斑块状海草床对大型底栖动物群落的影响
2021-10-20孟周严润玄韩庆喜
孟周,严润玄,韩庆喜
(宁波大学 海洋学院,浙江 宁波 315832)
海草床主要由大叶藻构成(韩秋影等,2008;Short et al,2007),是鱼类、贝类、甲壳类等多种海洋生物栖息、觅食和育幼的重要场所(Newell,2001)。大叶藻对于稳定海底基质的物理化学性质,缓冲潮流对海底沉积物的扰动,促进环境中重金属离子的循环,以及维护近岸水域生态系统中较高的生物多样性等均起到了重要作用(Leopardas et al,2014)。海草床大型底栖动物通过挖洞、摄食、潜沙等活动改造和扰动海草床,在碎屑为基础的海草食物网中发挥着重要的生态功能,是初级生产者和更高营养级的捕食者之间能量传递和转移的中间桥梁和纽带(Valentine et al,1993)。
在富营养化、疏浚以及暴风雨等人为或环境因素的影响下(Duarte,2002),生境破碎化和海草床斑块化日益严重(Horinouchi,2007)。海草床可为小型鱼类等动物提供躲避捕食者的避难所(Jackson et al,2001;Smith et al,2011),因而海草结构的改变会改变海草床的生态服务功能(Horinouchi,2007)。
大型底栖动物常用于生物资源评估和生态环境质量评价(周细平等,2020),以及近海污染和有机质累积的监测(Han et al,2013)。国内关于矮大叶藻海草床大型底栖动物的研究仅限于广西合浦矮大叶藻海草床大型底栖动物群落(张景平等,2011)和山东荣成复合海草床大型底栖贝类的调查研究(刘建影等,2017),而在海南陵水县新村港与黎安港海草床的研究并不涉及矮大叶藻(涂志刚等,2016)。上述研究发现,这些未受干扰的海草床具备完善的生态功能和良好的生态状况。矮大叶藻在山东半岛的莱州湾地区和广饶地区呈零星斑块状分布,且面临着正在开挖海参池的威胁(Orth et al,2006;刘坤等,2012),海草床明显退化,斑块化现象严重。因此,对斑块化的矮大叶藻海草床大型底栖动物群落进行研究,以评估退化海草床在近岸海域潮间带生态系统中的生态服务功能状况势在必行。
本文根据山东广饶斑块状海草床区域大型底栖动物的调查数据和资料,对山东广饶海草床内外大型底栖动物群落组成及结构的差异进行研究,并与环境因子的相关性分析相结合,揭示斑块状海草床对大型底栖动物群落组成和结构的影响,旨在为矮大叶藻衰退海草床的生态服务功能评价提供数据基础,为未来矮大叶藻海草床的监测和恢复提供科学依据。
1 材料和方法
1.1 调查海域
本次调查的时间为2013 年5 月,调查采样点位于山东东营广饶县境内,莱州湾西部,黄河入海口以南,具体位置为37毅20忆46.4义N,118毅57忆16.9义E,采样海域的平均温度(本次调查的5 月份) 为24.84 益,平均pH 值为7.36,平均盐度为22.09,海水溶解氧的含量为9.02 mg/L。调查站位地理位置如图1 所示。
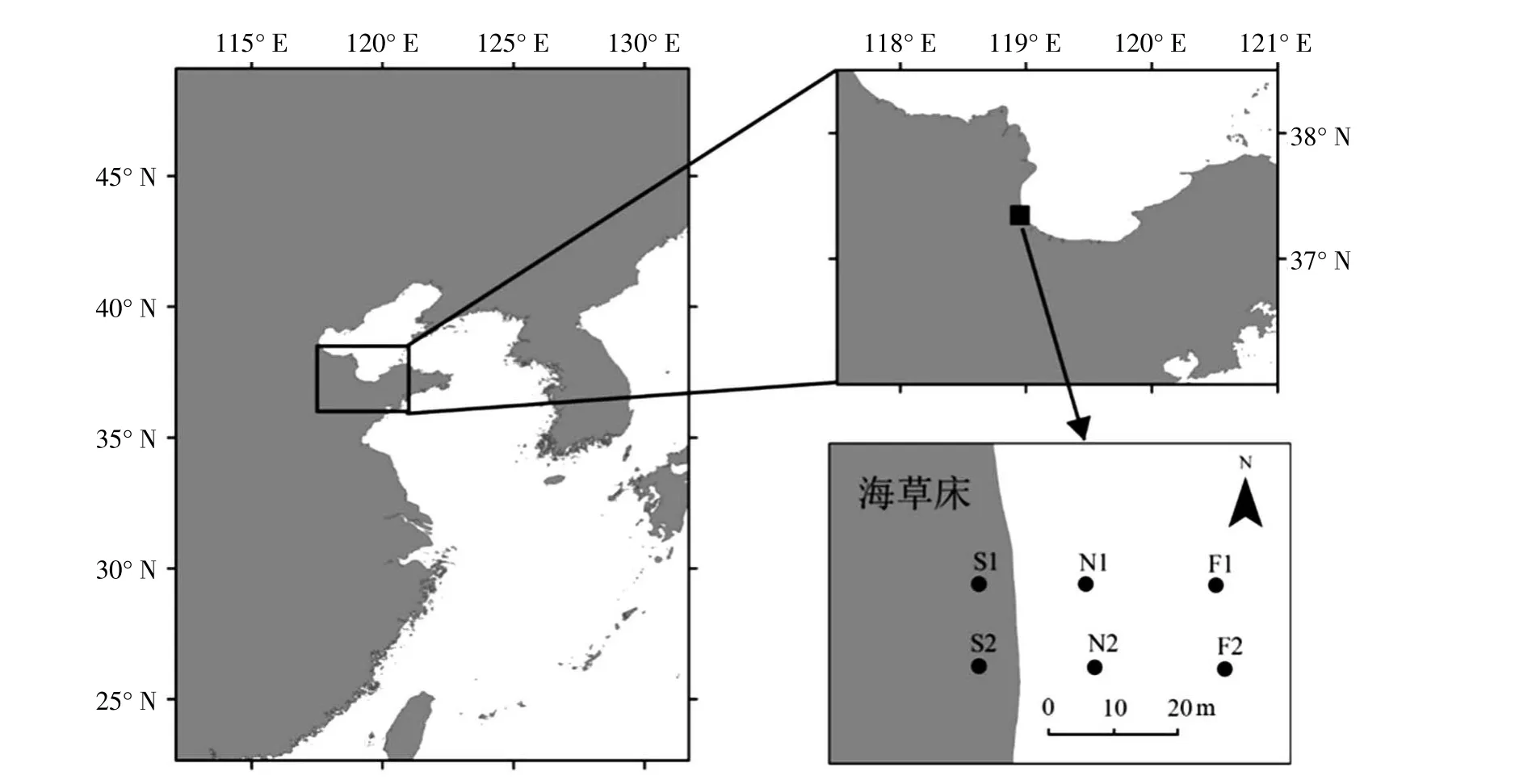
图1 2013 年5 月山东广饶大型底栖动物采样站位示意图
1.2 采样方法
本次采样区域位于山东省东营市广饶县滨海潮间带海草床及附近海域,调查选取2 条断面,用GPS 定位仪定位记录经纬度,以较大面积的斑块状分布的海草床为采样起点,并沿海草床边缘向海面方向延伸10 m 和30 m 取样,即海草床内部站位(S1 和S2)、距离海草床10 m 区域(N1 和N2)和距离海草床30 m 区域(F1 和F2)。采集的样品包括底质样品和大型底栖动物。采集时用0.5 m 伊0.5 m的取样框,对0.25 m2内所有样品进行收集,后用0.5 mm 网目的筛子进行冲洗,所得样品用95%的酒精进行固定后带回实验室。在实验室用1%的虎红酒精溶液对酒精固定样品进行染色,将大型底栖动物拣出。拣出的生物样品在解剖镜下进行鉴定,部分生物特征如多毛类刚毛等辅以显微镜观察,尽量鉴定到种。分类计数后的标本用精度为1 mg 的电子天平称重;并换算求得每个物种的优势度、生物量(g/m2)和丰度(ind/m2)。样品的冲洗、鉴定和称重等均依照《海洋调查规范》 (GB 12763.6—91)进行。同时,对同步获取的底质粒径组成进行仪器分析。调查规范依据《海洋调查规范》第6 部分:海洋生物调查进行(国家海洋局,2007)。
1.3 多样性指数和优势种
依据大型底栖动物群落的丰度数据计算群落的丰度(A)、物种数目(S)、香农威纳多样性指数(H忆;log10)、均匀度指数(J忆)以及物种丰富度指数(d),以上指数均采用PRIMER v6.0 的diverse程序组进行计算。
优势种的计算:优势种的定义在不同资料、文献中有所区别,本文通过优势度Y 来计算,以优势度Y > 0.02 作为判定优势种的标准(Pinkas et al,1971)。其计算公式为:
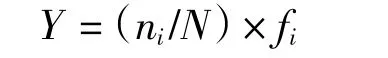
式中,N 为底栖群落的个体数目总和,ni为第i 种的个体数;fi为该物种的出现频率。
1.4 功能群的划分
功能群依据大型底栖动物的摄食特性和食性划分(李少文等,2013;葛宝明 等,2008;Lin et al,2018),包括:
(1)浮游生物食者:以滤食水体中浮游动植物和有机质颗粒为主,包括藤壶、大部分双壳类和少数腹足类。
(2)肉食者:猎食其他动物,如某些环节动物、十足类等。
(3)杂食者:取食植物藻类残体和小型动物,如某些腹足纲、双壳纲和蟹类等。
(4)碎屑食者:通过吞食沉积物获取有机物质,如某些多毛类、双壳类和海参等。
(5)植食者:摄食植物和海藻,如部分腹足纲种类和蟹类等。
1.5 粒径的分析与划分
粒径的测量采用了马尔文仪器有限公司的Mastersizer 2000 激光粒度仪测定,粒径测量范围为0.02~2 000 滋m,依据粒径的大小分为砂(64~2 000 滋m)、粗砂(16~64 滋m)、粉砂(4~16 滋m)和黏土(<4 滋m) (Folk et al,1957)4 种底质类型。
1.6 软件的使用
在本研究中,采用地理信息软件ArcGIS Desktop 10.6 绘制采样站位分布图;采用Microsoft Office 2010 进行优势度计算;采用多元统计软件PRIMER( Plymouth Routines in Multivariate Ecological Research)v6.0 进行大型底栖动物群落的聚类分析(Cluster)、非度量多维尺度分析(nonmetric multi-dimensional scaling,nMDS)以及用于检验群落结构沿环境压力梯度变化显著性的连续性检验(Seriation test)。采用PASW Statistics 18.0 中单因素方差分析(ANOVA)进行两组样品间差异显著性分析,分别以0.05 和0.01 作为极显著差异和显著差异的界值标准。
2 结果
2.1 大型底栖动物群落物种组成、丰度和优势种
2013 年5 月在山东省广饶县潮间带海草床及附近潮间带海域共捕获992 只大型底栖动物个体,分属3 门4 纲15 目28 科41 种(物种名录见表1)。甲壳动物的物种数量最多,包括4 目11 科16 种,占据该海域大型底栖动物群落物种总数的39.02%;多毛类动物的物种数量略少,包含4 目8 科15 种,占群落物种总数的36.59%;软体动物物种数目最少,含7 目9 科10 种,占群落物种总数的24.39%。在所有的41 种大型底栖动物中,有33 种在海草床区域出现,且有12 种仅在海草床区域出现,在远离海草床的站位均未采集到,包括7 种多毛类、2 种甲壳动物和3 种软体动物;在离海草床10 m 和30 m 处,仅采得大型底栖动物21 种和20 种,有8 个物种未在海草床附近出现,包括多毛类动物1 种、软体动物1 种和甲壳动物6 种。

表1 山东广饶海草床及附近海域大型底栖动物物种名录及功能群划分
本研究中大型底栖动物的平均群落丰度为661.33 ind/m2。丰度在各动物门类中,以软体动物所占丰度比例最大,为51.71%;多毛类次之,占33.27%;甲壳动物较少,占15.02%。丰度的空间分布具有显著差异,海草床区的群落动物丰度高达1 390.00 ind/m2,高于距海草边缘10 m 站位的226.00 ind/m2和30 m 站位的368.00 ind/m2。所有物种中,光滑河篮蛤和丝异须虫是大型底栖动物群落丰度的主要贡献者,两者的丰度分别占群落总丰度的43.15%和17.94%。以空间分布为可变因素进行各个物种丰度的单因素ANOVA 分析,结果显示丝异须虫存在显著的空间分布差异(P=0.03<0.05),沿海草床向外丰度值显著降低;色斑角吻沙蚕亦是群落丰度的重要贡献者,其丰度亦表现出显著的空间分布差异(P=0.036<0.05),在海草床区的出现频度为100%,且丰度值远高于非海草床站位,该种在非海草床区域的出现频率仅25%,且丰度值也显著较低。
浮游生物食者是该海域大型底栖动物功能群的主要组成部分,其丰度占群落总丰度的49.40%;碎屑食者次之,其丰度占总丰度的37.10%;肉食者和杂食者较少,分别仅占7.96%和5.54%。对各个功能群丰度按距离梯度进行单因素ANOVA 分析,结果显示肉食者的空间分布存在显著性差异(P=0.016<0.05),Duncan 检验显示海草床内的肉食者丰度显著高于海草床外10 m 和30 m 处;其他三种功能群,如杂食者、浮游生物食者和碎屑食者的丰度均未表现出统计学意义上的显著的空间分布差异。以距离为单因素,各个大型底栖动物类群的丰度ANOVA 分析结果显示,甲壳动物、软体动物和多毛类动物的丰度在沿海草床向外部延伸的距离梯度上并无显著差异,即没有显著的空间分布差异。
该海域共有大型底栖动物优势种7 种(表2),包括多毛类的丝异须虫、奇异稚齿虫、色斑角吻沙蚕,甲壳动物的三叶针尾涟虫和日本拟背尾水虱,以及软体动物的光滑河篮蛤和红明樱蛤。优势种中以色斑角吻沙蚕的出现频率最低,仅66.67%,主要出现于海草床区域;光滑河篮蛤、丝异须虫和日本拟背尾水虱在所有采样站位都有出现,出现频度为100%,其密度在海草床区域均高于海草床外围。
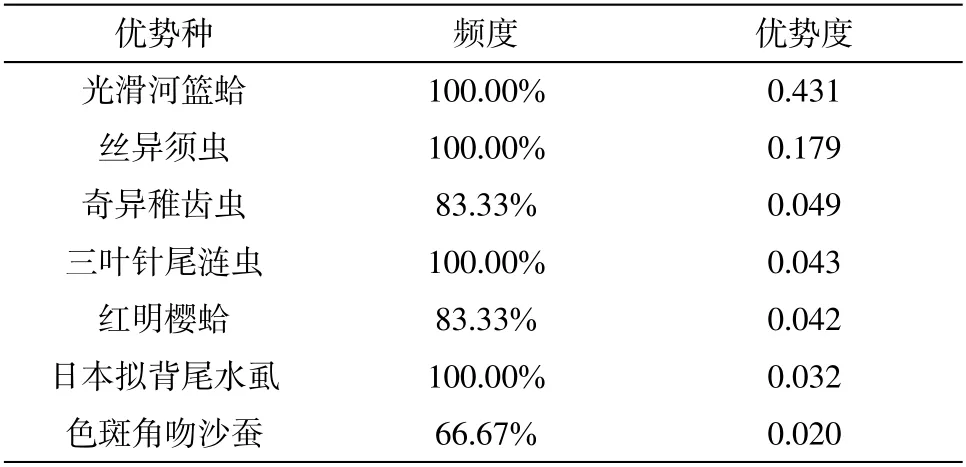
表2 2013 年5 月山东广饶海草床大型底栖动物优势种
2.2 相关性分析
在本研究采样区域,按距离梯度从海草床向外部延伸,进行了底质的采样以及后续的实验室粒径测量分析,具体站位的底质粒径分布状况如表3 所示。粒径共分为4 个粒级,分别为小于4.00 滋m 的黏土、4.00 ~ 16.00 滋m 的粉砂、16.00 ~ 64.00 滋m的粗砂以及64.00~2 000 滋m 的砂质。以粒度百分比和中值粒径(Md劓)为变量,以距离为可变因子的单因素ANOVA 分析结果显示,各组在距离梯度上均呈现显著差异。对于小于4.00 滋m 的黏土组分来说,10 m 组>30 m 组,且差异显著(P=0.048<0.05);对于4.00~16.00 滋m 的粉砂组分来说,10 m组>海草床组>30 m 组,且差异显著(P=0.011<0.05);对于16.00~64.00 滋m 的粗砂组分来说,30 m组低于10 m 组和海草床组,且差异显著(P =0.036<0.05);对于64.00~ 2 000 滋m 的砂质组份来说,与粗砂组分相反,30 m 组高于10 m 组和海草床组,且差异显著(P=0.036<0.05);中值粒径Md劓的分析结果与砂质相同,30 m 组高于10 m 组和海草床组,且差异异常显著(P=0.009<0.01)。较粗粒径的粗砂和砂来说,30 m 组的较粗粒径组成百分比显著大于海草床组和10 m 组。

表3 山东省广饶县海草床及附近海域的粒径分布状况
对大型底栖动物群落参数与沉积物粒径进行相关性分析,结果显示,仅物种数目与底质粒径中的粗砂成分呈密切的正相关关系(表4)。

表4 2013 年5 月山东广饶海草床大型底栖动物与底质粒径相关性分析
2.3 大型底栖动物群落结构分析
大型底栖动物的群落丰度数据先经过平方根转换和标准化处理,后转化为Bray-Curtis 相似性矩阵,再进行聚类分析(Cluster)和非度量多尺度分析(nMDS),并将Cluster 分析结果与MDS 排序结果叠加,基于聚类分析和非度量多尺度分析,在合理的相似性水平(一般超过20%)下将大型底栖动物划分成数量合适的不同组(刘迅等,2017)。如图2,在60%的相似性水平,可将研究区域的大型底栖动物群落分为4 个组,分别是F2、S1、S2和F1,以及N1 和N2。
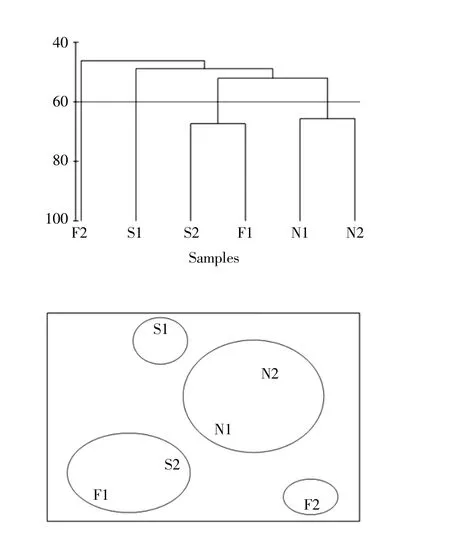
图2 山东省广饶县海草床大型底栖动物群落的聚类分析和nMDS 排序
大型底栖动物群落的SIMPER 分析结果显示,海草床内(S1 和S2)大型底栖动物群落的平均相似度为50.00%,主要贡献者为丝异须虫(贡献率27.47%)和光滑河篮蛤(贡献率23.56%);距海草床10 m 处(N1 和N2)的平均相似度为65.67%,主要贡献者是丝异须虫(贡献率19.03%)和红明樱蛤(贡献率13.05%);距海草床30 m 处(F1 和F2)的平均相似度为41.25%,主要贡献者是光滑河篮蛤(贡献率42.35%) 和丝异须虫(贡献率19.74%)。
海草床区域(S1 和S2) 和10 m(N1 和N2)区域的大型底栖动物群落的平均不相似性为45.29%,主要贡献者是光滑河篮蛤(贡献率42.35%)和小头虫(贡献率19.74%);海草床区域(S1 和S2)和30 m(F1 和F2)区域的大型底栖动物群落的平均不相似性为49.08%,主要贡献者亦为光滑河篮蛤和小头虫,但其贡献率分别仅为9.52%和6.42%;海草床外10 m 区域(N1 和N2)和30 m(F1 和F2)区域的大型底栖动物群落的平均不相似性为52.90%,主要贡献者是光滑河篮蛤(贡献率14.44%)和红明樱蛤(贡献率7.59%)。
运用PRIMER6.0 软件中的连续性检验(Seriation test)来检验群落结构沿海草床距离梯度上的连续性,检验结果Rho=0.031,P=0.476 >0.05。因此,在沿海草床向外延伸的距离梯度上,并无可见的线性关系。
3 讨论
3.1 矮大叶藻的分布及生态重要性
矮大叶藻(Zostera japonica),也称为日本大叶藻或日本鳗草(李政等,2020),隶属于大叶藻科(Zosteraceae),大叶藻属(Zostera),为多年生草本高等植物,广泛分布于亚洲的中国、日本、韩国、越南等地,在中国广泛分布于亚热带和温带沿海地区。矮大叶藻与大叶藻(Zostera marina) 相比,藻体较小但环境适应能力更强,能够生活于水温和光照波动较大、环境扰动较强的浅海潮间带(张晓梅等,2013)。20 世纪初,矮大叶藻随着牡蛎运输,从日本扩散至从加拿大太平洋西岸的不列颠哥伦比亚到美国俄勒冈之间的广大海域(Posey et al,1988)。矮大叶藻作为生态入侵种不断扩展延伸,将泥滩改造成为海草床,提高了当地生态系统的初级生产力和生物多样性,成为极少数具有正面影响的入侵种之一(Posey et al,1988)。
海草植被覆盖与环境因子都是影响潮间带大型底栖动物群落的重要生态因子(张景平等,2011;Bell et al,1986)。海草不仅可以通过影响海流强度和方向来改变底质粒径和有机物组成,同时其植株也为微型藻类、微小型底栖动物、附着动物和底上动物提供了复杂多样的微生境(杨宗岱 等,1984)和庇护场所(Xu et al,2016),提高海草床定居动物的丰富度、密度(Posey et al,1988;Snelgrove et al,1994)、生物多样性和生产力(Xu et al,2016)。
近年来黄河口附近海域大型底栖动物的研究备受关注,但关于海草床大型底栖动物的研究则未有报道。与相邻山东省东营市黄河口碱蓬植被覆盖的潮间带海域的大型底栖动物群落研究相比,碱蓬覆盖海域的大型底栖动物仅27 种(未发表数据),物种数量远少于海草床海域获得的41 种;若仅从本研究分析,海草床区域的33 种底栖动物的物种数量也远高于10 m 和30 m 处的21 种和20 种。与碱蓬植被海域和无植被覆盖海域的比较说明,海草床能够为海洋大型底栖动物提供更好的栖息地环境和复杂的生境,即使退化海草床作为较差的庇护所,庇护所内被捕食死亡率也会明显低于无覆盖海床同类的死亡率(Bell et al,1986),因此可供养更多的物种数量和更高的群落丰度。
3.2 大型底栖动物群落、环境因子的相关性分析
本研究海域地处黄河口,此处由于长期的黄河入海泥沙冲积,潮滩平缓宽阔,在退潮时滩面仍可保持较大湿度,矮大叶藻在此海域呈现镶嵌状或连续状分布。沉积物粒径是影响海草床分布及生物学特征的重要因素(Schan et al,2003)。大型底栖动物作为海草床生态系统的重要组成部分之一,其分布与沉积物类型和海草群落特征密切相关(Blanchet et al,2004)。矮大叶藻适宜生长的沉积物组成以砂和粉砂为主,这样的基质或许能够为矮大叶藻发达的地下茎提供更多的氧气(张晓梅等,2013)。
一般情况下,海底沉积物粒径和水流强度呈正相关关系,沉积物颗粒直径越小,表明该区域的水动力越小(Andral et al,1999)。尽管本研究中的矮大叶藻植株较小且退化呈斑块状,但该海草床依旧可以通过影响水动力和生境状况,增加细颗粒物在海草床及其周围的沉降,改变周围底质的粒径组成,使较粗粒径的占比在距离海草床30 m 处(95.85%)明显高于海草床10 m 范围内的底质粒径(92.96%)(0.01 底质特征是控制底栖动物丰度和分布的重要因子(Han et al,2016),且异质性高的底质环境中的生物多样性要高于异质性低的底质环境(Schan et al,2003)。本研究中,粗砂为底栖动物营造了较高异质性的底质环境,物种数目与底质粒径中的粗砂呈显著相关关系。在软底质生境中,悬浮滤食性动物通常会在大型底栖动物群落中占据优势地位(Russell,1987)。在本研究中,大型底栖动物功能群中的浮游生物食者(亦即悬浮滤食动物)丰度占总丰度的49.40%,仅光滑河篮蛤和红明樱蛤两种浮游生物食者的生物量就贡献了群落总生物量的55.43%,这主要得益于粗砂带来的较多间隙为浮游生物提供了更多生存空间(Pires et al,1999)。 海草床生态系统中的海草本身及附着动植物,为大型底栖动物提供了丰富多样的食物来源(Lewis,1984;高亚平等,2010)、复杂的栖息微生境、良好的育幼场所(Costanza et al,1997)以及躲避捕食的避难所(Lee et al,2001),对大型底栖动物群落组成、多样性和空间分布格局具有重要的影响(Bostr觟m et al,2006)。另一方面,大型底栖动物作为初级生产者和鱼类等更高营养级消费者之间的桥梁,是底栖生态系统中物质循环、能量流动的关键角色;除此之外,它们可通过摄食和掘穴等扰动活动直接或间接地影响海草的生长底质环境,通过改变底质的粒径组成和有机质在不同深度中的含量和转运,协助维持海草床生态系统功能的完整性(张景平等,2011)。 在矮大叶藻海草床区域,多毛类、十足目动物等优势种的丰度等数量特征与海草床的覆盖显著相关,海草床附近的动物丰度要远高于远离海草床海域(Posey et al,1988;Tanner,2010)。在本研究中,群落总丰度、优势种丝异须虫和色斑角吻沙蚕的丰度,均与海草床呈现显著的正相关,亦即海草床区域的丰度显著高于无植被覆盖的邻近海域;与此同时,其余5 种优势种,即光滑河篮蛤、红明樱蛤、拟背尾水虱、奇异稚齿虫和三叶针尾涟虫,它们的丰度与斑块状海草床的覆盖并未显示出显著的相关性(P>0.05)。 本研究中,对处于同一高程的海草床区域和非覆盖区域进行采样,排除了高程对大型底栖动物群落分带和分布的影响,以精确地揭示海草床覆盖底质对大型底栖动物群落的影响。通过群落的聚类分析结果显示,海草床区域的两个大型底栖动物群落并未聚合在一起,大型底栖动物在斑块状海草床内部的分布并不均匀,揭示了退化海草床在生态服务功能方面的不稳定和不均一性。 山东省广饶潮间带海草床共有大型底栖动物41 种,包括甲壳动物(16 种)、多毛类动物(15种)和软体动物(10 种),海草床覆盖海域的物种数明显高于远离海草床区域的物种数;海草床覆盖海域的群落丰度亦明显高于非海草覆盖海域。 大型底栖动物群落丰度的主要贡献者为丝异须虫和光滑河篮蛤。群落优势种共有7 种,多毛类动物有3 种,甲壳动物和软体动物各2 种。沉积物粒径中粗砂与物种数量呈密切相关关系。粗砂间存在较多空隙更利于矮大叶藻根部的呼吸以及光滑河篮蛤和红明樱蛤等浮游生物食者的生存。 大型底栖生物功能群主要以浮游生物食者为主,各功能群丰度占比分别为浮游生物食者(49.40%)>碎屑食者(37.10%)>肉食者(7.96%)>杂食者(5.54%),仅肉食者的空间分布存在显著性差异,其他功能群空间分布差异并不显著。 综上所述,大型底栖动物群落的各分类类群和功能群在海草床距离梯度上并未表现出显著的线性变化,但海草床对底栖动物群落丰度、优势种等的影响非常显著。因此,斑块状海草床的生态服务功能仍然存在,但已经相当有限,因此对功能退化的斑块状海草床的保护势在必行,以期恢复其原有的生态服务功能。3.3 海草床与底栖动物群落的相互分析
4 结论
