小麦DON毒素研究进展
2021-10-18阮双司红起
阮双,司红起
安徽农业大学农学院,合肥230036
脱氧雪腐镰刀菌烯醇(DON)是一类B型单端孢霉烯族真菌毒素,具有典型的细胞毒性、免疫毒性和遗传毒性,于20世纪70年代在禾谷镰刀菌(Fusarium graminearum)侵染后的小麦、大麦等禾谷类作物上被发现并鉴定,是全球污染率较高的真菌毒素[1]。DON毒素是世界上污染面积最大、污染量最高的真菌毒素之一,其分布广泛、毒性强、传播快,主要寄存于小麦、大麦、玉米等谷类作物生产、收获、运输和仓储等过程中[2]。因DON毒素在牧畜业中会导致猪的拒食和呕吐,所以又被称为“呕吐毒素”[3]。在分子水平上,DON通过与核糖体结合来抑制蛋白质合成,参与分化、凋亡等相关信号转导过程,并激活关键细胞激酶,导致植物正常细胞破坏。近年来,由于各种环境因子的影响,世界各地DON污染越来越严重,在小麦、大麦、玉米等谷物中检测出大量DON毒素,其含量严重超出限量标准,对人和动物健康造成极大威胁。因此,本文就DON毒素机理以及小麦DON积累抗性等方面的研究进展进行了综述,以期为DON毒素的防控提供参考。
1 DON毒素
1.1 DON毒素理化性质
DON纯品为白色针状结晶,分子式为C15H20O6,化学名称为3α,7α,15-三羟基-12,13环氧单端孢霉-9-烯-8-酮(3,7,15-trihydroxy-12,13-epoxytrichothec-9-en-8-one),相对分子质量296.32,熔点151~153℃[4]。DON理化性质较稳定,在紫外线照射下没有荧光,易溶于水和极性溶剂(如甲醇、乙醇、氯仿、乙酸乙酯等),不溶于乙醚和正己烷[5]。DON在碱性条件下不稳定,但对高压、高热具有较强的耐受性,121℃高压加热25 min仅有少量毒力破坏。且在弱酸中不分解,加碱及高压处理可以破坏其部分或全部毒素。当pH为4.0时,170℃加热60 min,仅少量毒素被破坏;pH为10.0时,高压100℃加热60 min,DON毒素部分被破坏,120℃加热30 min或170℃加热15 min,DON毒素完全被破坏[6]。此外,DON具有较强的耐藏力,在病麦中4年,仍能保留原有的毒性[7]。
1.2 DON毒性机理
DON是由病原体禾谷镰孢菌产生的一种次级代谢产物,主要来源于单端孢霉烯族化合物,如尖孢镰刀菌、粉红镰刀菌等。主要产毒真菌为禾谷镰刀菌和黄色镰刀菌等,这些霉菌极易生长但难以控制[8]。DON毒性主要是通过氧化应激(oxidative stress,OS)引起,氧化应激是最重要的毒性机制之一,能够产生自由基,包括活性氧等,并诱导脂质过氧化,破坏细胞膜完整性、使细胞氧化还原信号和细胞抗氧化状态发生改变,最终造成DNA损伤和细胞死亡[9]。DON可以产生广泛的毒性效应,造成人与动物食物摄入量减少,营养效率和免疫效果降低。主要表现为明显的急性和慢性毒性作用,急性DON中毒会引发脂肪肝变性、伴随厌食症、低血糖和反常的瞬时胰岛素释放,造成代谢紊乱、非酒精性脂肪肝疾病等的发生[10];慢性DON中毒会引起拒食、呕吐、腹泻、发烧、反应迟钝、体重减轻等症状,严重时会损害造血系统导致死亡[11]。DON的高发病率令人担忧,因为DON能与其他真菌毒素如玉米赤霉烯酮(zearalenone,ZEN)及其隐蔽形式产生协同或叠加效应,对人和动物健康造成更大的威胁[12]。
此外,DON可以干扰肠道微生物和宿主线粒体之间协调的双向串扰,这可能与其诱导的肠道毒性有关[13]。通过DON处理可抑制仔猪海马神经细胞(PHNC)增殖,并导致线粒体膜电位降低、线粒体细胞色素(CYCS)和细胞凋亡诱导因子(AIF)的释放等,说明DON通过线粒体凋亡途径诱导PHNC细胞凋亡[14]。迄今为止,所有被评估的动物物种都易受到DON毒素的影响。根据动物毒力学试验,不同动物对DON的敏感程度不一:猪最敏感,其次为小鼠、大鼠、家禽等[15]。不同物种DON代谢物也不同(表1),除母体化合物DON外,其乙酰化真菌代谢物3-乙酰脱氧雪腐镰刀菌烯醇(3Ac-DON)、15-乙酰脱氧雪腐镰刀菌烯醇(15Ac-DON)以及植物来源的DON-葡萄糖苷(DON-3G)和细菌产物脱环氧基-DON(DOM-1)对人和动物也造成了一定影响[16]。研究表明,DON和15Ac-DON的毒性明显高于3Ac-DON和DON-3G[17]。其中,3Ac-DON可引起显著的氧化损伤,表现为天门冬氨酸基转氨酶(AST)、丙氨酸转氨酶(ALT)、乳酸脱氢酶(LDH)升高,过氧化氢酶(CAT)和超氧化物歧化酶(SOD)降低,过氧化氢(H2O2)和丙二醛(MDA)等含量增加;3Ac-DON通过内质网应激(endoplasmic reticulum stress)诱导动物肝脏死亡,恢复内质网应激功能可能是减少3Ac-DON对动物肝脏危害的预防策略[18];结合产 物DON-15-葡 萄 糖 醛 酸(DON-15-GlcA)、DON-3-葡萄糖醛酸(DON-3-GlcA)、游离DON是人类DON的生物标志物,这些毒素可通过母亲的胎盘屏障进入胎儿体内,抑制胎儿生长并降低免疫功能;且儿童和青少年的DON暴露量高于成年人,老年人DON暴露量最低,经常面食摄入者的DON暴露量要远高于稻米摄入人群,该研究为毒性伤害提供了早期预警[19]。

表1 不同物种DON代谢物Table 1 The DON metabolites of different species
1.3 DON毒素污染特征
DON毒素被联合国粮农组织(Food and Agriculture Organization of the United Nations,FAO)和世界卫生组织(World Health Organization,WHO)认定为最危险的自然发生的食品污染物[20]。DON污染在全球各地均有发生,在我国发生较为普遍,尤以温暖多雨、气候潮湿地区最为严重,其中华南、华中和华北地区的污染程度明显高于其他地区[21]。DON不仅污染谷类作物及其粮食制品,在动物的奶、蛋中也均发现有DON残留。此外,呕吐毒素依旧是污染最为严重的毒素类型,在麸皮中污染率可达90%~100%,半数以上超出国标限量值[22]。
1.4 小麦DON毒素分布规律及常用检测方法
小麦中DON分布呈现籽粒间和籽粒内外层间不均两个特征,籽粒间呕吐毒素主要分布在赤霉病粒和其他干瘪不饱满籽粒中;籽粒内外层间,呕吐毒素主要分布在小麦的外表皮,并且由外向内逐渐减少[23]。其中,小麦麸皮呕吐毒素含量最高,皮磨粉次之,心磨粉DON含量最低。小麦粉与小麦籽粒相比,呕吐毒素下降了413.1~921.7 μg·kg-1[24]。由于DON毒素已经严重影响小麦粮食安全,对人畜健康产生极大威胁,所以全球各个国家对小麦及其制品的DON含量已制定相关限量标准(表2),我国小麦及其制品DON毒素限量为1.0 mg·kg-1。
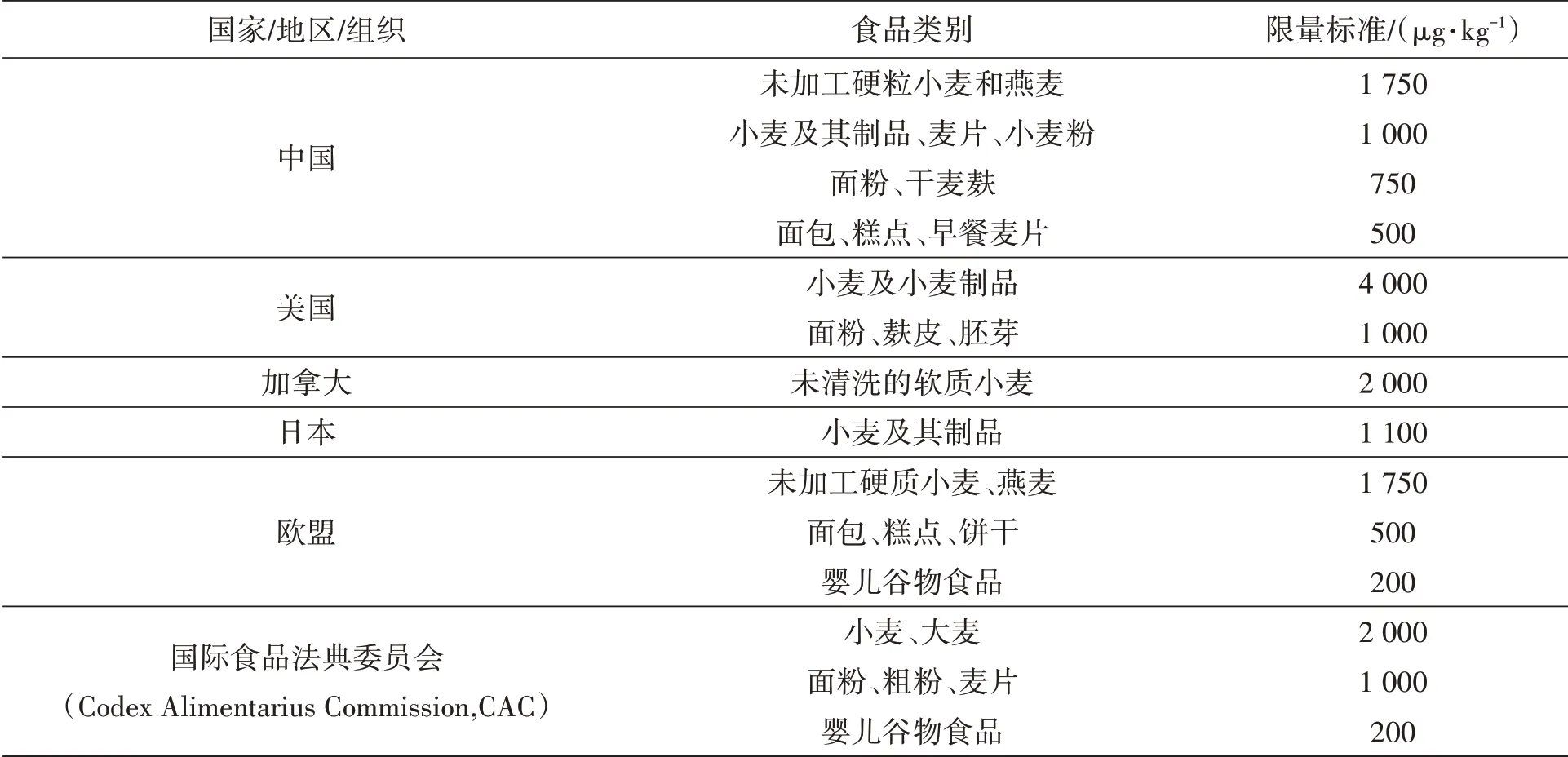
表2 部分国家/地区或组织对小麦及其制品的DON毒素限量标准[25-26]Table 2 The limit standard of DON toxin for wheat and its products in some countries,regions or organization[25-26]
目前,呕吐毒素的检测方法有很多,主要有同位素稀释液相色谱-串联质谱法(isotope dilution liquid chromatography/tandem mass spectrometry,ID-LC/MS/MS)、免疫亲和层析净化高效液相色谱法(immunoaffinity chromatography high performance liquid chromatography,IAC HPLC)、薄层色谱测定法(thin layer chromatography,TLC)和酶联免疫吸附测定法(enzyme-linked immuno Sorbent assay,ELISA),它们具有不同的检测特点,为DON毒素的测定提供了极大的便利(表3)。其中,同位素稀释液相色谱-串联质谱法操作简便、快速,具有很高的准确度和灵敏度,适用于粮食产品中多组分真菌毒素的同时检测,具有较高的实际应用价值[27];免疫亲和层析净化-高效液相色谱法具有检出限低、精密度高、回收率接近真实值、重现性好、操作简便等优点[28];薄层色谱测定法具有制备样品最少或不制备样品、操作简单、经济有效、容量大、可同时分离多个样品等独特优点[29];酶联免疫吸附测定法具有灵敏度高、特异性好、操作简便、检测成本低、一次性检测样本量大等特点,能够更好地满足我国食品企业、政府监管部门等开展检测工作[30]。此外,还有高效液相色谱-紫外法(HPLC-UV),该方法可同时检测小麦DON、15AC-DON和3AC-DON,准确度高、重现性好,样品净化方法使杂峰干扰少,可大幅度减少有机溶剂的使用,成本低,适用于小麦中B型单端孢霉烯族毒素的大批量检测[31]。
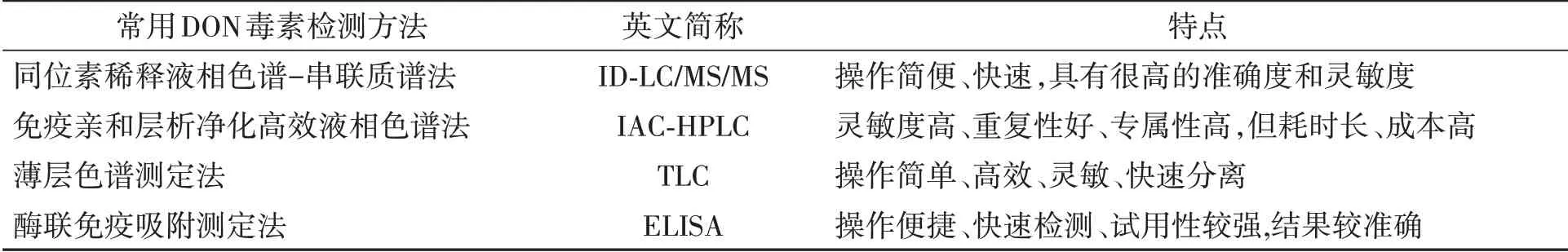
表3 常用的DON检测方法Table 3 Commonly used DON detection method
2 小麦抗DON毒素积累机制
2.1 小麦DON积累抗性遗传机理
小麦赤霉病抗性由数量性状控制,遗传基础较复杂,受环境和遗传因素的双重影响,其抗性主要分为抗初始病原体感染(TypeⅠ)、抗扩展(TypeⅡ)、抗DON毒素积累(TypeⅢ)、抗籽粒感染(TypeⅣ)、抗减产(TypeⅤ)五类。其中,TypeⅡ、TypeⅢ表型更稳定,被广泛研究,是最重要的两种小麦赤霉病抗性类型,对小麦产量和品质有直接影响,是赤霉病抗性改良的重点[32-33]。
小麦抗DON毒素积累(TypeⅢ)又分为毒素化学修饰和阻止毒素合成两类,可用小麦籽粒的毒素含量进行评估。DON是小麦籽粒中最主要的真菌毒素,也是致病因子。小麦籽粒积累DON毒素后,活性氧(ROS)含量提高,抗氧化能力下降,诱导机体发生氧化应激反应,最终导致机体和细胞的脂质过氧化和DNA氧化损伤,严重影响小麦的产量和品质[34]。若能控制小麦中DON毒素积累,就极大可能会降低小麦赤霉病危害。
研究表明,DON积累抗性狭义遗传力为74.54%,表现出较高的遗传力。其中,普通小麦DON毒素积累抗性主要表现为加性效应,部分表现为显性效应。而小麦籽粒中DON抗性遗传主要表现为显性效应。且DON含量与病小穗数、病小穗率、病粒率呈显著正相关。因此,在育种中以小麦籽粒DON含量作为赤霉病抗性选择的标准,可以在早期世代进行选择[35-36]。
2.2 小麦DON毒素生物合成途径及基因调控网络
近年来,国内外学者对小麦DON毒素合成与调控途径做了大量研究,明确小麦DON毒素的生物合成不仅受温度、湿度、pH、杀菌剂等理化因素的影响,也与一些有机物质、转录因子、蛋白激酶等有关。这些因素在DON毒素合成中还参与调控相关基因的表达,在小麦抗DON积累中起着关键性作用。
例如真菌代谢产物——多球壳菌素(myriocin)具有抑菌、免疫调节的作用,可通过破坏细胞膜的通透性,使细胞内物质释放,进而抑制小麦籽粒中镰刀菌的产毒活性,并减少了99%以上的DON、15AC-DON的合成。且myriocin参与下调TRI5基因的表达,降低了小麦DON毒性,有效控制了小麦籽粒中禾谷镰刀菌的污染[37];杀菌剂喹啉类物质可以破坏镰孢菌毒素异构酶的形成,抑制DON生物合成,使DON产量显著降低,并且抑制DON相关基因TR1、TR15的表达[38];除杀菌剂外,一些蛋白因子在影响DON毒素合成中起重要作用。小麦赤霉病菌FgRab7蛋白具有调控Tri基因簇表达和DON毒素生物合成的作用。该蛋白缺失使Tri基因表达量显著下降、DON毒素合成明显减少,说明FgRab7是影响DON合成的重要因子[39]。小麦禾谷镰刀菌中的两个Sec2同源物FgSec2A和FgSec2B位于菌丝和分生孢子的顶端区域,在小麦DON生物合成和分泌中起着至关重要的作用,其双缺失会导致DON含量大量减少[40]。小麦赤霉病菌中FgVPS26和FgRab7的激活态蛋白可以相互作用,且两者是与DON产毒相关的基因,在产毒的调控上有着密切的联系,均通过降低Tri基因的表达影响DON毒素的生物合成。FgRab7和FgVPS26也可以作为新型生物杀菌剂的作用靶标进而控制禾谷镰刀菌及其产生的DON毒素[41]。植物UDP-葡萄糖基转移酶(UGT)可以降低DON的毒性而提高赤霉病抗性,Zhao等[42]在小麦抗病品种苏麦3号中克隆了一个抗DON毒性的基因TaUGT5,位于2B染色体上,该基因过表达增强小麦胚芽对DON的抗性,可以更好的保护植物细胞结构,使细胞不受病原菌感染;同样,He等[43]也在苏麦3号中克隆并鉴定了一个赤霉病诱导高表达的新基因TaUGT6,通过酶活测定发现TaUGT6可以糖基化DON为D3G,且该基因能够通过降低DON毒性来提高植株对DON的耐受性从而提高赤霉病抗性。
此外,维生素E的抗氧化作用在一定程度上也能降低DON的毒性,尿黑酸牻牛儿基转移酶(HGGT)是维生素E生物合成的关键限速酶。HGGT基因定位于质体中,主要在小麦胚乳中表达。Wu等[44]研究克隆了HGGT基因的全长,并开发了基因特异标记,在31份小麦品种中进行标记分析,证明了HGGT基因在一定程度上阻止病原菌在穗部的扩展并降低籽粒中积累的DON毒素含量,可以用于改良小麦赤霉病抗性。该研究推测维生素E对赤霉病的降毒效应很有可能是受HGGT基因调控的作用。
2.3 与DON积累抗性相关的QTL研究
迄今为止,有关小麦DON积累抗性相关的QTL研究有很多,它们分布于小麦不同染色体上,表型变异率大小不一,表现不同程度的抗性(表4)。

表4 部分与DON积累抗性相关的QTLTable 4 Some QTLs related to DON accumulation resistance
对于小麦DON毒素抗性的QTL研究,因其所使用的定位方法、小麦遗传资源、生态环境不同而使定位结果也不同。挖掘与DON积累抗性相关QTL有助于了解抗赤霉病性状的遗传和调控机制,丰富优异基因资源,为改良和选育小麦抗病品种奠定分子基础。Somers等[55]利用小麦抗病品种武汉1号和Maringa杂交株系为材料,构建了91个衍生双单倍体系,在2DS、3BS和5AS染色体上定位到抗DON毒素积累的QTL,且在3BS和5AS上携带有利等位基因的品系,其DON毒素积累量降低 了17%;Kassa等[46]以 小 麦Arinax品 种 与NK93604杂交建立的93个DH株系为材料,对其抗赤霉病与DON积累进行了QTL定位,检测到4个与之相关的QTL,位于1AL、1BL、6BS、7AL染色体上,分别可解释27.9%、19.6%、7.8%、14.8%的表 型 变 异;Jiang等[56]利 用 小 麦 品 种Veery与CJ9306构建重组自交系群体,鉴定了两个新的QTL——QFhs.nau-2DL和QFhs.nau-1AS,与 抗DON毒素积累相关。其中,QFhs.nau-2DL可以用于分子标记辅助选择,以提高对DON积累的抗性;Abate等[49]通过Erin和Mo94-317杂交构建重组自交系群体,在3BS、4BL和5A染色体上鉴定了3个可降低DON毒素含量的QTL,可解释31%的DON表型变异;中国小麦品种Ning7840作为抗性亲本已被世界各地广泛使用,该种质包含一个抗赤霉病数量性状位点Fhb1,可解释高达53%的分离群体表型变异,Bernardo等[57]利用Ning7840与Clark构建BC7F2单株,证明了Fhb1可以降低DON含量,在美国冬小麦品种中可作为有效的抗FHB育种资源;Islam等[58]利用小麦品种Truman的近等基因系,在2ASc和2DS上定位到了两个可以降低DON毒素含量的QTL;Cristiano等[59]利用小麦品种Everest和WB-Cedar杂交获得176个双单倍体(DH)群体,在1BS、3DS、4BL和5AS染色体上鉴定了4个来自Everest的QTL,其中最大效应的QTL(Qksu-fhbE-5AS)可解释DON变异的13%,且携带所有标记的DON积累的QTL中的DH株系,其DON含量下降了36%,这也是首次利用SKNIR(single-kernel near-infrared)技术确定了与DON低积累相关的QTL,为今后的遗传育种研究提供了支持;Goddard等[60]利用2个定位群体(Anahuac 75×BR 18-Terena、BR 18-Terena×BRS 179)进行表型和QTL分析,在4B染色体上发现了一个抗DON积累相关的新主效QTL,占表型方差的17.8%,可作为育种的有效目标,用来改善籽粒品质和减少对杀菌剂的依赖;Zheng等[61]对625个与抗扩展和抗DON毒素积累相关的QTL进行元分析与多组学联合分析,筛选到17个位于区间内同时具有转录组和蛋白质组数据支持的基因,为后续目标QTL克隆和抗性机理解析提供候选基因,也为建立一个全面的小麦抗赤霉病分子育种平台奠定了基础。
值得一提的是,与小麦赤霉病抗性相关的数百个QTL中,只有7个位点被明确为小麦抗赤霉病基因,即Fhb1~Fhb7,并在小麦遗传育种中应用[62]。其中,Fhb1和Fhb7基因与DON毒素抗性相关。最新研究表明Fhb1基因编码一个注释为富含组氨酸的钙离子结合蛋白(histidine-rich calcium-binding-protein,His),由该蛋白基因3’外显子罕见缺失引起。Fhb1基因已克隆,并具有较强的抗扩展性,可降低籽粒中DON毒素的积累,其不仅可以提高小麦、拟南芥对FHB的抗性,也可用于改良其他易受禾谷镰孢菌感染的植物[63];Fhb7来源于长穗偃麦草(Thinopyrum ponticum),是抗FHB的主效基因,位于染色体7E长臂末端。该基因编码谷胱甘肽转移酶(GST),并已被克隆。将Fhb7基因导入到小麦品种中,可以降低小麦籽粒中DON毒素积累、产生解毒效应,显著提高小麦FHB抗性;此外,该基因可用于小麦赤霉病遗传育种,并在生物医学、粮食深加工、饲料和食品工业中具有潜在用途和广阔的市场前景[64]。
3 展望
禾谷镰刀菌是小麦赤霉病最主要的病原真菌,其产生的DON毒素是小麦中检出量最高,污染面积最大的真菌毒素之一,已严重危害人和动物健康。因此,减少DON污染对人畜的危害以及经济损失是当前各国政府高度关注的问题。对于DON毒素的生物合成与调控途径已有大量研究,物理、化学和生物防治也取得一定成效,但是还需进一步加强各种措施的综合应用,才能更好地防治DON毒素的污染。其次,关于DON毒素抗性的QTL定位和基因有很多,也为DON抗性机制的理解及赤霉病育种材料的创制奠定了一定的基础,但实际应用到小麦育种上的却很少。而培育与利用抗DON毒性及DON含量低的品种是控制赤霉病、减轻DON危害最为经济有效且对环境友好的途径。因此,更要大力开展对禾谷镰刀菌DON毒素基因调控网络的系统研究,充分阐明各基因间的相互联系和作用功能,才能更好地培育抗DON毒性且籽粒DON含量低的品种,并进一步开展聚合育种,以此减少DON对小麦以及其他农作物的污染水平,提高农产品的品质,保障粮食安全。
