二化螟P糖蛋白基因的克隆分析及对杀虫剂的诱导响应
2021-10-20孟祥坤吴赵露杨雪梅官道杰王建军
孟祥坤,吴赵露,杨雪梅,官道杰,王建军
二化螟P糖蛋白基因的克隆分析及对杀虫剂的诱导响应
孟祥坤,吴赵露,杨雪梅,官道杰,王建军
扬州大学园艺与植物保护学院,江苏扬州 225009
【】克隆二化螟()P糖蛋白基因()并对其分子特征和表达模式进行分析,明确对常用防治杀虫剂氯虫苯甲酰胺和阿维菌素的诱导响应并对其潜在的转录调控机制进行探索。使用基因克隆技术扩增全长基因序列,利用生物信息学技术对编码蛋白的分子特征和5′端转录调控区中的转录因子结合位点进行分析。使用荧光定量PCR方法对在二化螟不同龄期和不同组织中的表达模式及在杀虫剂氯虫苯甲酰胺和阿维菌素不同剂量处理下的诱导响应进行测定分析。cDNA序列全长4 584 bp,由23个外显子构成,编码1 259个氨基酸,含有两个跨膜区和两个核苷酸结合区,具有ABC转运蛋白家族典型的结构特征,如对底物转运具有重要功能的Walker A、Walker B及D、H、P、Q-Loop等特征序列。主要在二化螟幼虫期表达,在3—4龄幼虫中具有最高的表达量,在蛹期和成虫期的表达量较低。组织表达分析表明,主要高表达于二化螟的前肠和中肠组织,在后肠、脂肪体、马氏管等其他组织中的表达量较低。相比于对照,使用LC30和LC70剂量氯虫苯甲酰胺分别处理二化螟3龄幼虫12 h和24 h后,的表达量未发生显著变化。但在处理36 h后,LC30处理组试虫中显著上调表达,而LC70处理组试虫中则显著下调表达。使用低剂量0.05 mg·L-1的阿维菌素处理二化螟试虫12 h后,相比于对照,显著下调表达,在处理24 h和36 h后的表达水平没有发生显著变化,使用0.15 mg·L-1的阿维菌素处理二化螟试虫24 h和36 h后被显著诱导上调表达。对的5′端转录调控区的序列分析发现,在转录调控区中预测到多个转录因子结合位点,其中包括5个潜在的CncC结合位点。二化螟在重要解毒代谢组织中肠中高表达,并且能够被杀虫剂氯虫苯甲酰胺和阿维菌素诱导表达,表明可能参与对氯虫苯甲酰胺和阿维菌素的解毒代谢。5′端转录调控区中含有多个转录因子CncC结合位点,可能对的转录表达具有重要调控作用。推测在氯虫苯甲酰胺或阿维菌素胁迫下,可能受到转录因子CncC的转录调控并参与对氯虫苯甲酰胺或阿维菌素的解毒代谢。
二化螟;P糖蛋白;分子特征;杀虫剂诱导;转录调控
0 引言
【研究意义】二化螟()是我国水稻上危害最为严重的常发性害虫之一,每年导致我国水稻大量减产[1]。目前二化螟的防治主要使用化学农药,但是大量使用农药引发的害虫抗药性等问题日益严重[2-4]。监测发现,二化螟已对氯虫苯甲酰胺、阿维菌素等几种常用防治杀虫剂产生了不同水平的抗药性[5-7]。因此,通过对二化螟抗药性研究,探索潜在的抗性机制,不仅为新型高效杀虫剂开发提供理论依据,同时也为二化螟田间防治的药剂选择提供指导,对于农业害虫防治具有重要意义。【前人研究进展】对外源化合物的解毒代谢是昆虫适应环境及对杀虫剂产生抗药性的主要策略之一。昆虫对植物有毒物质和杀虫剂等外源化合物的解毒代谢主要分3个反应阶段:I相反应中,细胞色素P450(cytochrome P450,P450)和酯酶(esterase,EST)分别氧化和水解外源化合物,生成水溶性更高的代谢产物;II相反应中,谷胱甘肽-转移酶(glutathione-transferase,GST)和尿苷二磷酸糖基转移酶(UDP-glycosyltransferase,UGT)通过催化结合反应进一步增加代谢产物的水溶性;III相反应中,腺苷三磷酸结合盒转运蛋白(ABC转运蛋白,ATP-binding cassette transporter)将水溶性的代谢产物从细胞内转运到细胞外[8-9]。ABC转运蛋白是多细胞动物中最大的转运蛋白家族,含有8个(ABCA—ABCH)亚家族成员,在生物中发挥泵的作用将有毒物质排出细胞。昆虫ABC转运蛋白中的ABCB、ABCC和ABCG家族成员广泛参与杀虫剂的转运代谢,与昆虫的杀虫剂抗药性密切相关[10]。对二化螟的杀虫剂抗药性研究中,已发现包括I、II相反应中的多个P450、EST和UGT基因可能参与二化螟对杀虫剂的解毒代谢,但关于二化螟对杀虫剂解毒代谢的III相反应研究却鲜有报道[6,11-13]。【本研究切入点】ABCB家族中的又称P糖蛋白基因(P-glycoprotein gene,)。广泛参与对药物的转运,是昆虫中被研究最多的ABC基因,在多种昆虫中参与对杀虫剂的转运和抗药性[10]。作为III相解毒代谢反应中最重要的ABC基因之一,是否参与了二化螟对杀虫剂的抗药性还有待于进一步研究。【拟解决的关键问题】通过对二化螟克隆分析,明确其在二化螟中的表达模式以及在杀虫剂诱导下的表达变化,通过对5′端转录调控区的序列分析,探索潜在的转录调控机制,为进一步深入了解二化螟抗药性机制打下基础。
1 材料与方法
试验于2020年在扬州大学园艺与植物保护学院农药分子靶标与环境毒理实验室完成。
1.1 供试昆虫与试剂
敏感品系二化螟试虫在温度为(28±1)℃,相对湿度为(70±5)%,光周期为16 h﹕8 h(光照﹕黑暗)的室内培养箱中,使用人工饲料进行饲养[14]。试虫饲养过程中不接触任何农药。
cDNA末端扩增试剂盒SMARTer RACE 5′/3′ Kit、DNA聚合酶 LA Taq、cDNA第一链合成试剂盒PrimeScriptTM1st cDNA Synthesis Kit、RNA提取试剂盒Takara MiniBEST Universal RNA Extraction Kit、凝胶回收试剂盒Takara MiniBEST Agarose Gel DNA Extraction Kit、荧光定量反转录试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser和荧光定量试剂TB Green Premix Ex TaqTM等购自宝生物工程(大连)有限公司(Takara);昆虫基因组提取试剂盒Insect DNA Kit(Omega)购于扬州祥瑞生物科技有限公司;克隆载体pEASY-T1和感受态细胞Trans5购于北京全式金生物技术有限公司。95%氯虫苯甲酰胺和93.7%阿维菌素原药由扬州大学园艺与植物保护学院冯建国老师提供。
1.2 CsPgp的克隆和序列分析
在二化螟转录组中鉴定到若干注释为的序列片段[15],进一步通过与同源基因的序列比对和拼接,使用Primer premier 5软件设计特异引物(表1)用于部分编码区序列的扩增。50 μL PCR体系中含有5 μL 10× LA PCR Buffer(Mg2+Plus),0.5 μL LA Taq(5 U·μL-1),上下游引物(10 μmol·L-1)各2 μL,8 μL dNTP Mixture(各2.5 nmol·L-1),cDNA模板2 μL,灭菌蒸馏水32.5 μL。PCR反应条件为95℃预变性2 min;95℃变性10 s,60—55℃(每循环降低0.5℃)退火15 s,72℃延伸3 min,循环10次;95℃变性10 s,55℃退火15 s,72℃延伸3 min,循环25次;72℃延伸10 min。使用琼脂糖凝胶电泳检测PCR产物,对大小正确的DNA条带进行胶回收、连接到克隆载体并转化感受态细胞。使用菌落PCR对挑选的单克隆进行鉴定并测序。根据测序结果和cDNA末端扩增试剂盒使用说明,设计特异引物(表1),用于5′端和3′端的序列扩增。使用CLUSTAL W和DNAMAN 7.0软件对克隆的基因序列进行比对分析,使用在线软件ExPASy Compute pI/Mw(http://ca.expasy. org/tools/pi_tool.html)对翻译的蛋白质分子质量和等电点进行预测。MEGA 7软件用于昆虫的进化树分析。NCBI序列分析软件用于CsPgp保守结构特征分析。
使用NCBI中二化螟基因组(Bioproject:PRJNA506136)的序列信息,分析基因组结构,并设计特异引物,以二化螟幼虫DNA为模板,扩增基因编码区上游5′侧翼区DNA序列。使用在线软件JASPAR(http://jaspar.genereg.net)、ALLGEN(http://alggen.lsi.upc.es)、http://www.fruitfly.org/seq_ tools/promoter.html和http://www.softberry.com对克隆的5′侧翼区DNA序列中的转录因子结合位点、启动子序列和转录起始位点进行预测分析。
1.3 CsPgp时空表达分析
根据克隆获得的序列信息,设计特异性引物,使用荧光定量PCR对在二化螟不同发育时期和不同组织中的表达量进行分析。分别收集二化螟各龄期幼虫、不同日龄蛹和成虫,每5—8头试虫为一个样品。在二化螟3龄幼虫中分别解剖脑、神经索、前肠、中肠、后肠、血淋巴、脂肪体、表皮和马氏管组织,在羽化24 h的雌成虫中解剖卵巢组织。每个样品分别收集3个生物学重复。根据试剂盒说明书,使用Takara MiniBEST Universal RNA Extraction Kit试剂盒提取样品RNA,并利用琼脂糖凝胶检测RNA的完整性。使用荧光定量反转录试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser合成用于基因表达量分析的cDNA模板,不同模板统一稀释至100 ng·μL-1。荧光定量PCR反应体系为10 µL 2×TB Green Premix Ex Taq,10 μmol·L-1的上下游引物各1 µL,cDNA模板2 µL,6 μL灭菌超纯水。PCR反应条件为95℃预变性2 min;95℃变性30 s,60℃退火30 s,40个循环。以二化螟中稳定表达的为内参基因[16-17]。使用2-∆∆Ct方法计算靶标基因在每个样品中的相对表达量,使用one-way ANOVA 进行差异显著性分析与多重比较,试验数据以平均数±标准误表示。
1.4 CsPgp对杀虫剂的诱导反应
根据前期试验结果,分别配制含有LC30(0.092 mg·L-1)和LC70(0.47 mg·L-1)氯虫苯甲酰胺[15]及含有0.05、0.15 mg·L-1阿维菌素的人工饲料。使用含有杀虫剂的饲料及正常饲料处理二化螟3龄幼虫,每个处理接入60头幼虫,重复3次。在处理12、24和36 h后收集样品试虫,每5头二化螟幼虫为一个样品,用于的表达量分析。使用one-way ANOVA Tukey’s test进行差异显著性分析,试验数据以平均数±标准误表示(*<0.05,**<0.01,***<0.001)。
2 结果
2.1 CsPgp的克隆与分析
克隆获得4 584 bp全长cDNA序列,其中开放阅读框3 780 bp,5′非翻译区125 bp,3′非翻译区679 bp(图1-A)。编码1 259个氨基酸,预测的蛋白质分子质量为137.8 kD,等电点为6.58。编码蛋白具有ABC全转运体家族典型的结构特征,包括两个跨膜区和两个核苷酸结合区,以及核苷酸结合区中对底物转运具有重要功能的保守特征序列如Walker A、Walker B及D、H、P、Q-Loop等(图1-A)。基因组结构分析显示,基因组全长38 596 bp,由23个外显子构成,外显子平均长度175 bp(图1-B)。对不同物种Pgp的进化树分析显示,CsPgp与其他鳞翅目昆虫Pgp共聚一支,具有较高的同源性(图2)。
2.2 CsPgp的表达谱分析
在幼虫期的表达量较高,其中在幼虫中期(3—4龄)具有最高的表达量(图3)。在蛹期和成虫期的表达量较低,且在不同时期的表达量差异不显著。组织表达分析显示,在测定的10个组织中均有表达,其中在前肠和中肠组织中显著高表达,在脂肪体中的表达量最低(图4)。
2.3 CsPgp对杀虫剂的诱导响应
与对照组试虫相比,使用0.05 mg·L-1剂量阿维菌素处理幼虫12 h后,的表达量显著降低,而在处理24 h和36 h后的表达量没有发生显著变化。当使用0.15 mg·L-1剂量阿维菌素处理幼虫12 h后,的表达量没有发生显著变化,但在处理24 h和36 h后的表达量分别增加至对照的2.2和2.8倍。分别使用LC30(0.092 mg·L-1)和LC70(0.47 mg·L-1)剂量的氯虫苯甲酰胺处理幼虫12 h和24 h后,幼虫中的表达量均未发生显著变化。但在处理36 h后,相比于对照,LC30处理组幼虫中的表达量显著增加至5.5倍,而LC70处理组幼虫中的表达量则显著降低(图5)。
2.4 CsPgp 5′端转录调控区序列分析
基于二化螟基因组序列信息,克隆了上游5′侧翼区约2 000 bp DNA序列,并对序列中的转录因子结合位点、启动子序列及转录起始位点等转录调控元件进行预测分析。分析结果显示,在5′上游调控区中预测到一个潜在的启动子序列(Score=0.91)及多个转录因子结合位点(Score>0.8)(图6)。预测的转录因子结合位点中包括5个Cnc::maf-S,3个EcR::usp及br、Deaf1、dl、tin、Dfd等。

A:CsPgp核酸序列及其编码的氨基酸序列,深灰色背景标注的为CsPgp跨膜区序列;浅灰色背景标注的为CsPgp核苷酸结合区,其中重要的结构特征使用单下划线标出,斜体氨基酸为ATP结合位点Nucleotide and deduced amino acid sequences of CsPgp. The transmembrane domains are marked by dark grey background, and the nucleotide-binding domains are marked by light grey background. The important structural characteristics in nucleotide-binding domains are underlined, and the italic amino acids indicated as the binding sites of ATP。B:CsPgp基因组结构Genomics structure of CsPgp。Intron:内含子;Exon外显子;UTR:非翻译区Untranslated region
3 讨论
二化螟是水稻作物上最主要的害虫之一,广泛分布于亚洲地区。由于长期使用化学农药防治,目前我国一些地区二化螟田间种群已对氯虫苯甲酰胺、阿维菌素等常用防治药剂产生了不同水平的抗药性[5-7]。根据2017—2019年有害生物抗药性监测结果,浙江、江西和湖南等部分地区二化螟种群对氯虫苯甲酰胺处于高水平抗性,对阿维菌素处于中等至高水平抗性;江苏、安徽、湖北和四川等部分地区二化螟种群对氯虫苯甲酰胺处于敏感至中等水平抗性,对阿维菌素处于敏感水平;不同地区二化螟种群对氯虫苯甲酰胺的抗性倍数逐年增加[2-4]。对二化螟抗药性机制的研究发现,在对氯虫苯甲酰胺产生抗性的二化螟中,P450和EST的酶活性显著增加,使用相应的增效剂分别抑制P450、EST和UGT的酶活性后二化螟对氯虫苯甲酰胺的敏感性显著增加[6,11,13]。在二化螟抗性种群中4个P450基因(、、和)和2个UGT基因(和)显著过表达,使用RNA干扰分别沉默这6个基因均能显著增加二化螟对氯虫苯甲酰胺的敏感性[12-13]。此外,使用亚致死剂量氯虫苯甲酰胺处理二化螟后,多个P450基因被诱导表达,可能参与对氯虫苯甲酰胺的解毒代谢[15]。对阿维菌素抗性机制研究发现,氨基酸突变导致的靶标敏感性降低和解毒代谢基因过表达造成的酶活性增强均能够引起昆虫和螨对阿维菌素产生抗药性,但关于二化螟对阿维菌素的抗药性机制却仍不清楚[18-22]。
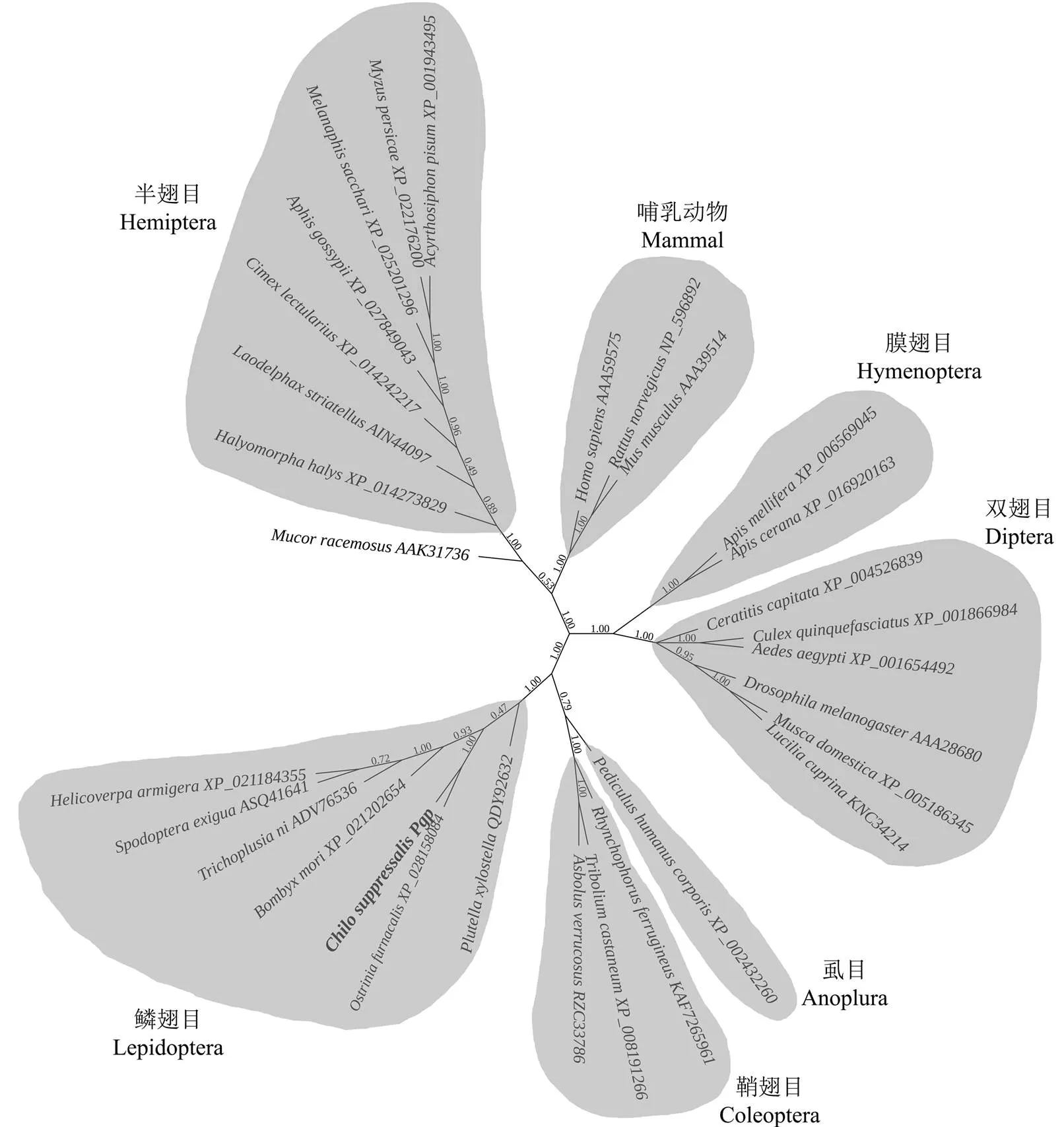
图2 不同物种Pgp的进化树分析
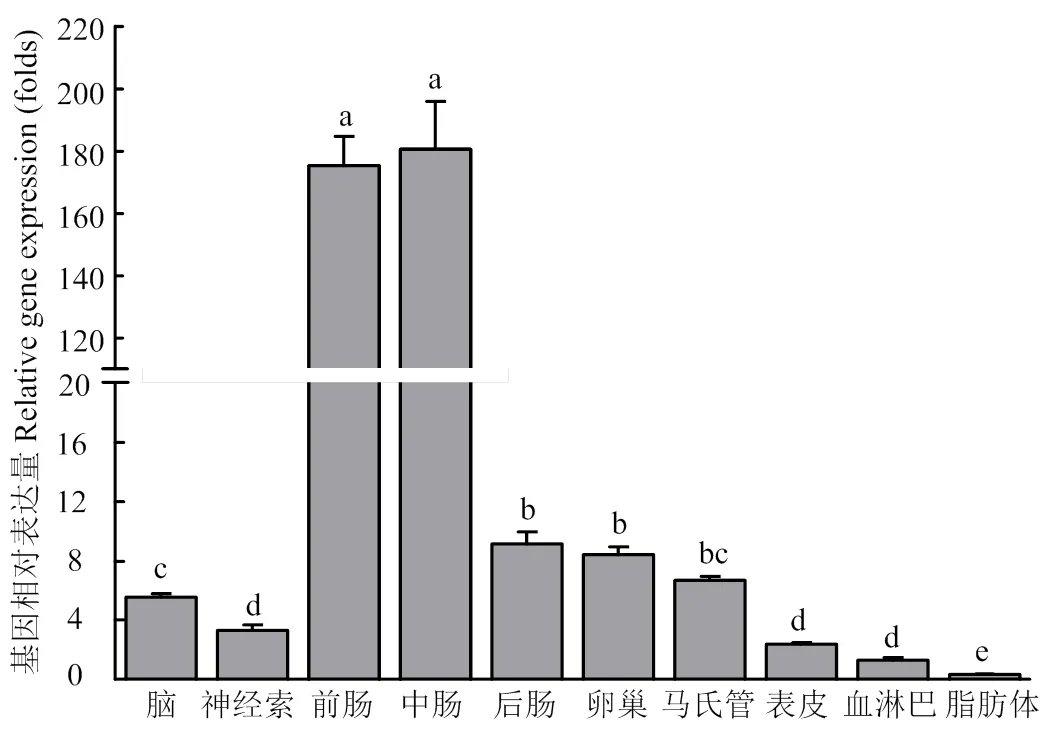
柱上标有不同字母表示CsPgp在不同组织表达量差异显著(P<0.05) Histograms with different letters indicate significant difference of CsPgp expression in different tissues (P<0.05)。脑:brain;神经索:nerve cord;前肠:foregut;中肠:midgut;后肠:hindgut;卵巢:ovary;马氏管:Malpighian tubule;表皮:cuticula;血淋巴:hemolymph;脂肪体:fat body
研究表明,ABC转运蛋白中的参与调节昆虫、螨虫对杀虫剂的转运代谢[10,23-24]。为进一步探索是否参与III相解毒代谢反应对氯虫苯甲酰胺和阿维菌素的转运代谢,本研究克隆了全长序列并进行分析。开放阅读框由23个外显子构成,编码1 259个氨基酸,包含两个跨膜区和两个核苷酸结合区,属于典型的全转运蛋白,在核苷酸结合区中含有对底物转运具有重要功能的所有保守特征序列[10]。基因表达分析显示,主要在二化螟幼虫期表达,在3—4龄幼虫中具有最高的表达量,在蛹期和成虫期表达量较低,与棉铃虫()的表达模式相似[24]。但在小菜蛾()中在不同幼虫期和蛹期的表达量差异不显著[25]。组织表达分析中,主要在前肠和中肠表达,小菜蛾同样在中肠中显著高表达,而棉铃虫则在头部和脂肪体中具有较高的表达量[24-25]。昆虫食性和习性的不同可能是导致表达模式差异的原因,进而造成在不同昆虫中可能具有不同功能。
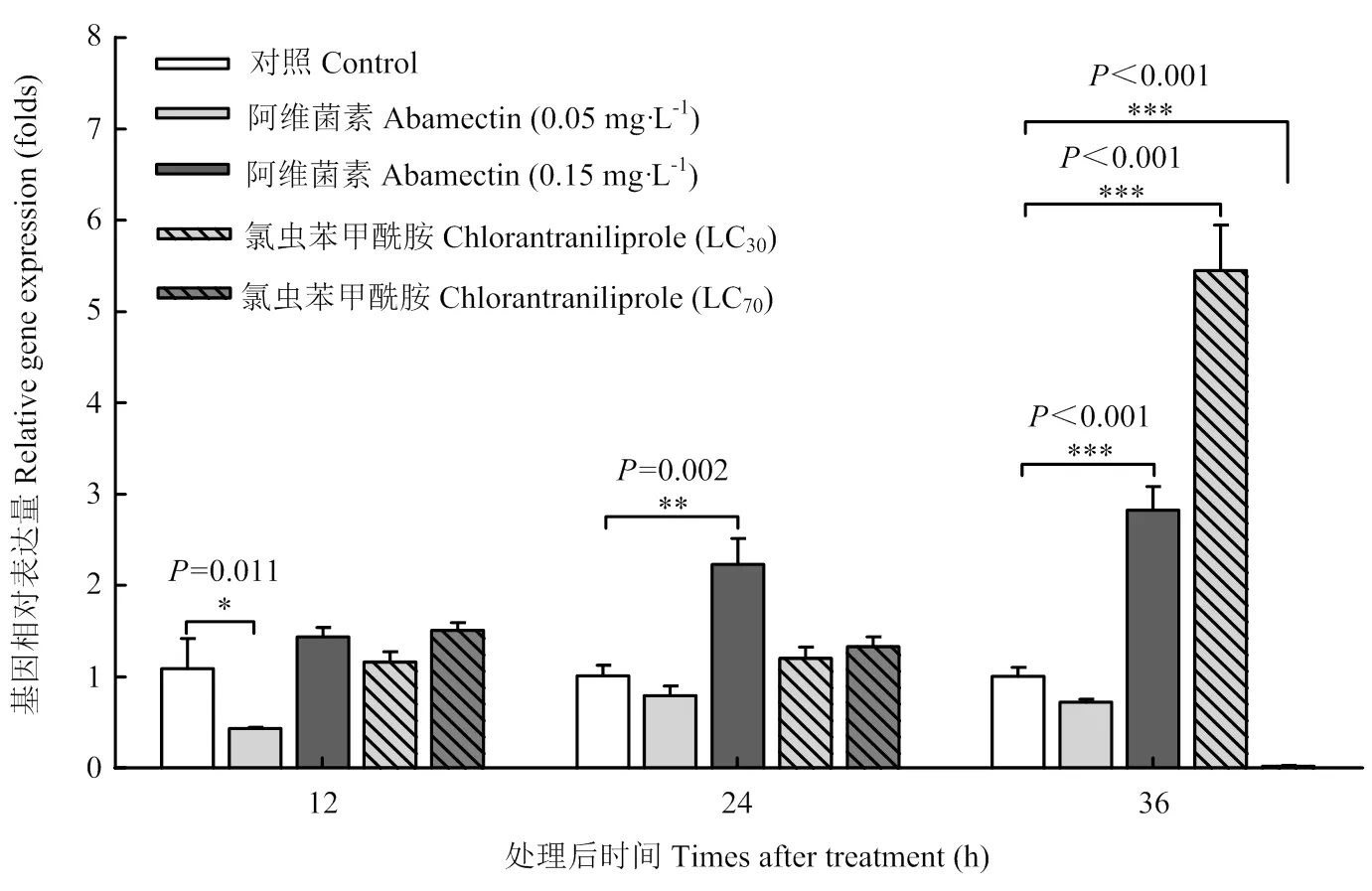
*表示存在显著差异* indicates significant difference (* P<0.05; ** P<0.01; *** P<0.001)

单下划线标注的为预测的转录因子结合位点(Score>0.8),双下划线标注的为预测的启动子序列(Score=0.91),其中粗体字母为预测的转录起始位点。以CsPgp编码区起始密码子的第一个碱基位置标为“+1”,其上游碱基标为“-” The predicted transcription factors are marked by single underline, and the predicted promoter sequence is marked by double underline. The transcription start site is indicated by bold letter. The first base of initiation code of CsPgp is indicated as “+1”, and its upstream sequences are indicated as “-”
与昆虫的抗药性机制相似,当受到外界压力胁迫如杀虫剂暴露时,昆虫同样可通过提高解毒代谢基因的表达来增加对压力胁迫的抵抗能力[26]。与解毒代谢基因的持续过表达相比,诱导表达被认为存在着一种代谢代价,只有当解毒代谢需要时才会激活解毒代谢基因的表达[27]。昆虫中除了P450、GST等解毒代谢基因可被诱导表达外,ABC转运蛋白基因同样可被杀虫剂诱导表达[9-10,28-31]。前期研究发现,二化螟中多个ABC转运蛋白基因可被亚致死剂量的氯虫苯甲酰胺诱导表达,其中()在处理后24 h轻微上调表达[9]。本研究发现,使用LC30和LC70剂量氯虫苯甲酰胺分别处理二化螟幼虫12 h和24 h后,的表达水平未发生显著变化。但在处理36 h后,LC30处理组试虫显著上调表达,而在较高的药剂浓度LC70处理组中,可能由于试虫的生理进程遭受严重破坏而导致显著下调表达。说明不同药剂浓度处理下试虫生理进程的不同可能造成的可诱导性存在差异,CsPgp可能参与二化螟对氯虫苯甲酰胺的转运代谢。使用0.05 mg·L-1的阿维菌素处理二化螟12 h后显著下调表达,在处理24 h和36 h后的表达量没有发生显著变化,可能是使用的药剂浓度太低,不能够对产生有效的诱导作用。但使用0.15 mg·L-1的阿维菌素处理二化螟24 h和36 h后被显著诱导上调表达。在小菜蛾、棉铃虫和朱砂叶螨()中同样发现可被阿维菌素诱导表达[22,24,32]。此外,使用RNA干扰减少的表达显著增加了阿维菌素对棉铃虫致死率[24],CRISPR/Cas9介导的基因敲除显著增加了甜菜夜蛾()对阿维菌素的敏感性[33]。这些结果说明包括二化螟在内的多种昆虫的参与了对阿维菌素的解毒代谢。
基因的转录表达受到多种作用因子的影响,如反式作用因子、顺式作用元件等。反式作用因子也称转录因子,是一类在细胞核内与特异靶标基因结合激活或抑制靶标基因转录的DNA结合蛋白[34]。目前在昆虫中已发现多种转录因子参与调控昆虫解毒代谢基因的转录表达,如Cap ‘n’ collar isoform C(CncC)[8,35-36]。使用溴氰菊酯处理黑腹果蝇()胚胎细胞可诱导的表达,并进一步引起解毒代谢和抗氧化基因的上调表达从而增加细胞对杀虫剂的解毒和耐受能力[37]。使用氯虫苯甲酰胺或辛硫磷处理家蚕()均能引起CncC在mRNA和蛋白质表达水平发生变化,并引起下游解毒代谢基因的上调表达进而增加P450和GST的酶活性[38-39]。斜纹夜蛾()中,可被茚虫威诱导表达并通过参与调控多个解毒代谢基因的过表达介导斜纹夜蛾对茚虫威的抗药性[40]。本研究中,通过对5′端转录调控区序列分析,发现多个转录因子结合位点,其中包括5个Cnc::maf-S(即CncC)结合位点。此外,使用氯虫苯甲酰胺处理显著上调了在二化螟中的表达水平(未发表数据)。这些结果说明可能受到转录因子CncC的转录调控,从而介导二化螟对杀虫剂的解毒代谢。
4 结论
二化螟由23个外显子组成,编码的蛋白质具有ABC转运蛋白家族典型的结构特征。主要在幼虫的前肠和重要解毒代谢组织中肠中高表达,并且能够被杀虫剂氯虫苯甲酰胺和阿维菌素诱导上调表达,说明可能参与对氯虫苯甲酰胺和阿维菌素的解毒代谢。5′端转录调控区中含有多个转录因子CncC结合位点,可能对的转录表达具有重要作用。推测在杀虫剂胁迫下二化螟可能通过CncC调控的转录表达,从而增加其对杀虫剂的解毒代谢。
[1] 刘万才, 刘振东, 黄冲, 陆明红, 刘杰, 杨清坡. 近10年农作物主要病虫害发生危害情况的统计和分析. 植物保护, 2016, 42(5): 1-9.
LIU W C, LIU Z D, HUANG C, LU M H, LIU J, YANG Q P. Statistics and analysis of crop yield losses caused by main diseases and insect pests in recent 10 years. Plant Protection, 2016, 42(5): 1-9. (in Chinese)
[2] 全国农业技术推广服务中心. 2017年全国农业有害生物抗药性监测结果及科学用药建议. 中国植保导刊, 2018, 38(4): 52-56.
National Agricultural Technology Extension Service Center. Monitoring results of pesticide resistance of agricultural pests and suggestions for scientific pesticide use in China in 2017. China Plant Protection, 2018, 38(4): 52-56. (in Chinese)
[3] 全国农业技术推广服务中心. 2018年全国农业有害生物抗药性监测结果及科学用药建议. 中国植保导刊, 2019, 39(3): 63-67, 72.
National Agricultural Technology Extension Service Center. Monitoring results of pesticide resistance of agricultural pests and suggestions for scientific pesticide use in China in 2018. China Plant Protection, 2019, 39(3): 63-67, 72. (in Chinese)
[4] 全国农业技术推广服务中心. 2019年全国农业有害生物抗药性监测结果及科学用药建议. 中国植保导刊, 2020, 40(3): 64-69.
National Agricultural Technology Extension Service Center. Monitoring results of pesticide resistance of agricultural pests and suggestions for scientific pesticide use in China in 2019. China Plant Protection, 2020, 40(3): 64-69. (in Chinese)
[5] WEI Y B, YAN R, ZHOU Q L, QIAO L Y, ZHU G N, CHEN M L. Monitoring and mechanisms of chlorantraniliprole resistance in(Lepidoptera: Crambidae) in China. Journal of Economic Entomology, 2019, 112(3): 1348-1353.
[6] LU Y H, WANG G R, ZHONG L Q, ZHANG F C, BAI Q, ZHENG X S, LU Z X. Resistance monitoring of(Walker) (Lepidoptera: Crambidae) to chlorantraniliprole in eight field populations from east and central China. Crop Protection, 2017, 100: 196-202.
[7] MAO K K, LI W H, LIAO X, LIU C Y, QIN Y, REN Z J, QIN X Y, WAN H, SHENG F, LI J H. Dynamics of insecticide resistance in different geographical populations of(Lepidoptera: Crambidae) in China 2016-2018. Journal of Economic Entomology, 2019, 112(4): 1866-1874.
[8] KALSI M, PALLI S R. Cap n collar transcription factor regulates multiple genes coding for proteins involved in insecticide detoxification in the red flour beetle,. Insect Biochemistry and Molecular Biology, 2017, 90: 43-52.
[9] MENG X K, YANG X M, WU Z L, SHEN Q W, MIAO L J, ZHENG Y, QIAN K, WANG J J. Identification and transcriptional response of ATP-binding cassette transporters to chlorantraniliprole in the rice striped stem borer,. Pest Management Science, 2020, 76(11): 3626-3635.
[10] DERMAUW W, VAN LEEUWEN T. The ABC gene family in arthropods: Comparative genomics and role in insecticide transport and resistance.Insect Biochemistry and Molecular Biology, 2014, 45: 89-110.
[11] SUN Y, XU L, CHEN Q, QIN W J, HUANG S J, JIANG Y, QIN H G. Chlorantraniliprole resistance and its biochemical and new molecular target mechanisms in laboratory and field strains of(Walker). Pest Management Science, 2018, 74(6): 1416-1423.
[12] XU L, ZHAO J, SUN Y, XU D J, XU G C, XU X L, ZHANG Y L,HUANG S J, HAN Z J, GU Z Y. Constitutive overexpression of cytochrome P450 monooxygenase genes contributes to chlorantraniliprole resistance in(Walker). Pest Management Science, 2019, 75(3): 718-725.
[13] ZHAO J, XU L, SUN Y, SONG P P, HAN Z J. UDP-glycosyltransferase genes in the striped rice stem borer,(Walker), and their contribution to chlorantraniliprole resistance. International Journal of Molecular Sciences, 2019, 20(5): 1064.
[14] 李波, 韩兰芝, 彭于发. 二化螟人工饲养技术. 应用昆虫学报, 2015, 52(2): 498-503.
LI B, HAN L Z, PENG Y F. Development of a standardized artificial diet and rearing technique for the striped stem borer,Walker (Lepidoptera: Crambidae). Chinese Journal of Applied Entomology, 2015, 52(2): 498-503. (in Chinese)
[15] MENG X K, DONG F, QIAN K, MIAO L J, YANG X M,GE H C,WU Z L, WANG J J. Transcriptome analysis reveals global gene expression changes ofin response to sublethal dose of chlorantraniliprole. Chemosphere, 2019, 234: 648-657.
[16] 徐红星, 王国荣, 鲁艳辉, 杨亚军, 郑许松, 田俊策, 吕仲贤. 二化螟实时荧光定量PCR内参基因筛选和表达稳定性评价. 中国水稻科学, 2019, 33(1): 75-84.
XU H X, WANG G R, LU Y H, YANG Y J, ZHENG X S, TIAN J C, LÜ Z X. Screening reference genes and evaluating of their expression stability for qRT-PCR normalization in(Lepidoptera: Pyralididae). Chinese Journal of Rice Science, 2019, 33(1): 75-84.(in Chinese)
[17] XU J, LU M X, CUI Y D, DU Y Z. Selection and evaluation of reference genes for expression analysis using qRT-PCR in(Lepidoptera: Pyralidae). Journal of Economic Entomology, 2017, 110(2): 683-691.
[18] DERMAUW W, ILIAS A, RIGA M, Tsagkarakou A, GRBIC M, TIRRY L, VAN LEEUWEN T, VONTAS J. The cys-loop ligand-gated ion channel gene family of: Implications for acaricide toxicology and a novel mutation associated with abamectin resistance. Insect Biochemistry and Molecular Biology, 2012, 42(7): 455-465.
[19] LIAO C Y, XIA W K, FENG Y C, LI G, LIU H, DOU W, WANG J J. Characterization and functional analysis of a novel glutathione-transferase gene potentially associated with the abamectin resistance in(McGregor). Pesticide Biochemistry and Physiology, 2016, 132: 72-80.
[20] RIGA M, TSAKIRELI D, ILIAS A, MOROU E, MYRIDAKIS A, STEPHANOU E G, NAUEN R, DERMAUW W, VAN LEEUWEN T, PAINE M, VONTAS J. Abamectin is metabolized by CYP392A16, a cytochrome P450 associated with high levels of acaricide resistance in. Insect Biochemistry and Molecular Biology, 2014, 46: 43-53.
[21] WANG X L, PUINEAN A M, O’REILLY A O, WILLIAMSON M S, SMELT C L C, MILLAR N S, WU Y D. Mutations on M3 helix ofglutamate-gated chloride channel confer unequal resistance to abamectin by two different mechanisms. Insect Biochemistry and Molecular Biology, 2017, 86: 50-57.
[22] YIN Q, QIAN L, SONG P P, JIAN T Y, HAN Z J. Molecular mechanisms conferring asymmetrical cross-resistance between tebufenozide and abamectin in. Journal of Asia-Pacific Entomology, 2019, 22(1): 189-193.
[23] LUO L, SUN Y J, WU Y J. Abamectin resistance inis related to increased expression of P-glycoprotein via the dEGFR and dAkt pathways. Insect Biochemistry and Molecular Biology, 2013, 43(8): 627-634.
[24] XIANG M, ZHANG L, LU Y, TANG Q L, LIANG P, SHI X Y, SONG D L, GAO X W. A P-glycoprotein gene serves as a component of the protective mechanisms against 2-tridecanone and abamectin in. Gene, 2017, 627: 63-71.
[25] TIAN L X, YANG J Q, HOU W J, XU B Y, XIE W, WANG S L, ZHANG Y J, ZHOU X G, WU Q J. Molecular cloning and characterization of a P-glycoprotein from the diamondback moth,(Lepidoptera: Plutellidae). International Journal of Molecular Sciences, 2013, 14(11): 22891-22905.
[26] ENDERS L S, RAULT L C, HENG-MOSS T M, SIEGFRIED B D, MILLER N J. Transcriptional responses of soybean aphids to sublethal insecticide exposure. Insect Biochemistry and Molecular Biology, 2020, 118: 103285.
[27] TERRIERE L C. Induction of detoxication enzymes in insects.Annual Review of Entomology, 1984, 29: 71-88.
[28] HE C, LIANG J J, LIU S N, WANG S L,WU Q J, XIE W, ZHANG Y J. Changes in the expression of four ABC transporter genes in response to imidacloprid inQ (Hemiptera: Aleyrodidae). Pesticide Biochemistry and Physiology, 2019, 153: 136-143.
[29] JIN M H, LIAO C Y, CHAKRABARTY S, ZHENG W G, WU K M, XIAO Y T. Transcriptional response of ATP-binding cassette (ABC) transporters to insecticides in the cotton bollworm,. Pesticide Biochemistry and Physiology, 2019, 154: 46-59.
[30] MERZENDORFER H. ABC transporters and their role in protecting insects from pesticides and their metabolites//COHEN E. Target Receptors in the Control of Insect Pests: Part II. 2014, 46: 1-72.
[31] SUN H, PU J, CHEN F, WANG J D, HAN Z J. Multiple ATP-binding cassette transporters are involved in insecticide resistance in the small brown planthopper,. Insect Molecular Biology, 2017, 26(3): 343-355.
[32] XU Z F, SHI L, PENG J F, SHEN G M, WEI P, WU Q, HE L. Analysis of the relationship between P-glycoprotein and abamectin resistance in(Boisduval). Pesticide Biochemistry and Physiology, 2016, 129:75-82.
[33] ZUO Y Y, HUANG J L, WANG J, FENG Y, HAN T T, WU Y D, YANG Y H. Knockout of a P-glycoprotein gene increases susceptibility to abamectin and emamectin benzoate in. Insect Molecular Biology, 2018, 27(1): 36-45.
[34] BAKSHI M, OELMüLLER R. WRKY transcription factors: Jack of many trades in plants. Plant Signaling and Behavior, 2014, 9(2): e27700.
[35] HU B, HU S Z, HUANG H, WEI Q, REN M M, HUANG S F, TIAN X R, SU J Y. Insecticides induce the co-expression of glutathione-transferases through ROS/CncC pathway in. Pesticide Biochemistry and Physiology, 2019, 155: 58-71.
[36] WILDING C S. Regulating resistance: CncC:Maf, antioxidant response elements and the overexpression of detoxification genes in insecticide resistance. Current Opinion in Insect Science, 2018, 27: 89-96.
[37] CHEN L, ZHANG T T, GE M Y, LIU Y H, XING Y P, LIU L, LI F L,CHENG L G. The Nrf2-Keap1 pathway: A secret weapon against pesticide persecution inKc cells.Pesticide Biochemistry and Physiology, 2020, 164: 47-57.
[38] CHENG X Y, HU J H, LI J X, CHEN J, WANG H, MAO T T, XUE B, Li B. The silk gland damage and the transcriptional response to detoxifying enzymes-related genes ofunder phoxim exposure. Chemosphere, 2018, 209: 964-971.
[39] MAO T T, LI F C, FANG Y L, WANG H, CHEN J, LI M X, LU Z T, QU J W, LI J X, HU J H, CHENG X Y, NI M, LI B. Effects of chlorantraniliprole exposure on detoxification enzyme activities and detoxification-related gene expression in the fat body of the silkworm,. Ecotoxicology and Environmental Safety, 2019, 176: 58-63.
[40] SHI L, SHI Y, LIU M F, ZHANG Y, LIAO X L. Transcription factor CncC potentially regulates the expression of multiple detoxification genes that mediate indoxacarb resistance in. Insect Science, 2021, https://doi.org/10.1111/1744-7917.12860.
Cloning and Analysis of P-glycoprotein gene and its transcriptional response to insecticide in
MENG Xiangkun, WU Zhaolu, YANG Xuemei, GUAN Daojie, WANG Jianjun
College of Horticulture and Plant Protection, Yangzhou University, Yangzhou 225009, Jiangsu
【】Thewas cloned from, and the molecular characteristics and expression profiles ofwere analyzed. Transcriptional responses as well as the potential transcriptional regulation mechanism ofto two common used insecticides (chlorantraniliprole and abamectin) were also studied.【】The full length ofwas cloned fromusing the gene cloning technology. The molecular characteristics and the transcription factor binding sites in 5′ transcriptional regulatory region ofwere analyzed employing the bioinformatics technologies. Expression profiles ofin different stages and tissues of, and the transcriptional responses ofto different doses of chlorantraniliprole and abamectin treatment were determined using the real-time quantitative PCR.【】The full length ofcdna is 4 584 bp and consists of 23 exons. The encoding protein has 1 259 amino acids containing two transmembrane regions and two nucleotide binding domains and the typical structural features of ABC transporter family such as the Walker A, Walker B and D, H, P, Q-Loop which have important function in substrate transfer.was mainly expressed in larval stage of, especially in the 3rd and 4th instar larvae, whileshowed low expression levels in the pupal and adult stages. Analysis of the tissue expressions showed thatwas predominately expressed in the foregut and midgut, and had very low expression levels in other tissues including hindgut, fat body and malpighian tubule. No significant change ofexpression was found in the 3rd instar larvae ofafter treated with LC30and LC70of chlorantraniliprole for 12 and 24 h, respectively, when compared with the control groups. However, the expressions ofwere significantly up-regulated in larvae after treated with LC30of chlorantraniliprole for 36 h, while the expressions ofwere significantly down-regulated in larvae after treated with LC70of chlorantraniliprole for 36 h. In the 0.05 mg·L-1of abamectin treatment,was remarkably down-regulated at 12 h post-treatment, while the expressions ofwere not significantly changed at 24 and 36 h post-treatment, respectively. However,was significantly induced in larvae after treated with 0.15 mg·L-1of abamectin for 24 and 36 h, respectively. Sequence analysis of the 5′ transcriptional regulatory region ofshowed that multiple transcription factor binding sites were predicted in the 5′ transcriptional regulatory region of, including five potential CncC binding sites.【】was highly expressed in the midgut ofand could be induced by chlorantraniliprole and abamectin, which indicated thatmight involve in the detoxification metabolism of chlorantraniliprole and abamectin in. Multiple CncC binding sites were found in the 5′ transcriptional regulatory region ofwhich might have important regulatory effects on the expression of. It was speculated thatmight be regulated by transcription factor CncC and participated in the detoxification metabolism of chlorantraniliprole or abamectin whenwas exposed to chlorantraniliprole or abamectin.
; P-glycoprotein; molecular characteristic; insecticide induction; transcriptional regulation

10.3864/j.issn.0578-1752.2021.19.008
2021-02-06;
2021-02-27
国家自然科学基金青年基金(31701807)、江苏省自然科学基金青年基金(BK20170491)
孟祥坤,E-mail:mxk@yzu.edu.cn。通信作者王建军,E-mail:wangjj@yzu.edu.cn
(责任编辑 岳梅)
