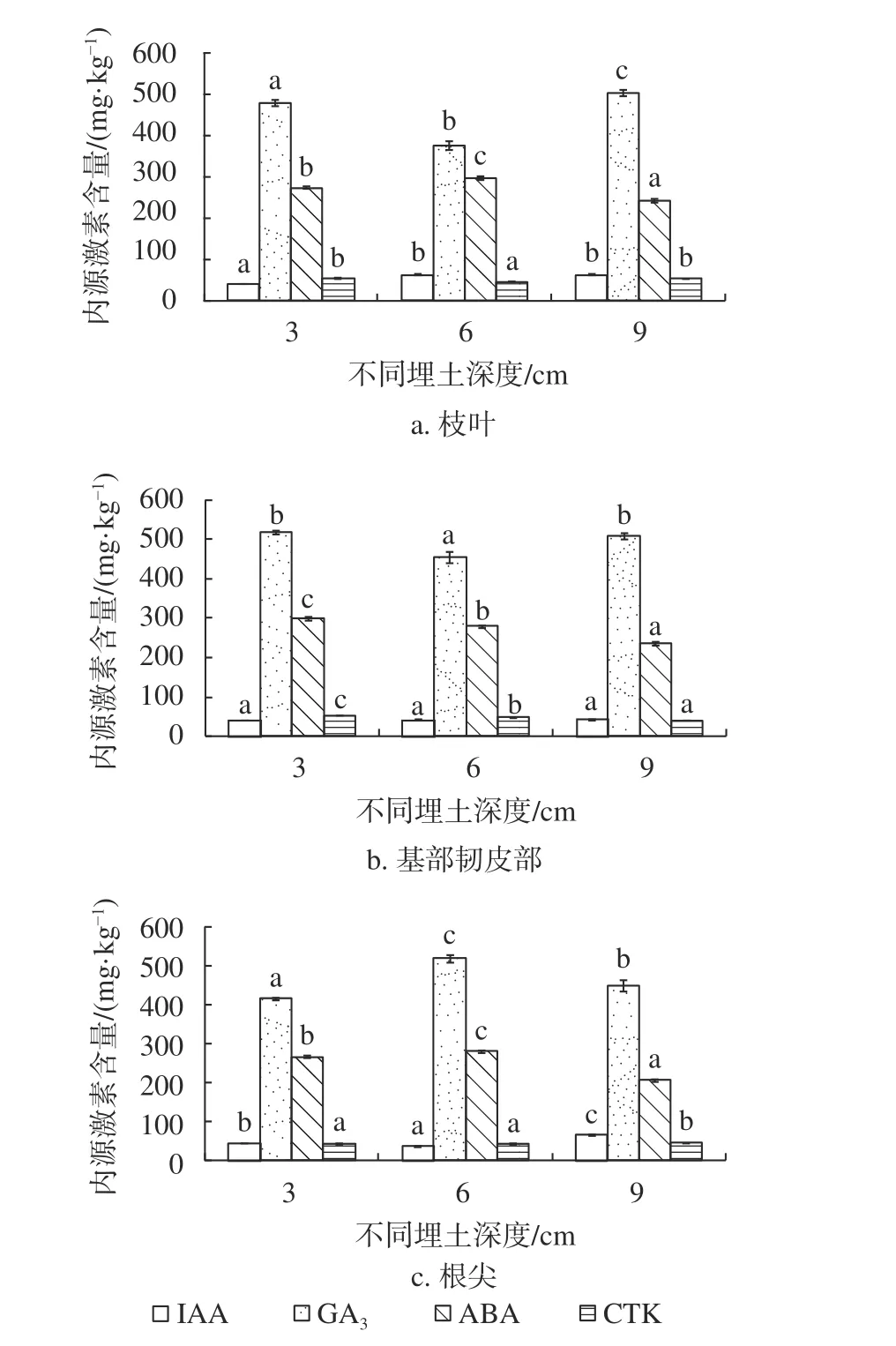
图1 不同埋土深度扦插苗器官内源激素含量Fig. 1 Endogenous hormone contents in different organs of C. lanceolata'Yang 020' cuttings in different soil depths
2.2.2不同埋土深度组培苗不同器官内源激素含量
由图2可知,随着埋土深度的增加,杉木枝叶及基部韧皮部IAA呈先上升后下降的趋势,根尖IAA呈逐渐上升的趋势;杉木枝叶及基部韧皮部GA3呈逐渐下降的趋势,根尖GA3呈先上升后下降的趋势;杉木枝叶ABA含量呈逐渐上升的趋势,基部韧皮部和根尖ABA含量呈先下降后上升的趋势;枝叶和根尖CTK含量呈先下降后上升的趋势,而基部韧皮部CTK含量则呈先上升后下降的趋势。同一埋土深度组培苗不同器官内源激素分配,随埋土深度处理呈现不同的分配规律且差异性显著。埋土深度3 cm处理的IAA含量体现为基部韧皮部>根尖>枝叶,GA3含量体现为基部韧皮部>根尖>枝叶,ABA含量体现为基部韧皮部>根尖>枝叶。埋土深度6 cm及9 cm处理的IAA含量均体现为枝叶>基部韧皮部>根尖,GA3含量均体现为根尖>枝叶>基部韧皮部,ABA含量均体现为基部韧皮部>枝叶>根尖。埋土深度3 cm及6 cm处理的CTK含量体现为枝叶>根尖>基部韧皮部,埋土深度9 cm处理的CTK含量均体现为根尖>枝叶>基部韧皮部。
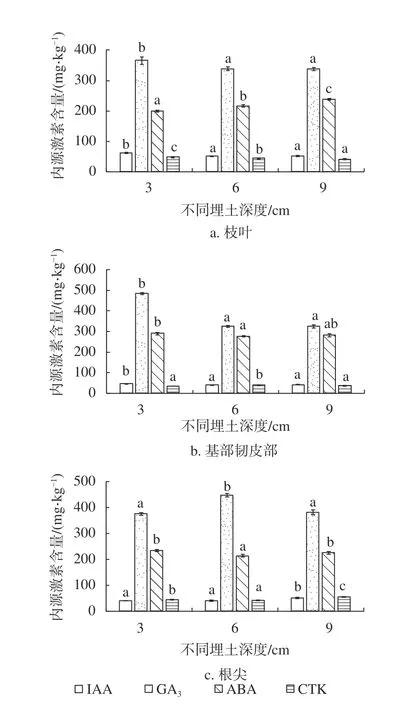
图2 不同埋土深度组培苗器官内源激素含量Fig. 2 Endogenous hormone contents in different organs of C. lanceolata'Yang 020' plantlets in different soil depths
2.3 不同埋土深度杉木无性系相关性分析
由表2可知,萌蘖数与繁殖方式呈极显著正相关(P<0.01),与GA3含量呈显著正相关(P<0.05),而与埋土深度呈极显著负相关(P<0.01)。IAA含量与埋土深度呈显著正相关(P<0.05),与器官部位呈显著负相关(P<0.05);GA3含量与繁殖方式呈极显著负相关(P<0.01),与器官部位和萌蘖数显著正相关(P<0.05);ABA含量与繁殖方式和IAA含呈呈极显著负相关(P<0.01);CTK含量与繁殖方式呈显著负相关(P<0.01)。
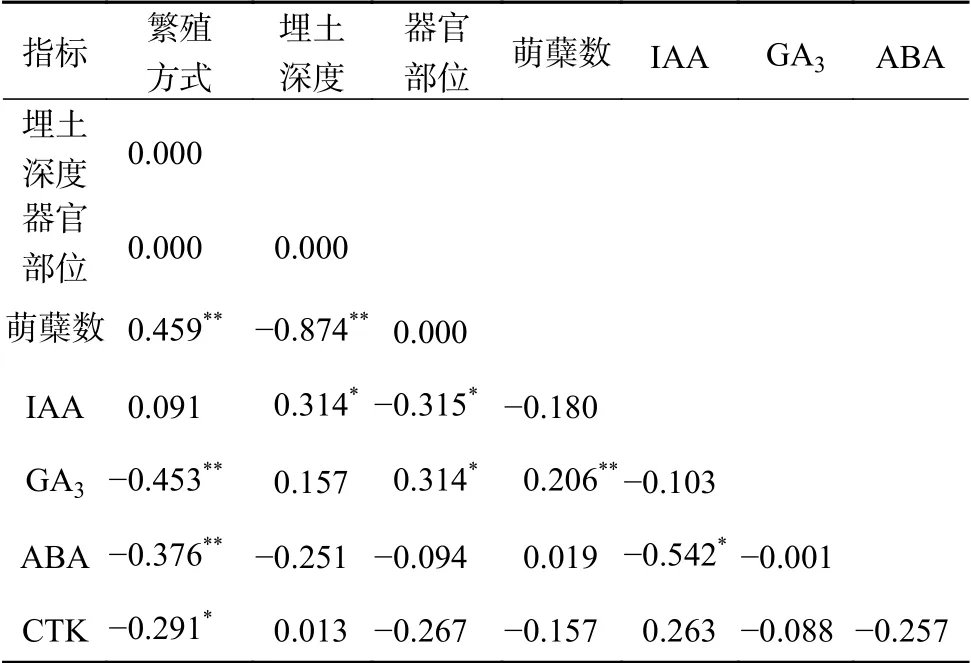
表2 不同埋土深度条件下杉木无性系萌蘖相关性Table 2 Correlation of tillering among C. lanceolata clones in different soil depths
3 结论与讨论
植物是有机体,由细胞、组织和器官组成,其正常的生长发育需要水分、矿质和有机物质的供应,其自身维持正常状况以适应外界环境是通过信息交流来完成的,这种信息交流的化学信使就是植物激素[14]。植物激素不是由特定器官产生,不同器官产生的一种或多种激素可以通过运输在一定的部位发挥作用,其运输速度和方向因激素种类不同而存在差异。内源激素含量在植物不同发育期以及不同器官之间均存在较大差异[15]。本研究结果表明,同一无性系不同器官以及同一器官不同生长期的内源激素含量差异较大。
植物内源激素的产生受环境因子的影响,激素间通过协调作用从而维持植物的正常生长发育和分化[16]。植物内源激素与植物生理过程密切相关。生长素、赤霉素及细胞分裂素被认为对植物的萌蘖特性发挥着重要的调控作用[17-18]。脱落酸的作用与细胞分裂素相反,被认为是植物对不同逆境产生响应的信号因子,不仅能抑制植物的生长,还对其他生长促进型激素产生拮抗作用[19]。本研究结果表明,在8月份时,扦插苗和组培苗的萌蘖能力最强。随着埋土深度的增加,同一种杉木无性系萌蘖能力均呈逐渐下降的趋势。植物萌蘖受到植物激素、温度和埋土深度等不同环境的综合调控,其中植物内源激素在调控植物生长发育过程中起关键作用。有研究表明,杉木伐桩休眠芽的萌发与温度有密切关系。温度升高,导致萌芽量下降。温度条件影响激素的合成,制约伐桩萌芽能力,而激素的供应与平衡又影响到伐桩萌芽数量[20]。这说明埋土深度越深,温度也随之升高,即对杉木萌蘖有抑制作用,这与本研究结果一致。从杉木萌芽中提取分离天然激素,经多次试验测得提取液中生长素和抑制物质的含量很低,而含有较多的赤霉素类和细胞分裂素类物质[21]。这意味着杉木萌蘖应与GA3含量有较大的关系。杉木萌蘖数与埋土深度呈极显著负相关,而与GA3含量呈极显著正相关,而与其他内源激素无显著相关性,这与前人的研究结果存在类似结论。但是GA3在杉木的生长发育中是否也起作用,或是在杉木的萌蘖和生长期间才转变为活性态,尚待进一步探讨。
本研究结果表明,随着埋土深度的增加,2个无性系苗萌蘖能力均呈降低的趋势,且不同埋土深度处理会影响植物的内源激素的合成。随着埋土深度的增加,杉木幼苗枝叶IAA含量呈上升趋势,GA3含量呈下降趋势;埋土深度6 cm处理有利于枝叶及根尖ABA的积累;埋土深度9 cm处理有利于根尖CTK的积累。杉木幼苗不同器官植物内源激素的含量差异应与杉木对埋土深度处理的生理应答机制有关。不同种类的内源激素在植物生长发育过程中有不同的作用,且不同种类之间有交互作用。一种激素不仅可以参与多个生理过程,而且可在某种生理过程通过诱导另一种激素的合成和释放来诱导这一生理反应[22-24]。本研究结果在一定程度上反映不同埋土深度杉木无性系萌蘖内源激素IAA、GA3、ABA、CTK之间的含量变化规律,至于在杉木萌蘖过程中内源激素之间如何维持平衡,以及如何协同作用调控杉木萌蘖等的机制尚不清楚,这有待于今后从生理生化、分子生物学等方面进一步深入研究。
