3 种诱食剂对斑点叉尾鮰生长和生理的影响
2021-10-18易皓明叶元土石瑶瑶萧培珍
■易皓明 叶元土* 吕 昊 吕 斌 石瑶瑶 孙 飞 郁 浓 萧培珍
(1.苏州大学基础医学与生物科学学院,江苏苏州 215123;2.北京桑普生物化学技术有限公司,北京 100162)
斑点叉尾鮰又称沟鲇,属于鲇形目、鮰科鱼类,1984年从美国引入到中国,具有食性杂、生长快、适应性广、抗病力强、肉质上乘等优点。近年来我国水产养殖业发展迅速,2018 年我国水产养殖总产量占到了世界的60%,2019 年我国鮰鱼养殖总产量达到29.77 万吨。随着全国水产养殖模式升级、规模壮大和集约化程度的提高,饲料的需求量也在不断增加,在水产养殖中饲料成本占到了65%左右。如何在养殖过程中有效降低饲料成本具有重要研究意义,如通过开发新的价格低廉的植物蛋白源替代动物蛋白源,但饲料中使用植物蛋白源往往会引起饲料的适口性差、摄食效果不佳等问题。研发并应用促进水产动物摄食的诱食剂物质是一种行之有效的方法,可显著提高饲料诱食性、适口性和饲料转化率等。
诱食剂又称促食剂、食欲增进剂或摄食促进物质,适宜地添加在饲料中可改善适口性、增加摄食量和提高消化吸收率的一类化学刺激物。已有研究表明,对水产动物具有诱食作用的物质有氨基酸及其混合物、动植物提取物、生物碱(甜菜碱等)、核苷酸类物质、含硫有机物等。目前,诱食剂在甲壳类和鱼类中均取得较好的促生长效果,研究表明,诱食剂的诱食效果因鱼种类和诱食剂化学性质不同而呈现出差异。氧化三甲胺(TMAO)是存在于海洋鱼类中的一种白色结晶物质,具有鲜味而常被用于诱食剂添加到水产动物饲料中,同时它也可以作为一个评判水产动物新鲜度的指标。饲料中添加TMAO对三角鲂(Megalobrama termi⁃nalis,1846)生长影响的结果显示,添加0.082%的TMAO能显著促进三角鲂的生长。二甲基-β-丙酸噻亭(DMPT)是一种对水产动物具有诱食作用的硫化物,对鱼类嗅觉传导途径和机体免疫有一定的促进作用,另外可作为甲基供体参与机体渗透压调节,主要来源于海藻和海洋动物中。酶解乌贼粉(ESP)主要是由乌贼或者是鱿鱼的内脏经细化、过滤、杀菌、烘干等加工得到的,不仅富含牛磺酸,还因含有特殊的芳香而作为水产动物诱食剂添加在饲料中。基础饲料中添加DMPT 诱食剂对南美白对虾(Litopenaeus Vannamei,1931)的摄食和生长影响的研究表明,DMPT的最适宜添加量为0.30 g/kg,且比10%乌贼膏的诱食效果更好。本试验选用了3种诱食剂,以期通过添加到饲料中对斑点叉尾鮰的生长速度、血清生化指标、鱼体成分以及抗氧化能力的影响,筛选出能促进斑点叉尾鮰摄食、改善其生理的诱食剂及最适宜添加量,从而为斑点叉尾鮰诱食剂饲料的大规模生产和应用提供理论基础。
1 材料和方法
1.1 试验鱼
斑点叉尾鮰采购自江苏省盐城市射阳县黄沙港镇,为当年同一批次的鱼种,规格统一,体质健壮,无病无伤,选取体重在(50±0.78)g 的鱼种,随机分成16 组,每组设置3重复,每重复投放40尾斑点叉尾鮰鱼,合计为1 920尾。
1.2 试验组别设计和原料来源
3 种诱食剂分别为氧化三甲胺(Trimethylamine N-oxide,TMAO,纯度≥98%)、二甲基-β-丙酸噻亭(Di⁃methyl-β-propiothetine,DMPT,纯度≥98%)和酶解乌贼粉(Enzymatic Squid Powder,ESP,营养组分见表1),均来源于北京桑普生物化学技术有限公司。其中C为对照组;添加量为0.05%、0.10%、0.15%和0.20%的TMAO 组 分 别 设 为T1、T2、T3 和T4 组;添 加 量 为0.01%、0.02%、0.03%和0.04%的DMPT 组分别设为D1、D2、D3 和D4 组;添加量为0.3%、0.5%、0.7%和0.9%的ESP组分别设为E1、E2、E3和E4组。基础日粮配方及营养水平见表2。
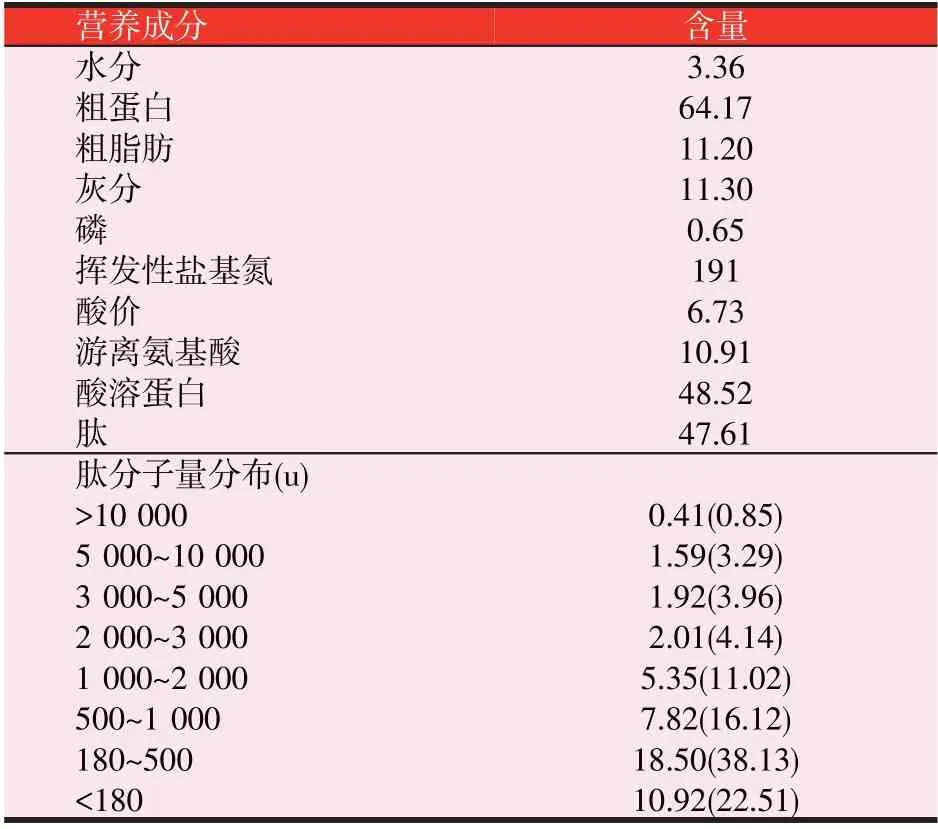
表1 酶解乌贼粉营养成分组成(g/100 g,干物质基础)
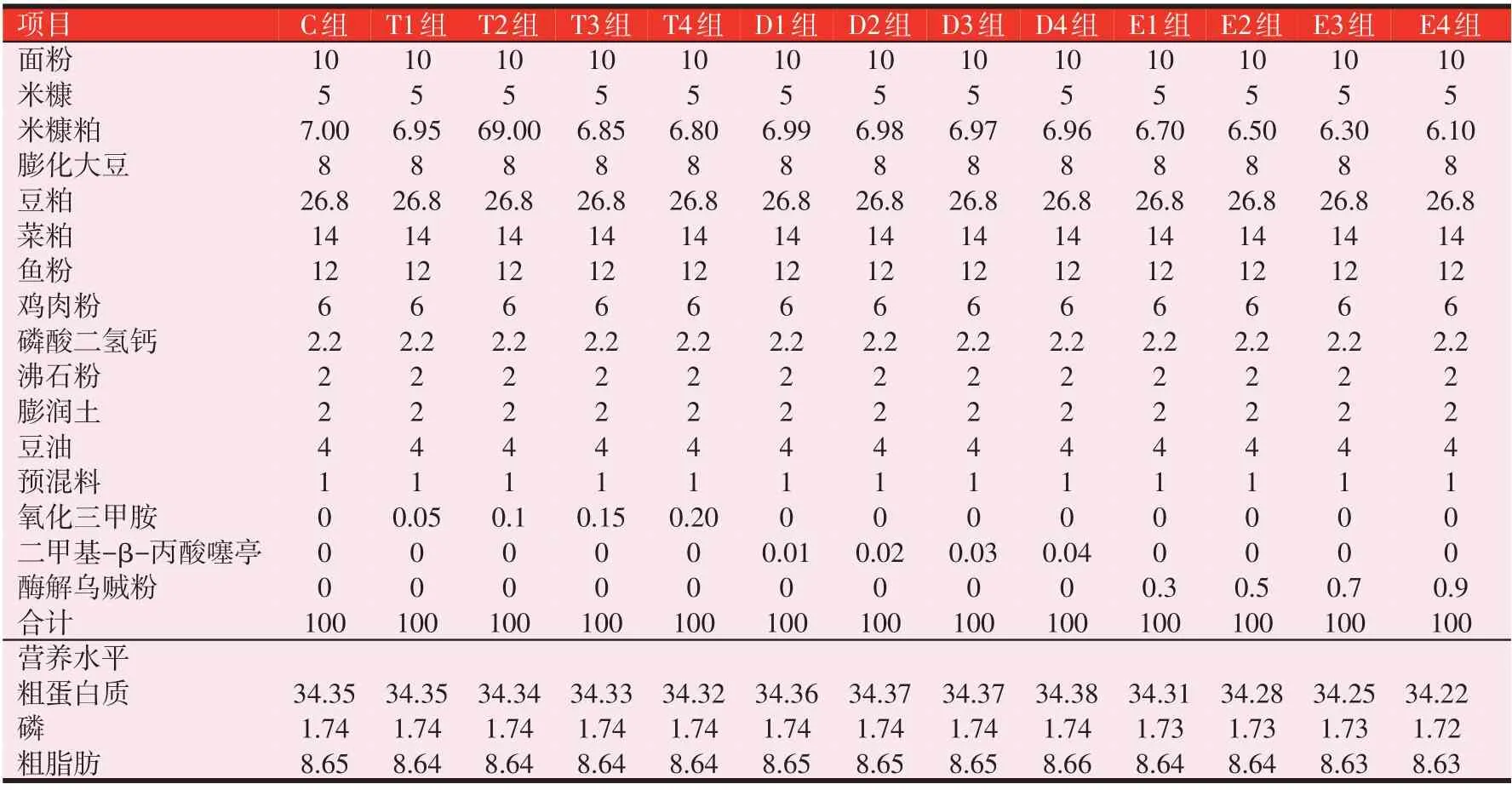
表2 基础日粮组成和营养水平(%,风干基础)
饲料原料经粉碎过60目筛,各原料按表2的要求称取,原料混合均匀后用小型环模制粒机(温度65 ℃)制成直径1.5 mm,长2~3 mm的颗粒状饲料,水分风干至13%左右,置于-20 ℃冰箱保存备用,使用前按需要量取出饲料自然解冻后投喂。
1.3 饲料营养成分和鱼体体成分的测定
鱼体水分含量的测定按GB 5009.3—2016《食品中水分的测定》的规定执行;鱼体中粗蛋白质含量的测定按GB 5009.5—2016《食品中蛋白质的测定》的规定执行;鱼体中粗脂肪含量的测定按GB 5009.6—2016《食品中脂肪的测定》的规定执行,鱼体中灰分含量的测定按GB 5009.4—2016《食品中灰分的测定》的规定执行。
1.4 养殖管理
运至试验基地的斑点叉尾鮰分组后在室外池塘中的网箱(规格为长1.5 m×宽1.5 m×深2 m)里养殖,鱼体均重为(50±0.78) g,随机分成16 个组,每组设置3 个重复,每重复投放40尾斑点叉尾鮰。对照组的斑点叉尾鮰饲喂基础饲料,其他组饲喂相对应的诱食剂饲料,日投饵量为鱼体重的3%~5%,每天06:00 和17:00分别投喂1次。试验期间网箱水温为26~35 ℃,溶解氧浓度控制在7~8 mg/L,氨氮浓度小于0.48 mg/L,亚硝酸盐氮浓度小于0.07 mg/L,pH为7.2~7.6。
1.5 样品采集与分析
8周的饲养试验结束后对斑点叉尾鮰禁食24 h后采样,之后以重复为单位称重计数。每重复随机取5尾鱼,用冰袋麻醉,称重后分离出内脏、肝脏、脾脏,并分别称重;每重复需另随机取10尾鱼进行尾静脉取血,离心机在4 ℃及转速3 500 r/min的条件下离心10 min,收集血清,用液氮迅速冷却后于-80 ℃冰箱保存。从每重复中抽取2尾保留全鱼样本,用于测定鱼体常规体成分含量。
增重率(WGR,%)=100×(W末-W始)/W始
特定生长率(SGR,%/d)=100×(lnW末-lnW始)/t
肥满度(CF,g/cm3)=100×W末/L3
饲料系数(FCR)=Wf/(W末-W始)
成活率(SR,%)=100×N末/N始
脏体指数(VSI,%)=100×W内/W
肝体指数(HSI,%)=100×W肝/W
式中:t——饲养天数(d);
N始——初始尾数(尾);
N末——终末尾数(尾);
L——终末体长(cm);
Wf——总摄食饲料的干重(g);
W末——终末均重(g);
W始——初始均重(g);
W内——内脏重(g);
W肝——肝脏重(g);
W——体重(g)。
血清指标中的谷丙转氨酶(cerealthirdtransami⁃nase,ALT)、谷草转氨酶(aspartate transaminase,AST)、碱性磷酸酶ALP(alkaline phosphatase,ALP)活性及总蛋白(total protein,TP)、白蛋白(albumin,ALB)、球蛋白(globulin,GLOB)、胆固醇(cholesterol,CHOL)、三酰甘油(triglyceride,TG)、高密度脂蛋白(high density lipo⁃protein,HDL)和低密度脂蛋白(low density lipoprotein,LDL)的含量采用苏州市九龙医院的雅培C800全自动生化分析仪测定。超氧化物歧化酶(superoxide dis⁃mutase,SOD)和丙二醛(malonic dialdehyde,MDA)均采用南京建成生物工程研究所生产的试剂盒测定。总抗氧化力(total antioxidant capacity,T-AOC)采用北京索莱宝科技有限公司生产的试剂盒测定。
1.6 数据处理
Excel 2016初步整理原始数据,用SPSS 22.0软件进一步分析。采用单因素方差分析(one-way ANOVA),之后进行Duncan’s 多重比较检验差异性。最终数据以“平均值±标准差(mean±S.D.)”表示,以P<0.05为显著差异水平。
2 结果与分析
2.1 3种诱食剂对斑点叉尾鮰鱼体成分的影响(见表3)
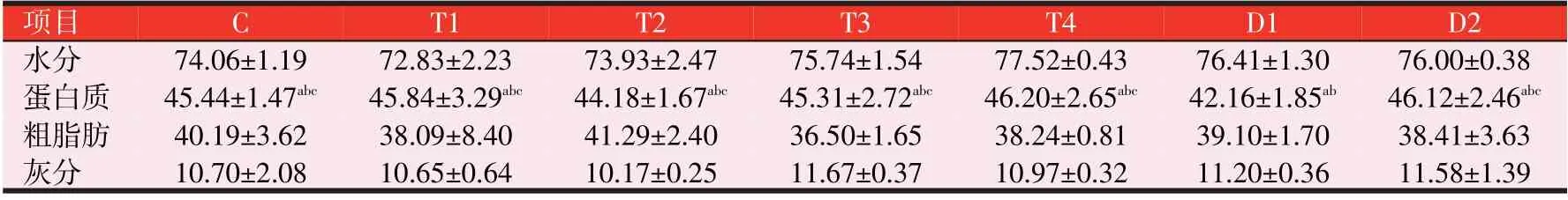
表3 3种诱食剂对斑点叉尾鮰鱼体成分的影响(%)
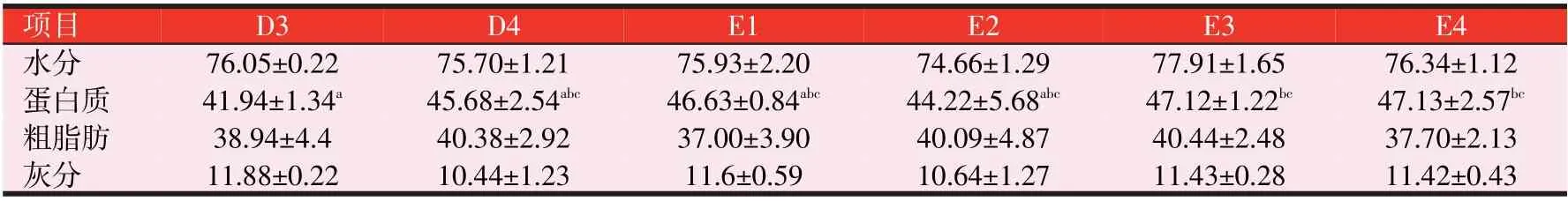
表3(续) 3种诱食剂对斑点叉尾鮰鱼体成分的影响(%)
由表3可知,各试验组的斑点叉尾鮰鱼体水分含量均无显著性差异(P>0.05);干物质中,与C组比较,各试验组的斑点叉尾鮰鱼体粗蛋白质、粗脂肪含量和灰分含量均无显著性差异(P>0.05)。
2.2 3种诱食剂对斑点叉尾鮰生长性能的影响(见表4)
由表4可知,SGR,与C组比较,各组均没有显著性差异(P>0.05);但T组中T2的SGR上升了4.22%,T3上升了1.20%;E组中E1的SGR上升8.43%(P>0.05),E2、E3和E4的SGR上升3.61%~4.82%(P>0.05);FCR,与C组比较,E组的FCR无显著性差异(P>0.05);T组中T4的FCR 升高了17.65%;D 组中的D4 的FCR 升高了16.81%(P<0.05)。形体指标,与C组比较,D1组的HSI有显著差异(P<0.05),其他各组均无显著性差异;T组的T3、T4及D组和E组的CF均显著下降(P<0.05)。T组的T1、T3和T4的VSI显著下降(P<0.05),且D组的D2、D3和D4及E组的E1、E3和E4亦是如此(P<0.05)。
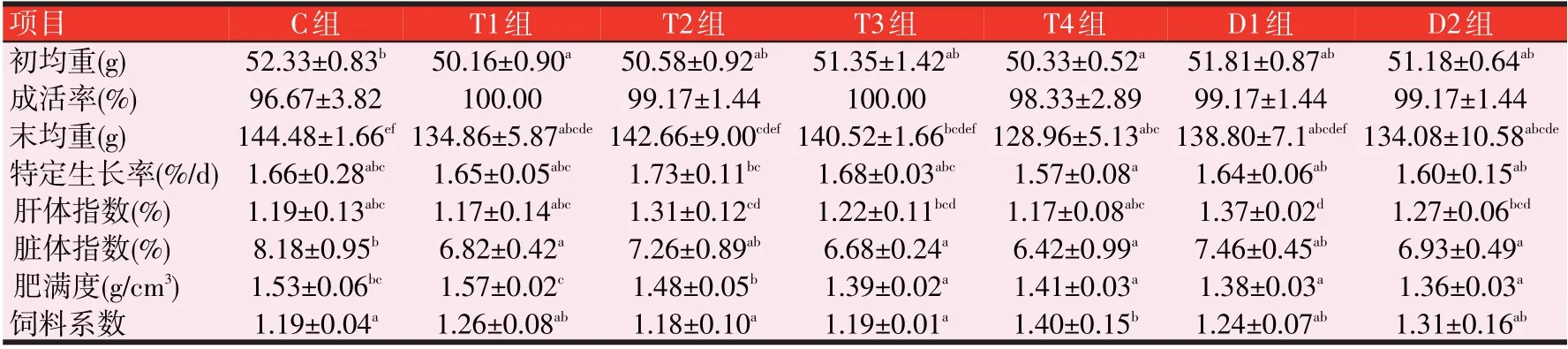
表4 3种诱食剂对斑点叉尾鮰生长性能的影响
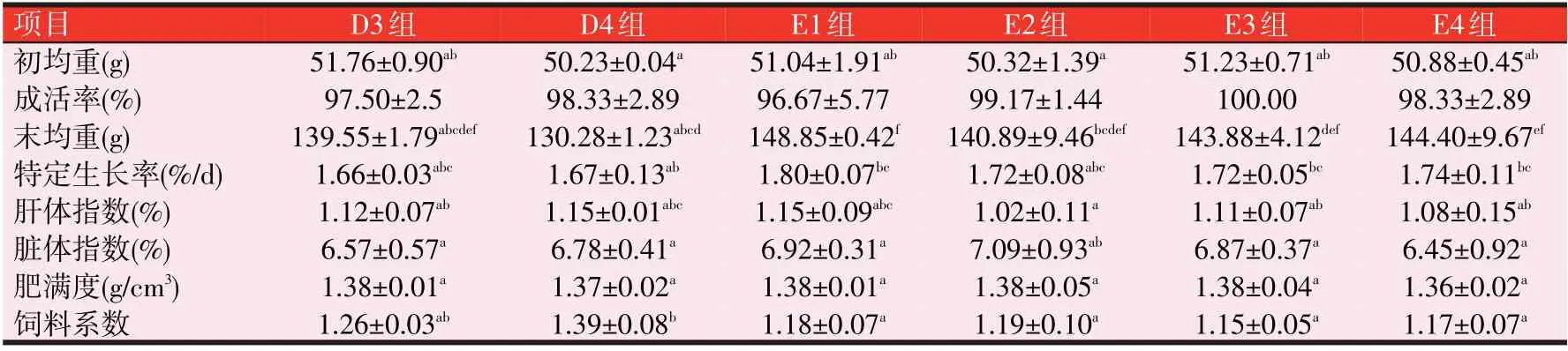
表4(续) 3种诱食剂对斑点叉尾鮰生长性能的影响
2.3 3种诱食剂对斑点叉尾鮰血液指标的影响(见表5)
表5 可知,与C 组比较,T、D 和E 组的TP、ALB、GLOB、ALT、AST、ALP、CHOL和TG无显著性差异(P>0.05),T、D 和E 组TP、ALB 和GLOB 均有不同程度的升高(P>0.05),T 组T4 的TP、ALB 和GLOB 分别升高了10.34%、14.54%和8.38%,D 组D4 的分别升高了18.52%、22.87%和16.45%,E 组E1 的分别升高了21.69%、24.35%和20.42%。CHOL:各组均有不同程度的升高,其中T组随着添加量的增加呈现出先升高再降低的趋势,T3 升高了25.27%(P>0.05);D 和E 组随着添加量的增加呈现出先升高再降低再升高的趋势,D1和E4均升高了43.90%(P>0.05)。TG:T组T2、T3和T4随着添加量的增加呈现出先升高再降低的趋势,T3升高了13.59%(P>0.05)。D1升高了39.11%(P>0.05);HDL:T 组升高了9.14%~20%(P>0.05);D1、D2和D3升高了18.29%~33.71%(P>0.05),D4显著升高了50.86%(P<0.05);E1、E2 和E3 升高20.57%~40.57%(P>0.05),E4 显著升高57.71%(P<0.05)。LDL:E2、E3和E4升高(P>0.05),E1显著性升高(P<0.05)。
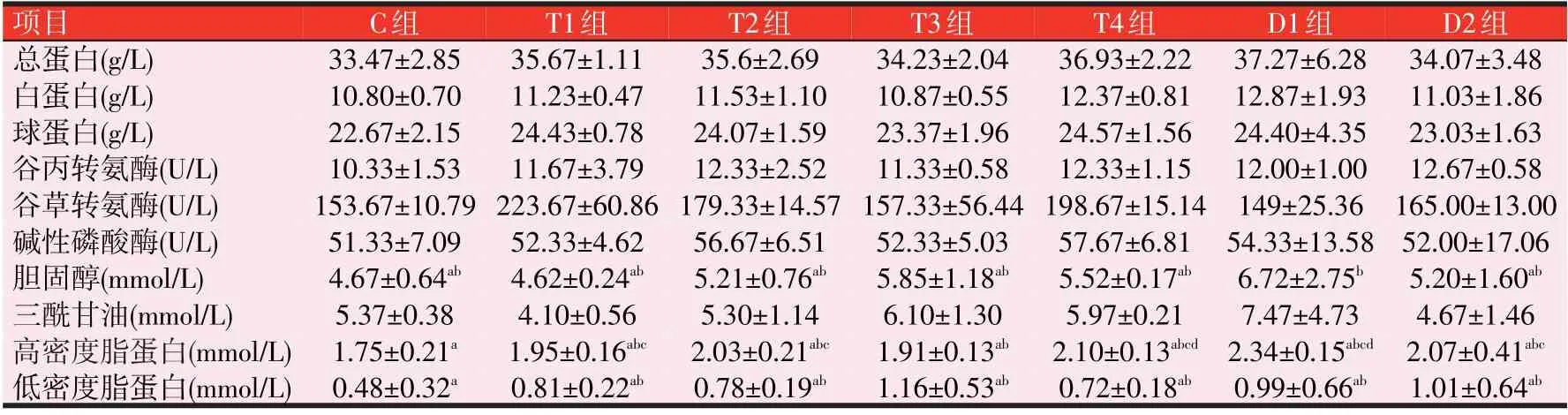
表5 3种诱食剂对斑点叉尾鮰的血清生化指标的影响
2.4 3种诱食剂对斑点叉尾鮰抗氧化能力的影响(见表6)
由表6可知,斑点叉尾鮰血清的T-AOC:与C组比较,各组均无显著性差异(P>0.05),T 组中T3 和T4 提高了32.61%,D 组中D2 和D4 分别提高了17.39%、26.09%。E组有一定的上升趋势,随添加量的增加而升高,提高了10.87%~84.78%。SOD:与C组比较,E组中E1、E2 差异不显著(P>0.05),E3、E4 存在显著性差异(P<0.05);T和D组均无显著性差异(P>0.05)。血清MDA:与C组比较,T组的T1、T2、T3和T4均显著下降(P<0.05),且随TMAO 的添加量增加而升高;D 组的D1、D2 和D4 及E 组的E2 和E3 亦是如此(P<0.05),D组和E组均随诱食剂的添加量先降低再升高。
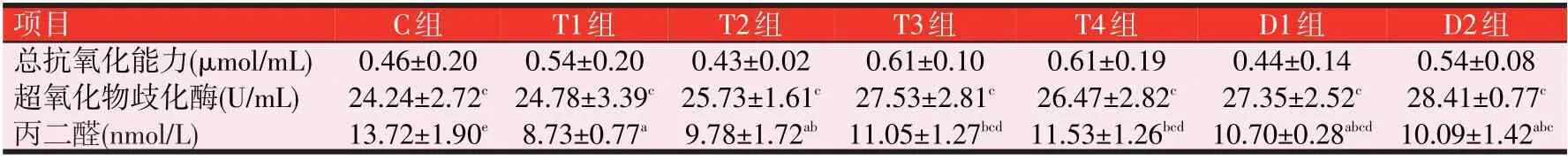
表6 3种诱食剂对斑点叉尾鮰抗氧化能力的影响
3 讨论
3.1 3 种诱食剂对斑点叉尾鮰生长性能和鱼体成分的影响
试验结果表明,饲料中添加3种诱食剂能够不同程度地促进斑点叉尾鮰的生长,这与在其他鱼虾等水产动物上的研究结果一致。诱食剂对鱼类起刺激作用的始端是鱼的嗅觉和味觉器官,鱼的味蕾遍布体内外,在口腔、唇、触须、鳃、咽、食管、体侧和鳍上均有分布,鱼类嗅觉的灵敏度与初级嗅板数量有关,嗅板数目越多则其嗅觉上皮的相对面积就大,鱼类的嗅觉也就越灵敏,斑点叉尾鮰的初级嗅板数目达25~28对,比鲤鱼和革胡子鲶等鱼类嗅觉更灵敏。本试验中,在饲料中添加0.3%的酶解乌贼粉对斑点叉尾鮰特定生长率提高了8.43%。乌贼粉具有浓郁的芳香能刺激斑点叉尾鮰的嗅觉器官而增加采食量,其中含有甘氨酸、丙氨酸、谷氨酸等游离氨基酸可有效改善饲料的适口性和利用率。乌贼粉的蛋白质含量一般在30%以上,必需氨基酸及鲜味必需氨基酸在氨基酸总量中约为50%和40%,EPA、DHA 总和高达35%,在饲料中添加适量的乌贼粉对颗粒饲料的制粒有辅助黏结的作用,还对甲壳类动物的蜕壳也具有一定的促进作用。采用酸水解、碱水解、酶解或者是菌酶协同处理方法,可以得到更容易被鱼体吸收的乌贼粉水解多肽和氨基酸。在饲料中添加了4%的富含氨基酸、小肽的虾蛄蛋白浆,能有效促进中华绒螯蟹的摄食。在攀爬鲈鱼(Anabas testudienus, 1792)饲料中添加豆粕和乌贼副产品的发酵混合物来替代鱼粉,替代比例在25%时其总产量和经济效益最高。TMAO主要来源于海洋生物,海洋生物生活环境的盐度高低与体内TMAO含量成正比,其结构与甜菜碱相似并具有特殊香味,TMAO的诱食机制是通过刺激水产动物的嗅觉器官使神经兴奋性提高。DMPT 是含硫有机物类中一种最典型的诱食剂,已有报道在饲料中添加282.78 mg/kg 的DMPT 可增加消化酶的活性,改善了生长性能和消化能力,并且增强了正在生长的草鱼的肠道结构完整性,这可能与食欲调节因子神经肽Y有关。本试验中TMAO 和DMPT 虽然对斑点叉尾鮰的特定生长率有一定升高趋势,但是作用并不显著,可能是因为鱼的种类差异,也可能是单一诱食剂对斑点叉尾鮰的诱食效果不明显,在后期将继续深入研究TMAO和DMPT组合对斑点叉尾鮰的诱食性及相关食欲基因的表达量,探究内在的促摄食机制。饲料中添加氨基酸-核苷酸-DMPT 复合诱食剂对鲫鱼的诱食效果强于单一的氨基酸、核苷酸及DMPT,但该复合
诱食剂对草鱼和乌鳢的诱食效果却不及单一的氨基酸、核苷酸及DMPT,可能是因为杂食性鱼类对化学性复合诱食剂较敏感。D组和E组的肥满度均显著下降,而TMAO 的添加量在0.15%和0.20%时的肥满度同样显著下降。饲料中TMAO 的添加量在0.05%、0.15%和0.20%时可以显著降低斑点叉尾鮰的脏体比。DMPT 的添加量在0.02%、0.03%和0.04%及酶解乌贼粉的添加量在0.3%、0.7%和0.9%时亦是如此。这表明诱食剂可以改善脂肪在斑点叉尾鮰内脏组织中的附着和沉积,有利于提高鱼体机能,促进动物体生长,与郑晶等在杂交鳢上的研究结果一致。由表3可知,3 种诱食剂对斑点叉尾鮰鱼体水分及鱼体干物质的粗蛋白质、粗脂肪和灰分含量没有显著影响,但斑点叉尾鮰的全鱼粗脂肪含量随着饲料中3 种诱食剂的添加量的增加呈现下降趋势,说明3种诱食剂可能对斑点叉尾鮰脂肪沉积具有调节作用。
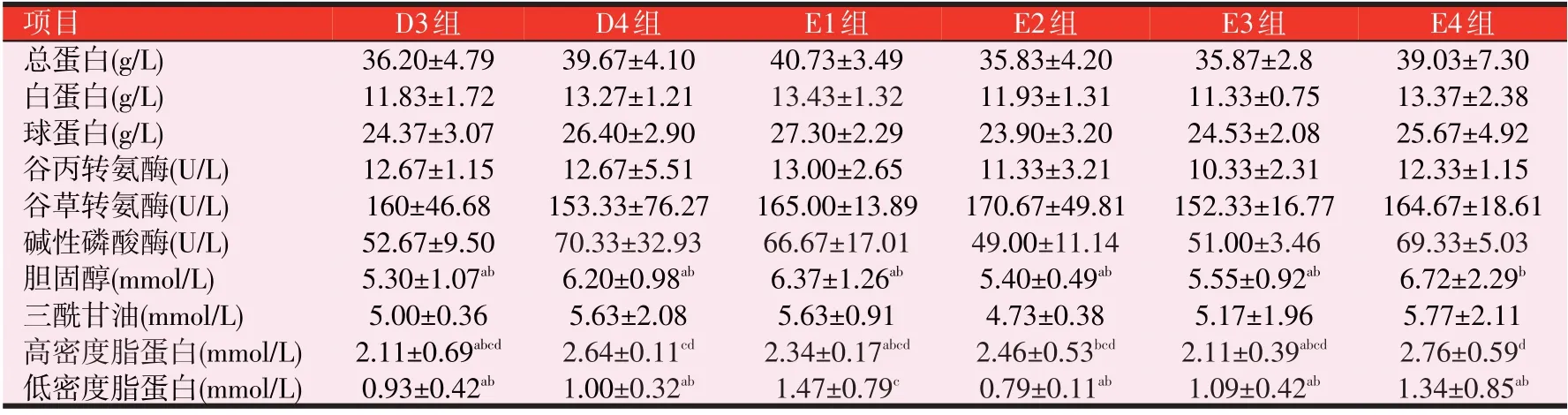
表5(续) 3种诱食剂对斑点叉尾鮰的血清生化指标的影响

表6(续) 3种诱食剂对斑点叉尾鮰抗氧化能力的影响
3.2 3种诱食剂对斑点叉尾鮰血清生化指标的影响
动物机体的生理状况可以通过血清中各项生化指标来反映,血清TP 含量能反映水产动物的营养状况和免疫应激状态,一旦受到胁迫和病害等应激因素均可以引起鱼类血清TP含量下降。饲料中TMAO添加量为0.2%、DMPT 添加量为0.04%和ESP 添加量为0.3%的斑点叉尾鮰血清TP 分别升高了10.34%、18.52%和21.69%。3 种诱食剂均不同程度提高了斑点叉尾鮰TP含量,说明这3种诱食剂具有促进斑点叉尾鮰机体的免疫能力。ALT 和AST 是线粒体中重要的氨基酸转氨酶,当二者含量升高时蛋白质代谢就会增强,当肝脏发生病变时也会引起血液中的ALT 和AST 的活性升高。在本试验中诱食剂对斑点叉尾鮰血清的ALT和AST活性影响不显著,因此蛋白质的代谢速率变化较小,也可能是造成血清TP 差异不显著的一个原因,同时表明3种诱食剂对斑点叉尾鮰的肝脏没有负面影响。ALP 作为能够反映动物机体肝脏功能损伤情况,肝脏受到损伤ALP活性便会上升。饲料中添加3种诱食剂ALP没有显著差异,表明对斑点叉尾鮰的肝脏没有负面影响。饲料中添加外源性诱食剂能升高斑点叉尾鮰血清CHOL和TG含量。胡俊茹等在凡纳滨对虾的日粮中添加外源性诱食剂增加了血清CHOL 和TG 含量,且肝脏机能状态良好。TG含量的升高是体内能量增加的原因,水产动物的细胞分裂速率加快,细胞膜和细胞器发育也需要三酰甘油提供必要的脂类,从而促进水产动物生长。CHOL 是作为皮质类激素和性激素的一种前体物质,还参与细胞膜和细胞器膜的合成等。各试验组添加诱食剂后血清中CHOL 和TG 均出现升高的趋势,这说明诱食剂通过调节斑点叉尾鮰的摄食,影响鱼体的生理生化反应,加快新陈代谢,促进生长。HDL 是将肝外的CHOL 运输到肝脏内的运载工具,LDL 是将肝脏内的CHOL 运输到肝外的运载工具。本试验中,在饲料中添加0.3%的ESP 显著升高了LDL 的含量,添加量为0.5%、0.9%时HDL 含量显著升高;在饲料中添加0.04%的DMPT也显著升高了HDL含量。
3.3 3种诱食剂对斑点叉尾鮰抗氧化能力影响
T-AOC 能有效清除内源性过量ROS,作为机体抗氧化系统的重要组成部分,对维持机体氧化和抗氧化的动态平衡状态具有重要意义。本试验中,饲料中添加3 种诱食剂对斑点叉尾鮰血清中的T-AOC 均有不同程度的提升。饲料中TMAO的添加量为0.15%和0.2%时,斑点叉尾鮰血清中T-AOC均提高了32.61%,TMAO具有一定的抗氧化特性,缺氧条件下可作为一种氧供体。饲料中DMPT 的添加量为0.02%和0.04%的T-AOC 分别提高了17.39%、26.09%。随ESP 的添加量的增加而呈现上升趋势,E 组提高了10.87%~84.78%。SOD 作为一种抗氧化酶可以清除生物氧化过程中产生的超氧阴离子自由基,在清除超氧阴离子自由基的同时生成的过氧化氢(H2O2),H2O2可被CAT清除生成水和氧气。MDA 是脂肪酸过氧化产物之一,其含量的多寡可反映机体内脂质过氧化的程度。当ESP 添加量达到0.07%和0.09%时会显著降低SOD的活性,因此添加量不宜超过0.07%,否则会损害斑点叉尾鮰的肝脏健康。TMAO 的添加水平在0.05%、0.10%、0.15%和0.20%是可以显著降低MDA 含量,DMPT 的添加水平在0.01%、0.02%和0.04%时显著降低了MDA 含量,ESP 在添加量为0.5%时降低了MDA的含量。饲料中添加鱼溶浆、酵母膏和复合诱食剂均显著降低了MDA 的含量,此结果与本试验结果一致。本试验结果显示,饲料中添加外源性诱食剂能提高斑点叉尾鮰血清中T-AOC 和SOD 活性,显著降低MDA 含量,表明诱食剂可以显著改善机体的抗氧化能力。
4 结论

