多层FitzHugh-Nagumo 神经元网络的控制同步
2021-10-13程卉
程 卉
(武汉大学数学与统计学院, 湖北 武汉430072)
1 引言
复杂网络是生物、物理、工程和社会科学中很多系统的通用模型[1-3]. 近些年来, 人们开始对网络中的代数关系、统计内容和图论相关性质以及它们和网络的动力学之间的关系产生浓厚的兴趣. 随着神经网络在信息传输、模式识别和图像处理等方向的广泛应用, 神经系统的动态行为成为研究的热点[4-6].
生物神经系统是一个由数亿个神经细胞构成的极其错综复杂的信息网络. 生物机体的整个系统和器官在信息的加工和传播过程中受到控制和协调, 使其能够适应内外环境的差异,完成各种生理过程[7]. 神经元作为神经系统中最基本的结构单元, 其许多的动态行为通过放电活动来表现出来. 许多个神经元通过电突触和化学突触紧密相连, 组成一个巨大的复杂的神经元网络, 具有高层次和多功能的特性. 为了能更加了解神经元信息传递的过程以及其认知、思想、控制等功能, 我们需要探索更多的神经元和网络的动力学行为.
过去生物神经研究主要集中在大脑的不同部分, 但现代的研究更偏向于使用复杂网络的模式来分析不同神经元网络的结构和动力学行为[8]. 神经元网络中最基本的结构单元是神经元, 而突触建立了神经元之间进行信息交流和传递的桥梁. 由于神经元的类型和神经元间连接的突触方式的多样性, 神经系统的拓扑结构复杂多样, 具有非线性的动力学行为. 庞大的神经系统对于信息的处理并不是由单个的神经元完成的, 而是由相互作用的神经元集群共同完成的. 所以, 我们需要从复杂网络动力学的角度来探究神经元系统进行信息处理、管理、整合的方式, 通过研究神经元系统的复杂同步行为和时变动力学行为来进一步了解神经元的高级认知功能.
同步是非线性系统中最基本的合作行为形式之一, 在自然界和社会现象中经常发生, 同时在神经元系统中也普遍存在[9,10]. 根据许多研究表明, 探究和理解大脑的功能已开始广泛应用混沌同步理论. 在神经科学中, 对于健康和不健康的大脑的功能分析在理解神经元与其网络之间复杂的相互作用机制方面扮演着重要的角色[11]. 但是导致脑部疾病的一个重要原因是神经元间无法同步, 像帕金森氏病, 自闭症等影响身体功能的疾病已被证实与神经元间缺乏同步有关[12,13].
到目前为止, 研究人员们已经提出了很多种控制方案来实现神经元网络的同步, 例如主动控制、自适应控制、反同步控制、滑模控制、有限时间同步控制等. 至今对FitzHugh-Nagumo(简称FHN) 神经元网络在外部施加电刺激的同步控制已经得到了许多有效的研究[14-19]. 文献[14] 利用后推和滑膜控制技术的鲁棒控制实现了两个间隙连接耦合的混沌FHN神经元的同步, 通过仿真证明了控制方案的有效性. 文献[15] 考虑了两个具有不同离子电流的非耦合FHN 神经元与外部电刺激之间的同步, 应用鲁棒自适应滑模控制器解决了系统的不确定性和外部干扰, 在Lyapunov 稳定性定理的意义上给出了保证稳定同步的充分条件.文献[16] 利用空间采样策略的新型控制方法实现了一类反应扩散FHN 系统之间的同步, 通过数值仿真较好地展示出控制器的效率. 现有研究神经元系统同步控制的文献大多考虑单层网络, 但在实际应用中, 神经元之间通过不同的突触连接方式相互联系, 形成多层次的神经元网络, 因此考虑多层神经元网络的同步控制具有重要价值. 本文从数学角度出发, 考虑多层神经元网络, 多层网络中同步的控制条件与单层网络有所不同, 且相互之间有一定的联系, 这些均在文中有详细证明和介绍. 另外神经元节点间通过分量来耦合, 且考虑控制节点的分量对同步的影响, 这对于以前的研究而言是创新部分, 为控制神经元网络的同步提供了一些新的思路.
本文研究了由FHN 神经元模型耦合而成的多层神经元网络, 探究控制神经元节点的不同分量对于系统达到同步的影响. 在单层网络和多层网络两种情况下, 基于舒尔补式[20]和李雅普诺夫稳定性理论[21], 分别给出了保证神经元系统稳定同步的充分条件. 通过比较分析,从理论上讨论了两者间同步条件的差异与联系.
2 数学准备
我们首先介绍一些本文中将要使用的符号.Rn和Rn×m分别表示n维欧几里得空间和所有n×m维实矩阵的集合,R+是正实数集,‖·‖表示向量或矩阵的二范数,λmax(·) 表示对称矩阵的最大特征值,λmin(·) 表示对称矩阵的最小特征值.In×n表示维数为n的单位矩阵.
2.1 数学知识准备



利用上式, 不难验证以下有用结论
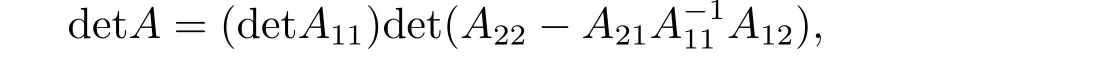
上式就是关于行列式的Schur 补公式, 也称为Schur 公式.
2.2 FitzHugh-Nagumo(FHN) 神经元模型
自1951 年以来, 对电化学细胞的定量研究主要受到了霍奇金和赫胥黎对长乌贼巨型轴索神经传导的杰出研究的推动. 霍奇金和赫胥黎主要是建立四维模型来表示在乌贼的巨大轴突上传播脉冲的过程[22], 该模型能够描述神经元膜内的自激振荡、混沌、多重稳定等非线性的现象, 为人们探索神经元的兴奋性提供了基本框架. 上千个神经元是如何相互连接, 怎么协调有序放电是理论研究和实验探索的重要课题. 这些年来, 复杂网络理论的发展为探索这一问题提供了一个很好的新方法.
由于这些方程太过复杂, 无法完全分析, 为了帮助理解霍奇金- 赫胥黎方程的性质, 更简单的方程组确实是必要的. 因此, 考虑到生理背景, FitzHugh(1961) 和Nagumo(1962) 独立地导出了一个二维系统, 该系统提供了一个四维的霍奇金- 赫克斯利方程的简化[22-25].
Fitzhugh-Nagumo (FHN) 神经元模型可以用下式描述:

其在同步流形s=(s1,s2) 处线性化后可以得到

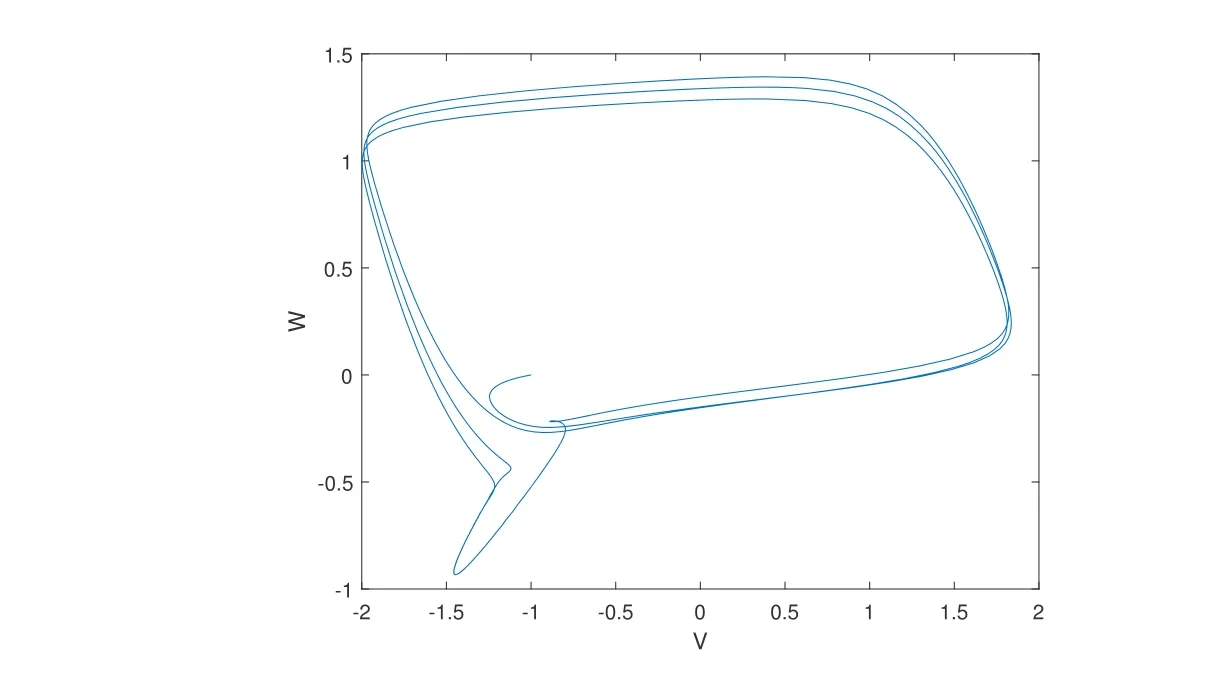
图1:FHN 模型中状态变量W 和V 的相图, 其中 = 0.08,a = 0.7,b = 0.8,Iex = 0.5cos( 5t0).可以看到, 系统的轨道收缩到极限环, 所以变量V 和W 是有界的. V 的边界可以取2.
3 主要结论
3.1 单层网络
考虑一个二维的具有N个神经元的复杂网络. 描述网络动态演化的方程为:


因为本文着重研究多层神经元网络中控制节点分量对于网络动力学行为及同步现象的影响, 所以内联矩阵均考虑取对角元素为0 或1 的对角矩阵. 一方面是因为在一般的多层网络中也会考虑取对角矩阵, 这样在进行研究证明时会更方便, 如文献[26];另一方面, 考虑控制节点分量对于同步的影响时, 内联矩阵取对角矩阵同时也可以考虑节点之间耦合不同分量时同步条件是否有所变化.
当内联矩阵P=diag(1,0) 时, 对于FHN 模型网络, 向量方程(3.1) 可以写成标量形式:
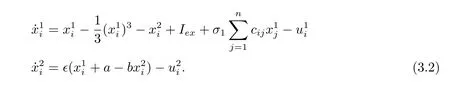
令ei≜xi-s(1≤ i ≤ N), 其中s=(s1,s2)∈R2是同步态, 将网络方程(3.2) 在s上


若换成其他内联矩阵时网络依旧能同步, 大部分神经元网络文章中均有介绍, 但同步条件与本文有所不同, 这不是本文考虑的重点, 所以此处不再赘述.
上面讨论了在单层生物神经元网络的情况下, 当节点仅通过第一分量相互耦合时, 可以对节点的第一分量进行线性控制, 从而实现网络的局部同步. 那么当节点通过第二分量相互耦合时, 考虑是否可以通过控制节点第二的分量来实现网络同步.

那么有误差系统
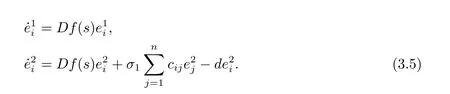

其中s1是同步态s= (s1,s2) 中的第一个分量, 那么神经元网络(3.4) 是局部同步的, 即t →+∞,ei →0.
证 选取系统(3.4) 的李雅普诺夫函数如下:

其中Q1=σ1C-dIn×n-∈bIn×n.
那么有
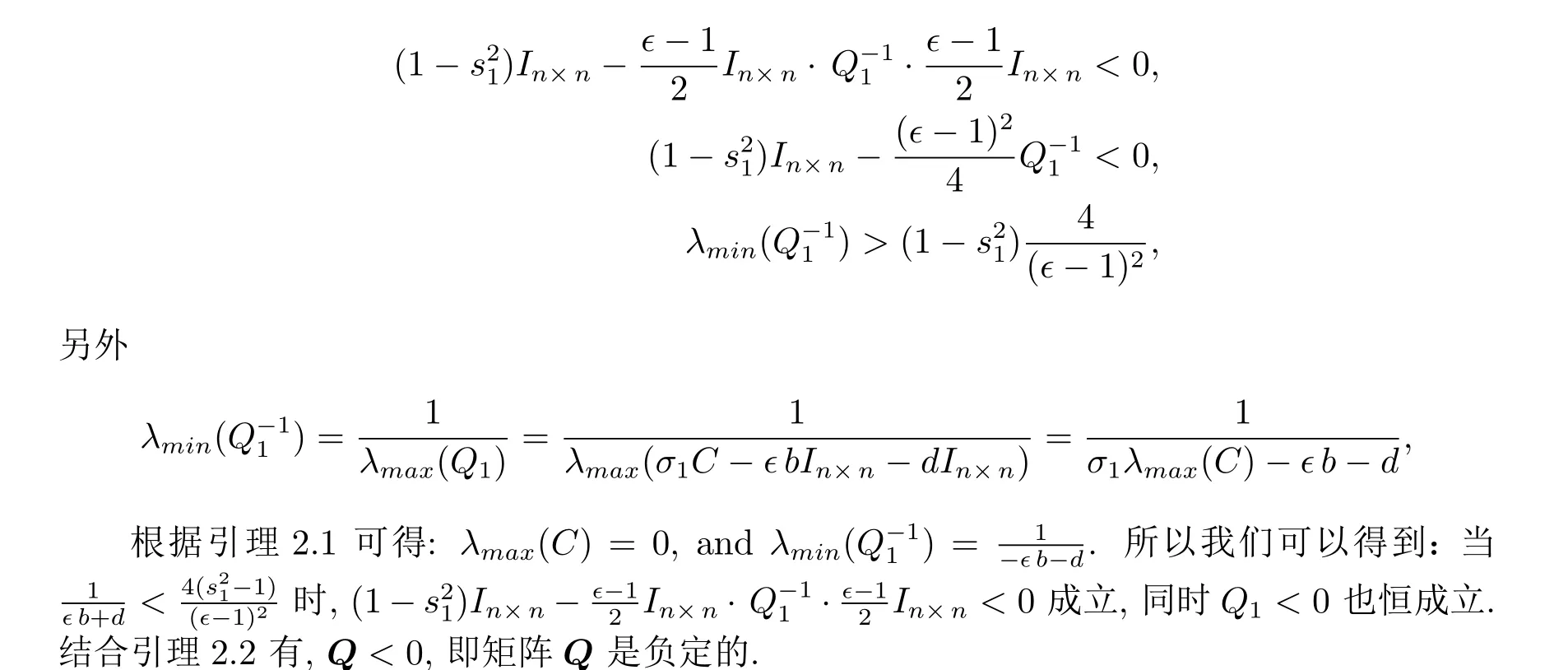
根据LaSalle 不变原理, 系统的每个解收敛到集合的最大不变集˙V(t)=0, 即E1i=0,E2i=0(1≤i ≤n). 所以当t →+∞,ei →0(1≤i ≤n), 网络处于局部同步状态.
注3.3 对于动态神经元网络(3.4), 当节点通过第二个分量相互耦合时, 对节点的第二个分量施加控制, 使网络同步所需的控制强度也受到神经元节点自身动力学的影响, 与耦合强度和耦合矩阵无关. 此时的控制强度与耦合控制第一分量时所需的控制强度有所不同, 且所需的控制强度更大.

3.2 两层网络

网络(3.7) 是局部同步的, 即t →+∞,ei →0.

则K <0. 所以有

同样地有

那么f(x)<0 且F <0.
另一方面,K2=σ2G-d2In×n-∈bIn×n <0 始终成立. 根据引理2.2, 我们可推断出当d1,d2取合适的正实数时,K <0, 也就意味着K是负定的.
根据LaSalle 不变原理, 系统的每个解收敛到集合的最大不变集˙V(t)=0, 即E1i=0,E2i=0(1≤i ≤n). 所以当t →+∞,ei →0(1≤i ≤n), 网络处于局部同步状态.
注3.5 对于两层生物神经元动态网络(3.6), 当对每个节点的两个分量都施加线性控制时, 网络可以达到局部同步, 其所需的控制强度d1,d2相互限制、相互影响, 与节点本身的动力学有关. 它们也与耦合强度、耦合矩阵等无关. 当其中一个分量上的控制强度降为0 时, 另一个分量上所需的控制条件与单层网络情况相同.
定义下式

当g <0 时网络可以达到一个同步状态. 若取∈=0.08,a=0.7,b=0.8, 那么FHN 模型中的神经元节点的分量是有界的. 从图1 中我们可以计算出s1的界可以取值为2, 我们可以画出g关于d1和d2的图像, 如图2 所示.
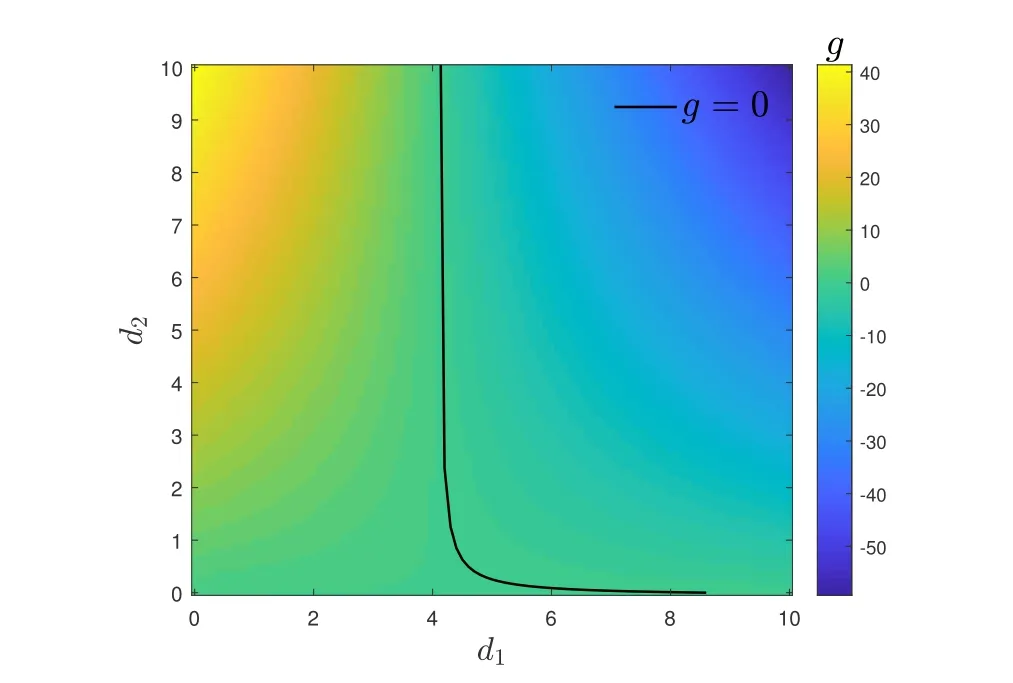
图2: 两层生物神经元网络中分别控制节点的两个分量时, 网络同步与控制强度之间的关系.
由上图2 可知, 当d2=0 时,d1=d, 即与仅控制第一分量时所需的控制强度相同. 同样地, 当d1= 0 时,d=d, 即与仅控制第二分量时所需的控制强度相同. 当d2>0,d1<d, 这说明在对每个节点的第二个分量施加控制后, 网络达到同步时对第一个分量所需的控制强度会相应降低. 相应地, 当d1越小, 所需的d2会变得更大. 这相应地说明, 两者之间是相互影响, 共同促进网络同步的关系.
4 结论
综上所述, 我们讨论了由FHN 模型耦合而成的多层神经元网络, 探究控制神经元节点的不同分量对于系统达到同步的影响. 对单层网络和两层网络进行了分类和讨论, 通过严格的理论验证, 分别给出了保证神经元系统稳定同步的充分条件.

我们可以发现, 由FHN 模型耦合的单层生物神经元网络和多层生物神经元网络中实现同步时所需的控制强度是不同的. 当多层网络中仅控制一个分量时, 所需的控制强度与单层网络中所需的控制强度相同, 且控制第一分量对于同步的影响要优于第二分量. 多层网络中两个分量上的控制相互影响, 共同促进网络的同步, 另外同步所需的充分条件是单层网络情况的扩展.
未来的工作包括研究由其他模型耦合的网络中控制节点分量对同步的影响, 以及通过牵制控制部分节点, 分析控制节点中某一分量对同步的影响.
