Neotype designation and redescription of Tropidophorus guangxiensis Wen, 1992 (Squamata:Sauria: Scincidae), with description of a new subspecies from central South China
2021-10-18Ke-JiGuo,FuShu,Nan-FeiWu等
The genusTropidophorusconsists of small semi-aquatic lizards that dwell in lowland forest steams (Barbour, 1921;Bauer & Jackman, 2008). Here, we designate the neotype and re-describeT. guangxiensisWen, 1992 based on newly collected topotypic specimens. We also describe a new subspecies from Xuefeng Mountain, Hongjiang County, Hunan Province, central South China. Based on two mitochondrial genes (12S rRNA and 16S rRNA), the phylogenetic position ofT. guangxiensisis allocated for the first time. Additionally, our data strongly support that the new subspecies is phylogenetically closely related toT. g. guangxiensis. We also present a morphological identification key for known species and subspecies ofTropidophorusin China.
The genusTropidophorus, first described by Duméril &Bibron (1839) (type species:Tropidophorus cocincinensisDuméril & Bibron, 1839), currently comprises 29 species that range from north-eastern India, Bangladesh, mainland South-East Asia, and southern China, to Peninsular Malaysia,Borneo, as well as the southern Philippines and Sulawesi(Truong et al., 2010a; Pui et al., 2017; Uetz et al., 2021; Zhao et al., 1999). In China, fourTropidophorusspecies have been recorded, namely,T. berdmorei(Blyth, 1853),T. guangxiensisWen, 1992,T. hainanusSmith, 1923, andT. sinicusBoettger,1886 (Figure 1A) (Cai et al., 2015; Ji & Wen, 2002; Qi, 2019;Shen et al., 2014; Tian & Jiang, 1986; Wen, 1992; Zhang,2011; Zhao & Adler, 1993; Zhao et al., 1999).Tropidophorus guangxiensisWen, 1992 was described based on three specimens (one juvenile and two adult males) collected on 15 June 1985 from Daming Mountain, Guangxi Zhuang Autonomous Region, southern China. No female specimens have been recorded since its original description, and little is known about the morphological variation of the species.Moreover, the type specimens are presumed to be lost (Mr.Qi-Ping Hu, personal communication, 2021). During field work in the Guangxi Zhuang Autonomous Region of China, we collected two new specimens ofT. guangxiensisfrom Daming Mountain, Wuming County. To clarify the taxonomic confusion ofT. guangxiensis, we designated a neotype and paraneotype and provided an updated description based on the newly designated type series and original description.
A total of six specimens were collected during field work in Xuefeng Mountain, Hongjiang County, Hunan Province,China, between 2007 and 2016. Two specimens were collected from Daming Mountain, Wuming County, Guangxi Zhuang Autonomous Region, China, in 2019. The collected specimens were fixed in formalin after euthanasia, then flushed with water and preserved in 70% ethanol. Tissue samples taken from specimens after euthanasia but before fixation were preserved in 95% ethanol. All specimens were deposited at the College of Life Science, Hunan Normal University (HNU), Changsha, China. Sex of specimens was determined through minor dissection. The new sequences produced in this study were deposited in GenBank under accession Nos. MW411357-MW411360 and MW413384-MW413387 (Supplementary Table S1). Morphological terminology and measurements were taken with a digital caliper to the nearest 0.1 mm (see Supplementary Materials and Methods).
Whole genomic DNA was extracted from tissue samples,and two mitochondrial genes, i.e., 12S rRNA and 16S rRNA,were amplified and sequenced. Details on DNA extraction,primers, and polymerase chain reaction (PCR) protocols are provided in the Supplementary Materials and Methods.Phylogenetic relationships of the genusTropidophoruswere reconstructed based on the two mtDNA fragments. In addition,sequences of 22 congeners of the genus were downloaded from GenBank (Supplementary Table S1). Phylogenetic trees were reconstructed using maximum-likelihood (ML) and Bayesian inference (BI) (Supplementary Materials and Methods). Pairwise sequence divergences were calculated using uncorrectedP-distances implemented in MEGA v6.0.6(Tamura et al., 2013) (Supplementary Table S2).
The ML and BI phylogenies were congruent, with relatively high nodal support for most terminal nodes (Figure 1B). The genusTropidophoruswas divided into two monophyletic groups (BPP/BSP=1.00/100). The four Chinese species (T.berdmorei, T. guangxiensis, T. hainanus, andT. sinicus) were assigned to the same clade, withT. guangxiensismost closely related toT. sinicus.The specimens from Hongjiang County formed a sister lineage toT. guangxiensis(BPP/BSP=1.00/100). The interspecific genetic distances (based on 16S rRNA) forTropidophorusranged from 0.3% to 10.7%. The genetic distance betweenT. guangxiensisand the putative new subspeciesT. guangxiensiswas 2.7%, equivalent to that betweenT. latiscutatusandT. matsuii(2.1%) and betweenT.thaiandT. robinsoni(2.4%).
The newly collected specimens ofT. guangxiensismatched the diagnosis of the original description. In addition, based on our data, the Hongjiang population ofTropidophoruswas most similar toT. guangxiensis, but showed a certain level of genetic and morphological differentiation from the topotypic population ofT. guangxiensis(see below). Due to the limited comparative materials of trueT. guangxiensis, we tentatively consider the Hongjiang population as a new subspecies ofT.guangxiensis.
Taxonomic account
Tropidophorus guangxiensis guangxiensis Wen, 1992(Supplementary Figures S1-S3; Supplementary Tables S3-S6)
Chresonymy:Tropidophorus guangxiensis: Wen YT, 1992,Asiatic Herpetological Research, Berkeley, 4: 18-22. Type locality: Damingshan, Wuming Xian (county), Guangxi Zhuang Autonomous Region, China, 1 240 m a.s.l.
Justification of neotype designation:Wen (1992) describedT. guangxiensisbased on three specimens (one juvenile and two adult males), which were deposited at the Department of Biology, Guangxi Medical College (GMC). According to information provided by Mr. Qi-Ping Hu (Director of the Department of Cell Biology and Genetics, former Department of Biology, Guangxi Medical University, Personal Communication, 2021), all three type specimens are presumed lost. Furthermore, as our new data suggest there may be additional cryptic diversity withinT. guangxiensis,designating a neotype and re-describing the species definition have become essential. The neotype and paraneotype were from the same locality as the original type series, and they match the diagnosis of the species based on the type series by Wen (1992), including GMC 85-029, GMC 85-030, and GMC 85-032. Based on available morphological data, we believe the neotype and paraneotype represent the same taxa as the original type series, i.e.,T. guangxiensis.Herein, we redescribe the speciesT. g. guangxiensisbased on fresh specimens from the type locality and designate a neotype for this species. Diagnosis was updated based on the fresh specimens from the type locality and the original description.
Neotype:HNU GKJ-2019007 (adult female), collected on 21 August 2019 by Ke-Ji Guo, Qi-Lin Pan, and Yan-Wu Lu in Daming Mountain National Nature Reserve (N23°30'1'' and E108°26'19''), Wuming County, Guangxi Zhuang Autonomous Region, China, at an altitude of 1 244 m a.s.l.
Paraneotype:HNU GKJ-2019009 (sub-adult male), collected on 22 August 2019 by Ke-Ji Guo, Qi-Lin Pan, and Jian-Chun Li in Daming Mountain National Nature Reserve (N23°48'3''and E108°45'32''), Wuming County, Guangxi Zhuang Autonomous Region, China, at an altitude of 1 280 m a.s.l.
Diagnosis:Upper head scales strongly striated; supranasal absent; frontonasals 2, joined, longer than wide; prefrontals 2,joined or separated by small scale; interparietal single and small, without small transparent spot; parietals separated by interparietal, posterolateral border surrounded by 5-6 scales on each side; nuchal scales absent; supraciliaries 8,supraciliary row complete along length of lateral edge of supraoculars; loreals 2; presuboculars 1, upper anterior and lower posterior margin slightly convex, and terminal pointed;supralabials 8; infralabials 6-7; shallow groove on loreal-labial border, from posterior corner of nasal across subocular obliquely downward to end of sixth supralabial; midbody scales in 28-29 rows; paravertebral scales 45-50, not widened; ventral scales 50-53; midbody ventral scales 27-29;ventral scales of neck keeled; scale rows at tenth subcaudal 11-13; lamellae under fourth toe 17-18; scales of limbs keeled above and below; supralabials and infralabials black,each scale with white spot center; chin and throat grayish white with black marble; ventral of neck gray, each scale white in middle forming longitudinal white stripes; venter yellowish white; underside of tail white, subcaudals darkened on both sides, forming longitudinal white stripes.
Description of neotype(detailed morphometric measurements in Supplementary Tables S3, S4): Body size relatively small (SVL of 62.5 mm; TaL of 68.8 mm).
Snout acute, rounded anteriorly; projecting beyond lower jaws; nostril laterally oriented, oval, closer to snout-tip than to orbit; head distinct from neck and body, longer than wide;head shape slightly flattened; upper head scales strongly striated; rostral wider than high, visible from above;supranasal absent; frontonasals 2, joined, longer than wide;prefrontals 2, separated by small scale; frontal narrowing posteriorly, in contact with frontonasal, prefrontals, first and second supraoculars, and frontoparietals; frontoparietals joined, bordered by frontal, second, third, and fourth supraoculars, parietals, and interparietal; interparietal single,without small transparent spot; parietals separated by interparietal, posterolateral border surrounded by five scales on each side; nuchal scales absent; nasal single, nostrils in posterior center, located closer to anterior loreal; loreals 2,weakly straited; posterior loreal higher and broader than anterior loreal; anterior loreal undivided, posterior loreal in contact with anterior loreal, prefrontal, first supraciliary,preocular, upper presubocular, second and third supralabial;preoculars 1; presuboculars 1, upper anterior and lower posterior margin slightly convex, and terminal pointed, in contact with third and fourth supralabials; supraciliaries 8, first two obviously larger, supraciliary row complete along length of lateral edge of supraoculars; supraoculars 4, second widest;postocular single; temporals strongly keeled; primary temporals 3; secondary temporals 4; eyes relatively small;pupil circle, discernable in preserved specimen; upper palpebrals 15; lower palpebrals 15-16; lower eyelid moveable,with six enlarged opaque scales, separated from supralabials by two rows of small scales; supralabials 8, fifth largest and sixth highest; upper parts slightly straited and lower parts smooth, except for first smooth overall; infralabials 7/6, first longest; shallow groove on loreal-labial border, from posterior corner of nasal across subocular obliquely downward to end of sixth supralabial; external ear opening ovoid, without lobules;tympanum superficial, tympanum diameter much smaller than eye length; mental wider than long, rounded anteriorly, in contact with first infralabial on each side and postmental;postmental undivided, in contact with mental, first infralabial on both sides and anterior pair of chinshields; chinshields in three pairs, first pair in contact anteriorly, second and third pairs separated by gular scale; first and second pairs of chinshields in contact with infralabials, third pair in contact anteriorly with infralabials and separated posteriorly from infralabials by small scales; tongue short; undivided anteriorly,tip obtuse, not pointed; teeth relatively small and somewhat pointed.
Body slender, midbody scales in 29 rows; dorsal scales in eight rows across back, slightly larger than lateral scales,imbricated; sharply keeled with acute-to-normal points, except several rows following interparietal smooth or keeled with at least two lines: paravertebral scales 50, not widened; lateral scales slightly smaller than dorsal and ventral scales, keeled with single line, obliquely upward or near horizontal; ventral scales smooth, except for ventral scales of neck keeled;ventral scales slightly larger than dorsal and lateral scales, in 50 transversal rows; midbody ventral scales in 29 transversal rows, counted from posterior margin of fore limb to last scale before precloacals; precloacals 4, inner precloacals overlapping outer ones, central two enlarged.
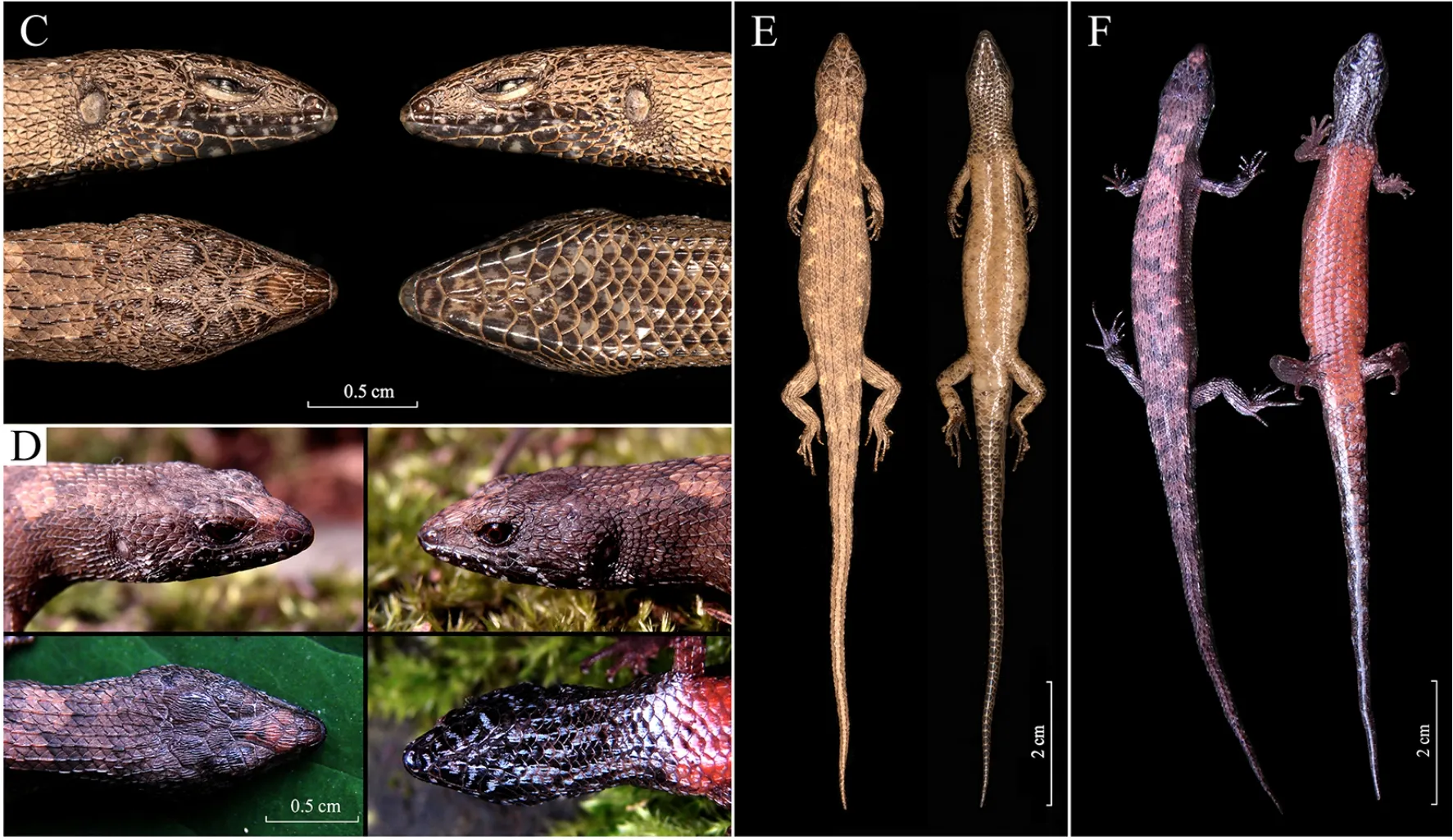
Limbs slender and short. Forelimbs shorter than hindlimbs;fingers 5, fourth finger includes claw beyond seventh supralabial and wrist reaches posterior margin of tympanum when adpressed forelimbs are carried forward; relative length of fingers I Tail laterally compressed, relatively long, longer than SVL;tail tip acute; tail base wider than rest of tail; tail gradually tapering to point; dorsal scales of tail keeled, forming continuous ridge, scale rows 13, at tenth subcaudal;subcaudals 59; medial subcaudals divided from first to fifth,remaining ones approximately 1.5 times wider than neighboring scales; visceral fat bodies absent in abdomen. Coloration in life(Supplementary Figures S1-S3): Pupil black, iris brownish red. Head rusty brown dorsally and laterally, with yellowish, blackish cloudy spots; dorsum brown with irregular brownish red transverse bands and spots from neck to middle of body, body sides dark gray with irregular light brownish yellow dots from behind tympanum to groin;dorsally dark gray with brownish red dots; supralabials and infralabials black, each scale with white center spot; chin and throat grayish white with black marble; ventral neck gray, each scale white in middle, forming longitudinal white stripes;ventral body orange; ventral tail white, subcaudals darkened on both sides, forming longitudinal white stripes; limbs orange ventrally. Coloration in preservative(Supplementary Figures S1-S3):Coloration and ornamentation remain mostly the same after preservation, except orange coloration of ventral body fades to brown. Variations:Measurements, pholidosis, and color patterns of the paraneotype are summarized in Supplementary Tables S3, S4. Distribution and ecology:Tropidophorus guangxiensis guangxiensisis currently known only from the Daming Mountain National Nature Reserve (type locality) and Mulun Nature Reserve in Guangxi Zhuang Autonomous Region,southern China (Figure 1A; Supplementary Table S6).Individuals were found beneath withered branches and fallen leaves in roadside ditches under evergreen broadleaf forest at 1 240-1 280 m a.s.l. (Supplementary Figure S4). These skinks are known to be active in late August, but their diet and reproductive biology remain unknown. This subspecies is sympatric withGonyosoma frenatum(Gray, 1853),Pareas boulengeri(Angel, 1920), andTakydromussp. Comparisons:Based on the designated neotype and paraneotype, we comparedTropidophorus guangxiensis guangxiensiswith congeners from China (morphological data of congeners were taken from literature: Fei et al., 2010; Guo et al., 2010; Rao et al., 2011; Wen, 1992; Yang & Inger, 1986;Yang & Rao, 2008; Zhang et al., 2012; Zhao & Adler, 1993;Zhao et al., 1999) (Supplementary Table S5):Tropidophorus guangxiensis guangxiensiscan be distinguished fromT.berdmorei,T. hainanus, andT. sinicusby supralabials 7-8(vs. 6 in all latter species); the subspecies can be distinguished fromT. berdmoreiby paravertebral scales 49-50 (vs. 64), upper head and dorsal body scales striated or keeled (vs. smooth), midbody scale rows 28-29 (vs. 32-40),and lamellae under fourth toe 17-18 (vs. 22).Tropidophorus guangxiensis guangxiensiscan be distinguished fromT.hainanusby parietals separated posteriorly (vs. in contact posteriorly), loreals 2 (vs. 4), supraciliaries 8 (vs. 5),supraciliary row complete along length of lateral edge of supraoculars (vs. supraciliary row interrupted by fourth supraocular), midbody scale rows 28-29 (vs. 30-34), and lamellae under fourth toe 17-18 (vs. 13-16).Tropidophorus guangxiensis guangxiensiscan be distinguished fromT.sinicusby ventral scales smooth (vs. keeled), supraciliaries 8(vs. 6), infralabials 6-7 (vs. 5), and lamellae under fourth toe 17-18 (vs. 13-14). Tropidophorus guangxiensis hongjiangensis ssp. nov.Guo & Deng(Figure 1C-F; Supplementary Tables S3-S6). Chresonymy:Tropidophorus guangxiensis: Guo KJ, Mi XQ and Deng XJ, 2010, Chinese Journal of Zoology,45(1):159-162; Mi XQ, Xiong JW, Shu Y, Huang Z, Guo KJ,Huang Y, et al. 2012. Hunan Forestry Science & Technology,39(4), 50-53. Holotype:HNU GKJ-2016009, adult female, collected on 5 June 2016 by Ke-Ji Guo in Xuefeng Mountain National Forest Park (N27°18'57'' and E110°23'55''), Hongjiang County,Hunan Province, China, at an altitude of 1 125 m a.s.l. Paratype:HNU GKJ-2016022 (sub-adult female) and HNU GKJ-2016023 (juvenile), collected on 15 June 2016 by Wei-Ping Xia in the Xuefeng Mountain National Forest Park(N27°18'28'' and E110°23'55''), Hongjiang County, Hunan Province, China, at an altitude of 1 056 m a.s.l. HNU GKJ-200704001 (adult female), HNU GKJ-200704002 (adult male),HNU GKJ-200704003 (adult female), and HNU GKJ-200704004 (adult female) were collected on 13 April 2007 by Ke-Ji Guo in the Xuefeng Mountain National Forest Park(N27°18'56'' and E110°23'37''), Hongjiang County, Hunan Province, China, at an altitude of 1 126 m a.s.l. Etymology:This new subspecies is named after its type locality in “Hongjiang County.” Figure 1 Distribution, phylogenetic position, and type specimens of Tropidophorus guangxiensis hongjiangensis ssp. nov. Guo & Deng Diagnosis:Loreals 2; infralabials 6; supralabials 8;paravertebral scales 45-47; parietals separated; supraciliaries 6-8; supraciliary row complete; upper anterior and lower posterior margin of presuboculars slightly concave, terminal blunt and rounded; scales on upper head and dorsal body striated or keeled; ventral scales smooth; midbody scale rows 30-33; lamellae under fourth toe 16-19; throat and ventral of neck black; ventral limbs brown, darker than ventral body;middle and posterior ventral tail black. Description of holotype:Adult female, body size relatively small (SVL, 58.45 mm; TaL, 69.05 mm). Snout acute (IN:IO ratio 0.38), rounded anteriorly; projecting beyond lower jaws; nostril laterally oriented; oval, situated closer to snout-tip than to orbit; nostril-snout distance 1.2 mm,eye-nostril distance 2.4 mm, eye-snout distance 3.8 mm (NS:E-S ratio, 0.32; E-N:E-S ratio, 0.64); head distinct from neck and body, longer than wide; HL 9.8 mm, HW 7.8 mm (HL:HW ratio 1.25); head shape slightly flattened, HD 5.8 mm (HL:HD ratio 1.68), HNL 20.3. Upper head scales strongly striated;rostral wider than high, visible from above; supranasal absent;frontonasals 2, joined (suture length 1.2 mm), longer than wide, in contact with rostral, nasal, anterior loreal, and prefrontal on each side; prefrontals 2, separated by single small scale, in contact with frontonasal, anterior and posterior loreal, first supraciliary, first supraocular and frontal; frontal narrowing posteriorly, 1.51 times longer than distance to snout, and 1.61 times longer than interparietal length (2.0 mm), in contact with frontonasal, prefrontals, first and second supraoculars, and frontoparietals; frontoparietals joined(suture length 0.95 mm), bordered by frontal, third and fourth supraoculars, parietals and interparietal; interparietal single,small, without small transparent spot; parietals separated by interparietal and two small scales, posterolateral border surrounded by six scales on each side; without nuchal scales;nasal single, nostrils in posterior center, located closer to anterior loreal; loreals 2, straited; posterior loreal higher and broader than anterior one; anterior loreal undivided, posterior loreal in contact with anterior loreal, prefrontal, first supraciliary, preocular, upper presubocular, second and third supralabials; preocular 1; presubocular 1, upper anterior and lower posterior margin slightly concave, terminal blunt and rounded, in contact with third and fourth supralabials;supraciliaries 7, first two obviously larger, supraciliary row complete along length of lateral edge of supraoculars;supraoculars 4, second widest, followed by two small postsupraoculars; postocular single; pretemporals 2;postsuboculars 2, lower one in contact with sixth and seventh supralabials; temporals strongly keeled; primary temporals 3,lower one in contact with seventh supralabial; secondary temporals 5, upper one very large, in contact with parietal;eyes relatively small; pupil circle, discernable in preserved specimen; upper palpebrals 14; lower palpebrals 15; lower eyelid moveable, with six enlarged opaque scales, separated from supralabials by two rows of small scales; supralabials 8,fifth largest and sixth highest, upper part of supralabials slightly straited and lower part smooth from second to eighth supralabial; infralabials 7, first longest; shallow groove on loreal-labial border, from posterior corner of nasal across subocular obliquely downward to end of sixth supralabial;external ear opening ovoid, without lobules; tympanum superficial, tympanum diameter much smaller than eye length;mental wider than long, rounded anteriorly, in contact with first infralabial on each side and postmental; postmental undivided,in contact with mental, first infralabial on both sides, and anterior pair of chinshields; chinshields in three pairs, first pair in contact anteriorly, second pair separated by gular scale,third pair separated by three scales; first and second pairs of chinshields in contact with infralabials, third pair in contact with infralabials anteriorly and separated from infralabials by small scales posteriorly; tongue short; undivided anteriorly, tip obtuse, not pointed; teeth relatively small and somewhat pointed. Body slender, depressed, BW 12.5 mm (BW:SVL ratio 0.19); midbody scales in 33 rows; dorsal scales in eight rows across back, slightly larger than lateral scales, imbricated;scales of first four rows following interparietal striated, sharply keeled with acute to normal points posteriorly by fifth row;paravertebral scales 47, not widened; lateral scales slightly smaller than dorsal and ventral scales, keeled with single line,obliquely upward except for lower two rows near horizontal;ventral scales smooth, except for ventral scales of neck keeled; ventral scales slightly larger than dorsal and lateral scales, in 50 transversal rows, midbody ventral scales 31 transversal rows, counted from posterior margin of fore limb to last scale before precloacals; precloacals 4, inner precloacals overlapping outer ones, central two enlarged, left one overlapped by right one. Limbs slender, short. Forelimbs 24.55% of SVL, shorter than hindlimbs (FLL 67.12% of HLL); with five fingers, fourth finger includes claw beyond sixth supralabial and wrist reaching middle of tympanum when adpressed forelimbs are carried forward; relative length of fingers I Tail laterally compressed; base enlarged, gradually narrowing to acute point from end to tip; longer than tip of snout to vent length, latter almost 1.18 (SVL 84.65% of TAL);subcaudals enlarged, smooth, larger than dorsal scales, latter strongly keeled, form six continuous ridges on dorsal view;subcaudals 59, 14 scale rows at tenth subcaudal; medial subcaudals divided from first to sixth, remaining ones about two times wider than neighboring scales; visceral fat bodies absent in abdomen. Coloration in life(Figure 1D, F): Pupil black, iris brownish red. Dorsal head, back, and tail base dark brown with indistinct light brownish red transverse bands, faded from anterior to posterior: one between anterior corners of eye, two on neck, six between limb insertions, five on tail base; dorsum with two rows of brownish red dots from neck to point above vent; body sides dark gray with irregular brownish red dots from behind tympanum to groin; supralabials and infralabials black, each scale with white center spot; limbs dorsally dark gray with brownish red dots; chin and throat black with brownish red spots; ventral of neck dark brown, each scale white in middle, forming longitudinal white stripes; ventral from posterior margin of fore limb to underside of tail base orange;underside of tail tip black; color of tail underside darkened from base to tip; brownish red ventrally with indistinct irregular dark transverse stripes. Coloration in preservative(Figure 1C, E): Coloration and ornamentation remain mostly the same after preservation,except the orange coloration of ventral body fades into brown. Variation:Measurements, pholidosis, and color patterns of the paratypes are summarized in Supplementary Tables S3,S4. Distribution:Tropidophorus guangxiensis hongjiangensisssp. nov.is currently known only from the Xuefeng Mountain National Forest Park in Hongjiang County, Hunan Province,China (Figure 1A; Supplementary Table S6). Natural history:Tropidophorus guangxiensis hongjiangensisssp. nov. is a semi-aquatic, leaf-litter specialist inhabiting the forest floor of subtropical evergreen broadleaf and mixed forests. Specimens were collected in the late afternoon and evening (1600-2200 h) from under withered branches and fallen leaves in roadside ditches (Supplementary Figure S5),or near rocky forest streams. The diet of the new subspecies remains unknown. The gravid female holotype carried four eggs on 5 June 2016. This subspecies is sympatric withPlestiodon elegans(Boulenger, 1887), Oreocryptophis porphyraceus(Cantor, 1839),Pareas stanleyi(Boulenger,1914), andTakydromus intermediusStejneger, 1924. Comparisons: Morphological comparisons were based on fiveT. g. guangxiensisand sixTropidophorus guangxiensis hongjiangensisssp. nov. specimens (Supplementary Tables S3-S5). The new subspecies can be distinguished from the nominal subspecies by midbody scale rows 30-33 (vs.28-29), midbody ventral scales 30-33 (vs. 27-29), upper anterior and lower posterior margin of presuboculars slightly concave and terminal blunt and rounded (vs. upper anterior and lower posterior margin slightly convex and terminal pointed) (Supplementary Figure S3E, F, e, f). The new subspecies can be further distinguished fromT. g.guangxiensisby throat and ventral neck black (Figure 1D, F;Supplementary Figure S3B, D, a, b, c, d) (vs. gray(Supplementary Figures S1, S2, S3A, C, a, b, c, d)), ventral limbs brown and darker than ventral surface (Figure 1F;Supplementary Figure S3G, g, h) (vs. ventral limbs orange and consistent with ventral surface (n=2; Supplementary Figures S2, S3H, i, j)), middle and posterior ventral of tail black (n=6) (vs. middle and posterior ventral of tail white,(n=2); Figure 1E; Supplementary Figures S2, S3K). Comments:The previous records ofT. guangxiensis(Guo et al., 2010; Mi et al., 2012) from the Xuefeng Mountain National Forest Park in Hongjiang County, Hunan Province, i.e., type locality ofTropidophorus guangxiensis hongjiangensisspp.nov., are therefore allocated toTropidophorus guangxiensis hongjiangensisspp. nov. It should be noted that there is a relatively high level of genetic divergence betweenT. g. guangxiensisandTropidophorus guangxiensis hongjiangensisspp. nov., which is larger than the current interspecific genetic distances among recognized congeners (Supplementary Table S2).Future studies should collect additional specimens to gain a better understanding of the morphological and genetic variations with the genusTropidophorus. Water skinks in the genusTropidophorusare a group of small, secretive, and elusive forest-floor dwelling lizards, and little is known about their diversity and distribution. The discovery of the new subspecies suggests an underestimation of the diversity of this group and a wider distribution of the genus than previously thought. In the past, distribution of the genusTropidophoruswas thought to be from the southern subtropical region of China, with the northern boundary of its distribution located in the Nanling Mountains at a latitude of N24° (Zhao et al., 1999). However, our recent discoveries push this limit further north to approximately N26° in Guizhou Province (Yang & Rao, 2008; Zhang et al., 2012). Our discovery of a new subspecies further extends the northern distribution of the genus to N27° latitude. Updated key to Tropidophorus from China(Guo et al., 2010; Ji & Wen, 2002; Tian & Jiang, 1986; Wen, 1992; Zhao et al., 1999). 1)Loreals 4 ...............................................................hainanusLoreals 2 ............................................................................. 2 2)Supralabials 6 ..................................................................... 3 Supralabials 8 .guangxiensis(further diagnosed to two subspecies at 4)...................................................................4 3)Frontonasal single, upper head scales smooth ...berdmoreiFrontonasal divided, upper head scales keeled or striated ......................................................................................sinicus 4)Midbody scale rows <30, paravertebral scales ≥49 ..............................................................................g. guangxiensisMidbody scale rows ≥30, paravertebral scales ≤47.................................................................................. g. hongjiangensis NOMENCLATURAL ACTS REGISTRATION The electronic version of this article in portable document format represents a published work according to the International Commission on Zoological Nomenclature (ICZN);hence, the new names contained in the electronic version are effectively published under thatCodefrom the electronic edition alone (see Articles 8.5-8.6 of theCode). This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank Life Science Identifiers (LSIDs) can be resolved and the associated information can be viewed through any standard web browser by appending the LSID to the prefixhttp://zoobank.org/.Publication LSID:urn:lsid:zoobank.org:pub:6C1DA4C8-9002-4D2F-B638-0EC701 AF3A5ATropidophorus guangxiensis hongjiangensisLSID: urn:lsid:zoobank.org:act:A1382C7B-749B-4E6D-9B92-1B2653 CD0A28 SCIENTIFIC FIELD SURVEY PERMISSION INFORMATION Permission for field surveys was granted by the Administration Bureau of the Xuefeng Mountain National Forest Park and the Administration Bureau of the Daming Mountain National Nature Reserve of Guangxi. SUPPLEMENTARY DATA Supplementary data to this article can be found online. COMPETING INTERESTS The authors declare that they have no competing interests. AUTHOR CONTRIBUTIONS K.J.G., S.C.S., S.D.C., and X.J.D. conducted the field work.S.D.C. performed the molecular experiments and data analyses. K.J.G. and F.S. examined the morphology and prepared the manuscript. X.J.D., N.F.W., S.C.S., and M.H.revised the manuscript. All authors read and approved the final version of the manuscript. ACKNOWLEDGEMENTS We thank Jia-Wu Xiong, Yong Shu, Zhe Huang, Yang-Jing Liu, Yan Huang, Tong Zhang, and Zi-Jun Tang (Central South Inventory and Planning Institute of National Forestry and Grassland Administration) for their support during field work.We are grateful to Wei-Ping Xia, He-Cheng Yi (Administration Bureau of Xuefeng Mountain National Forest Park), Ning Huang, Li Lin, Qi-Lin Pan, Bo Xie, Yan-Wu Lu, and Jian-Chun Li (Administration Bureau of Daming Mountain National Nature Reserve of Guangxi) for permission to conduct the field survey and for support and assistance in research. We are sincerely grateful to Qi-Ping Hu, Director of the Department of Cell Biology and Genetics (formerly Department of Biology),Guangxi Medical University, for providing important information about the type specimens ofT. g. guangxiensis.We also thank Cai-Chun Yi (Changsha Environmental Protection College) for helping to revise the English.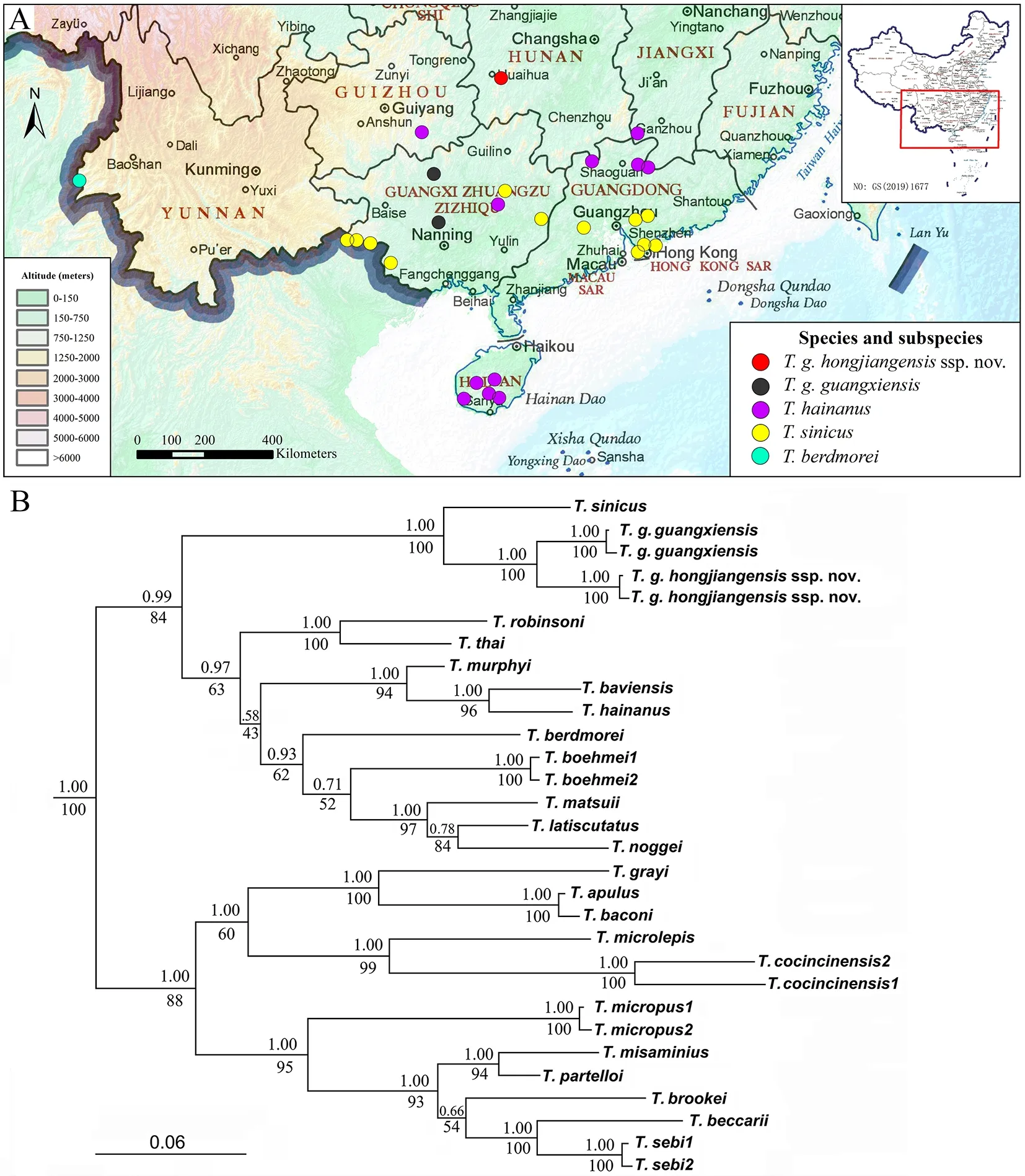
杂志排行

Zoological Research的其它文章
- Phylogenomic relationships and molecular convergences to subterranean life in rodent family Spalacidae
- Prosecution records reveal pangolin trading networks in China, 2014-2019
- Chromosome-scale genome assembly of brownspotted flathead Platycephalus sp.1 provides insights into demersal adaptation in flathead fish
- Deletion of phosphatidylserine flippase β-subunit Tmem30a in satellite cells leads to delayed skeletal muscle regeneration
- Role of juvenile hormone receptor Methoprene-tolerant 1 in silkworm larval brain development and domestication
- Captopril alleviates lung inflammation in SARS-CoV-2-infected hypertensive mice