糖料作物抗氧化酶基因应答非生物胁迫的研究进展
2021-10-17李金娜李海英
李金娜,李海英
(1.黑龙江大学农业微生物技术教育部工程研究中心,哈尔滨 150500;2.黑龙江大学生命科学学院/黑龙江省普通高校分子生物学重点实验室,哈尔滨 150080)
0 引言
糖料作物主要有甘蔗、甜菜、甜高粱和甜叶菊等。在我国北方糖料作物以甜菜为主,南方以甘蔗为主[1-2]。甘蔗是重要的糖料作物,我国食糖总产量90%左右来自甘蔗,近几年甜菜糖产量有增加趋势,2019/2020 年榨季以来,甜菜糖产量占全国食糖产量的13%以上[3]。甜叶菊生产的甜菊糖苷,因甜度较高且具有一定的保健作用,作为糖料作物的补充,在我国的南北方均有种植[4]。甜高粱还原糖和淀粉含量较高,目前多用于能源作物的开发应用,因其生长速度快、产量高、适应性强也被广泛种植[5]。糖料作物是制糖工业的原料作物,也是生物能源的良好原料。糖料作物的一些次生代谢产物如甜菜素等可以作为食品添加剂和化妆品的着色剂,因此糖料作物是地方发展特色工业,拉动区域经济增长的优势作物。由于不同的地理位置和气候条件,糖料作物和其他农作物一样经常会遭受不同逆境胁迫(生物胁迫和非生物胁迫),非生物胁迫通常作为植物减产的主要因素而备受研究人员的广泛关注,全球约5%的土地受到盐渍化的影响[6];每年非生物胁迫造成植物6%~20%的损失[7]。植物遭受非生物胁迫后,体内的活性氧(ROS)产生与清除处于不平衡状态,加剧了植物体内活性氧积累,从而对植物造成不同程度的伤害,严重的甚至会引起植物萎蔫死亡。为了稳定这种平衡,植物体内形成有效的活性氧(ROS)清除机制,包括酶促抗氧化系统和非酶促抗氧化系统两种[8-9],其中酶促抗氧化系统是清除活性氧的主要途径,酶促抗氧化系统中超氧化物歧化酶(SOD)、抗坏血酸过氧化物酶(APX)等活性的高低是衡量植物是否抵抗非生物胁迫的重要指标[10]。近年来,全球变暖、水资源短缺越发突显,致使非生物胁迫加剧[7],因此了解和掌握糖料作物抗氧化酶相关基因应答非生物胁迫对我国糖料作物的优质高产、抗逆品种的培育具有重要的理论指导意义。
1 非生物胁迫诱发植物活性氧的积累与清除途径
1.1 非生物胁迫
非生物胁迫主要包括盐碱、干旱、热、冷等胁迫。非生物胁迫严重危害植物生长及生存。越来越多的研究表明非生物胁迫对植物造成伤害的重要原因之一是使植物体内活性氧(ROS)的产量增加,清除能力降低,使植物体内活性氧(ROS)的产生与清除平衡遭到破坏,导致植物体代谢紊乱、衰老甚至死亡。非生物胁迫造成的初级胁迫主要为渗透胁迫和离子胁迫,渗透胁迫使质膜通透性增加,致使细胞内离子和有机质外渗,离子胁迫减少植物对钾离子的吸收,使植物容易缺乏PO43-和Ca2+。由初级胁迫引发的次级胁迫主要为氧化胁迫,氧化胁迫导致膜脂过氧化程度升高,膜的结构与功能改变[10]。
1.2 植物在非生物胁迫下活性氧(ROS)的积累途径
越来越多的研究表明,非生物胁迫对植物造成的损害最终都会产生过多的活性氧(ROS)积累,非生物胁迫诱发活性氧(ROS)积累包括以下4 种途径[10-13]:(1)叶绿体光电子传递链中活性氧(ROS)的积累;(2)线粒体呼吸链电子的漏出引发活性氧(ROS)的积累;(3)过氧化物酶体中光呼吸途径乙醇酸氧化生成乙醛酸引发活性氧(ROS)的积累;(4)细胞其他部位代谢过程活性氧(ROS)的积累,例如质膜中NADPH 氧化酶可产生活性氧(ROS),因此植物在不同非生物胁迫下活性氧(ROS)积累途径也不尽相同。
盐胁迫产生细胞活性氧(ROS)途径比较广泛,如光合电子传递、光呼吸过氧化氢(H2O2)释放、呼吸电子传递和酶活性。酶类包括葡萄糖氧化酶、黄嘌呤氧化酶,特别是植物过氧化物酶(POD)和NADPH 氧化酶。最重要的是质膜结合NADPH氧化酶(RBOH),它决定性地控制盐胁迫下细胞氧化还原稳态[14]。
干旱胁迫产生细胞活性氧(ROS)途径主要包括线粒体呼吸链电子的漏出引发活性氧(ROS)的积累,叶绿体光电子传递链中活性氧(ROS)的积累和过氧化物酶体中光呼吸途径乙醇酸氧化生成乙醛酸引发活性氧(ROS)的积累。在大田环境中,干旱胁迫经常与高温胁迫协同发生,也是实际生产中最常见的一种双重胁迫,当植物遭受干旱胁迫时,植物体内电子供需不平衡,在线粒体、叶绿体、过氧化物酶体、内质网、质膜和细胞壁中会产生大量活性氧(ROS),引起氧化胁迫[15]。氧化胁迫会破坏许多膜蛋白和酶的空间构型,使膜透性增加和离子泄漏,影响植物正常的生理生化功能,严重时会引起植物死亡[16-17]。高温胁迫造成活性氧(ROS)积累的途径主要是膜损伤和脂膜透性增加[18]。
低温胁迫产生细胞活性氧(ROS)途径主要包括质膜中NADPH 氧化酶可产生ROS和叶绿体光电子传递链中ROS 的积累,低温胁迫下植物脂质过氧化物丙二醛(MDA)大量积累,会使膜系统严重受损;同时ROS的产生与PSⅠ的光抑制密切相关,低温胁迫引起PSⅠ产生过氧化物和单线态氧,从而引起PSⅠ的光抑制伤害,因此活性氧(ROS)的产生与PSⅠ的光抑制密切相关[18-21]。
1.3 植物在非生物胁迫下活性氧(ROS)的清除途径
为了应对非生物胁迫所引发的氧化胁迫,植物在长期进化过程中形成了完善的抗氧化系统(图1)[17],抗氧化系统可以通过酶促抗氧化系统和非酶促抗氧化系统共同调节植物体内ROS水平,从而防止ROS引起的脂膜过氧化及其他伤害。酶促抗氧化系统主要包括超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)、过氧化物酶(POD)等13 种酶;非酶促抗氧化系统主要包括抗坏血酸(AsA)、还原型谷胱甘肽(GSH)、类胡萝卜素(Car)和生育酚(VE)等物质(图1A)[22]。酶促抗氧化系统为植物提供高效而专一的ROS清除体系(图1B)[23-25],因此对抗氧化酶基因应答非生物胁迫的研究具有重要意义。

图1 植物抗氧化系统(A)、酶促抗氧化酶系统和非酶促抗氧化系统途径(B)Fig.1 The antioxidant system(A)and the pathways of enzymatic antioxidant enzyme system,and non-enzymatic antioxidant system(B)in plant
2 非生物胁迫下糖料作物抗氧化酶相关基因的研究进展
截止到目前,国内外研究人员对糖料作物如甘蔗、甜菜、甜高粱和甜叶菊的抗氧化酶相关基因应答非生物胁迫下的研究主要集中抗氧化酶活性分析及表达模式分析等方面,而对其功能验证研究较少。
2.1 非生物胁迫下甘蔗抗氧化酶相关基因响应研究进展
在甘蔗中,目前研究人员对干旱和低温非生物胁迫下的抗氧化酶相关基因应答进行了研究(表1)。王盛[26]对甘蔗中铜/锌超氧化物歧化酶基因(Cu/Zn-SOD)进行克隆并进行了低温、盐、干旱和H2O2四种非生物胁迫应答模式分析,研究发现4 种非生物胁迫均诱导Cu/Zn-SOD基因的表达,同时将甘蔗Cu/Zn-SOD基因转入烟草中,但遗憾的是没有对Cu/Zn-SOD基因转基因植物应答非生物胁迫进行功能验证,不过为以后研究Cu/Zn-SOD基因与非生物胁迫的关系及甘蔗抗逆育种奠定了基础。
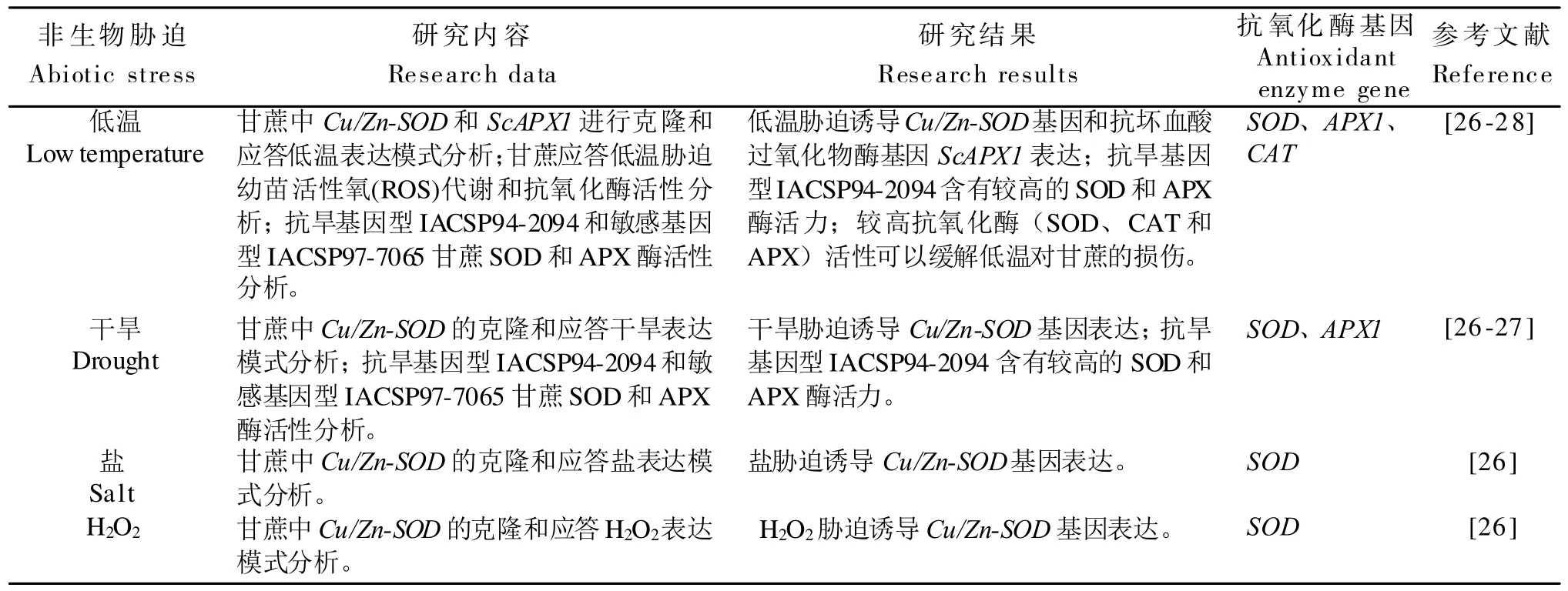
表1 甘蔗抗氧化酶基因应答非生物胁迫主要研究内容Table 1 Main research data of antioxidant enzyme system’s genes responseto abiotic stress in sugarcane
SALES[27]等人研究了超氧化物歧化酶(SOD)和抗坏血酸过氧化物酶(APX)可提高干旱和低温条件下甘蔗植株光合作用的恢复能力,通过对抗旱基因型‘IACSP94-2094’和敏感基因型‘IACSP97-7065’甘蔗进行光合及抗氧化酶活性等生理生化指标测定,结果表明,抗旱基因型‘IACSP94-2094’含有较高的SOD 和APX 酶活力,‘IACSP94-2094’抗氧化能力较强。同时‘IACSP94-2094’在干旱和低温条件下可以单独或协同维持植物生长。研究结果显示SOD 和APX 高活性与甘蔗植株在同时发生干旱和低温的严重胁迫后光合作用的完全恢复有关。
张保青[28]等人对甘蔗抗坏血酸过氧化物酶基因ScAPX1进行克隆和表达分析,研究发现随着低温(0~4℃)胁迫时间的延长,2个甘蔗基因型ScAPX1的表达量均是先上升后下降,但表达量存在差异,在整个胁迫过程中,抗寒性强基因型‘GT28’的表达量始终比抗寒性弱基因型‘YL6’高。甘蔗ScAPX1积极响应低温逆境胁迫,该基因的诱导表达与甘蔗基因型本身的抗寒性密切相关。该研究为深入研究甘蔗APX1在低温胁迫中的功能及甘蔗抗寒育种的分子机理提供依据。
2.2 非生物胁迫下甜菜抗氧化酶相关基因响应研究进展
很多研究人员对甜菜抗氧化酶相关基因进行系统性的研究,不同基因型甜菜应答盐、干旱、水分等非生物胁迫的抗氧化酶相关基因进行功能验证以及对抗氧化酶系统相关基因之间潜在的联系进行验证及分析(表2),为甜菜抗氧化酶系统相关基因响应非生物胁迫中的功能及甜菜抗非生物胁迫的分子机理提供依据。

表2 甜菜抗氧化酶基因应答非生物胁迫主要研究内容Table 2 Main research data of antioxidant enzyme system’s genes response to abiotic stress in sugar beet
温丽[29]通过模拟水分胁迫条件,测定分析了不同抗旱基因型甜菜体内活性氧(ROS)代谢系统的动态变化和重要保护酶基因在转录水平上的表达,结果表明抗旱性强的甜菜基因型体内SOD基因表达量大于抗旱性弱的基因型,但SOD基因的表达量与水分胁迫强度之间不存在相关关系;POD基因表达量多少与POD 酶活性变化呈现正相关关系。该研究从活性氧(ROS)代谢系统的动态变化及重要保护酶基因在转录水平方面阐明了甜菜抗旱生理生化机制,为甜菜抗旱生理基础研究及选育抗旱新品种提供理论依据。
赵晨曦[30]对甜菜‘M14’品系硫氧还蛋白过氧化物酶(Tpx)应答盐胁迫及氧化胁迫基因功能进行研究,通过构建BvM14-Tpx基因的酵母菌表达体系,测定转BvM14-Tpx基因的酵母菌体的生长曲线,验证BvM14-Tpx基因能够缓解H2O2和NaCl对大肠杆菌生长的抑制。同时该研究将BvM14-Tpx基因通过转基因手段转入到拟南芥突变株中,结果表明转BvM14-Tpx基因的拟南芥突变株能够使其具有与野生型拟南芥植株对盐及氧化胁迫的抵御能力。
蒋德生[31-34]等人研究甜菜‘M14’品系叶片中参与胁迫和防御的活性氧(ROS)清除机制中抗氧化酶系统7 种主要酶乙醇酸氧化酶(GOX)、过氧还蛋白(PrxR)、硫氧还蛋白(Trx)、抗坏血酸过氧化物酶(APX)、脱氢抗坏血酸还原酶(DHAR)、单脱氢抗坏血酸还原酶(MDHAR)及过氧化氢酶(CAT)。发现在盐胁迫条件下,BvM14-GOX、BvM14-PrxR、BvM14-Trx、BvM14-DHAR、BvM14-MDHAR、BvM14-APX及BvM14-CAT基因在甜菜‘M14’品系中应答盐胁迫诱导表达量更高,因此甜菜‘M14’品系具备更强的抗氧化酶活性。为进一步探究抗氧化酶系统相关基因的调控作用,将抗氧化酶系统关键基因进行耐盐功能的鉴定及评价,确定了甜菜耐盐抗氧化酶的关键基因及其抗氧化协同调控网络,最终阐明了甜菜‘M14’品系高效应答盐胁迫的抗氧化酶调控机制。
HOSSAIN[35]等人研究了甜菜耐盐驯化中的氧化还原活性氧途径,在转录水平上研究了氧化还原和活性氧(ROS)网络的调控。首先在甜菜基因组中鉴定了超氧化物歧化酶(SOD)、过氧化物氧化蛋白(Prx)、交替氧化酶(AOX)、质体末端氧化酶(PTOX)和NADPH 氧化酶(RBOH)的基因家族。盐胁迫可以诱导Cu/Zn-SOD、Mn-SOD、Fe-SOD3、所有AOX 亚型、2-Cys-PrxB、PrxQ和PrxIIF基因的积累。而Fe-SOD1、1-Cys-Prx、PrxIIB和PrxIIE基因表达量则随着盐度的增加而降低。为了深入了解抗氧化系统相关基因的调控,研究人员分析了甜菜抗氧化酶相关基因的启动子顺式元件。该结果表明,甜菜抗氧化酶基因启动子中特殊的顺式元件参与调节氧化还原和活性氧(ROS)调控网络,是提高甜菜耐高盐性的基础。
2.3 非生物胁迫下甜高粱抗氧化酶相关基因响应研究进展
糖料作物甜高粱中目前只有一篇硕士论文对抗氧化酶系统进行系统性分析。李俊付[36]研究甜高粱在不同水分胁迫下及不同发育阶段对水分亏缺所产生的适应性反应,重点进行了水分胁迫下甜高粱抗氧化酶的活性分析测定,同时对主要抗氧化酶基因进行了定量表达模式分析,结果表明,不同生育时期超氧化物歧化酶(SOD)、过氧化物酶(POD)基因的表达量不同,不同水分胁迫处理下甜高粱POD基因表达量无显著差异;SOD基因的表达差异比较明显。说明抗氧化酶系统参与了甜高粱水分胁迫响应,对甜高粱在干旱胁迫应答具有潜在的调节保护作用。该研究旨在找出甜高粱在非生物胁迫下的反馈调节机制。
2.4 非生物胁迫下甜叶菊抗氧化酶相关基因响应研究进展
在糖料作物甜叶菊中,研究人员对抗氧化酶相关基因应答盐、干旱等非生物胁迫的表达模式和酶活性进行分析。绳仁立[37]对5 个甜叶菊品种耐盐碱进行研究,通过结合膜系统、渗透调节物质、抗氧化酶活性等对其耐盐机理进行测定分析。该研究发现抗氧化酶系统酶活性较高的基因型具有较强的耐盐碱性,为甜叶菊在抗旱、抗寒、抗盐等领域的育种研究奠定基础。
CANTABELLA[38]等人从矿质营养、抗氧化代谢和甜菊糖苷含量的影响对甜叶菊的耐盐机制进行探讨,研究发现一个有趣的现象,在短期盐胁迫条件下,甜叶菊可通过诱导不同的抗氧化机制来严格控制活性氧(ROS)的生成。例如,在第16 d 观察到抗氧化酶(APX、POX、CAT、SOD)以及AsA-GSH 循环的其他酶显著增加,表明甜叶菊植株在这个阶段具有平衡的抗氧化能力;但是在长期盐胁迫条件下,AsA-GSH 循环酶发生失衡,主要表现为APX、DHAR和GR 的酶活性下降,而MDHAR的酶活性显著增加。该研究结果表明,甜叶菊能够诱导短时间(16 d)盐胁迫或维持长时间(25 d)盐胁迫的抗氧化酶来应对盐胁迫带来的损伤。
3 总结与展望
糖料作物应答非生物胁迫条件下积累过量的活性氧(ROS),因此,植物体对活性氧(ROS)的清除是必不可少的。本文通过综述国内外近10 年对不同糖料作物抗氧化酶相关基因应答在非生物胁迫酶活性和应答非生物胁迫表达模式的研究,发现研究集中在超氧化物歧化酶(SOD)基因和抗坏血酸过氧化物酶(APX)基因,因为SOD 是活性氧(ROS)清除途径中的第一个关键酶,APX 是该途径的第二个重要酶[39],因此近年来很多研究人员对编码抗氧化酶SOD基因和APX基因提高糖料作物抗非生物胁迫做了大量研究。
在整个抗氧化系统中,关于抗氧化酶编码基因SOD、APX、CAT、MDHAR、GR等应答非生物胁迫表达模式及其对应酶活性有较多研究,然而,对抗氧化酶编码基因还有愈创木酚过氧化物酶(GPX)基因、多酚氧化酶(PPO)等基因功能没有深入研究。
其次,抗氧化酶基因之间相互作用是否能够提高植物的抗非生物胁迫值得进一步研究和探讨。
最后,活性氧(ROS)清除系统不仅仅通过酶促抗氧化系统单一因素来缓解非生物胁迫带来的损害,还需要与非酶促抗氧化系统如抗坏血酸(AsA)基因和还原型谷光甘肽(GSH)基因协同调节植物体内活性氧(ROS)水平,从而防止活性氧(ROS)引起的脂膜过氧化及其他伤害。
因此,糖料作物未来的研究更应该将酶促抗氧化系统基因和非酶促抗氧化系统基因在逆境胁迫下同步分析,为糖料作物抗逆生理基础研究及选育抗逆新品种提供理论依据。
