Stem cell therapies in cardiac diseases: Current status and future possibilities
2021-10-11TaisHanaeKasaiBrunswickAdrianaBastosCarvalhoAntonioCarlosCamposdeCarvalho
Tais Hanae Kasai-Brunswick, Adriana Bastos Carvalho, Antonio Carlos Campos de Carvalho
Tais Hanae Kasai-Brunswick, Antonio Carlos Campos de Carvalho, National Center of Structural Biology and Bioimaging, Federal University of Rio de Janeiro, Rio de Janeiro 21941-902, RJ, Brazil
Tais Hanae Kasai-Brunswick, Adriana Bastos Carvalho, Antonio Carlos Campos de Carvalho, National Institute of Science and Technology in Regenerative Medicine, Federal University of Rio de Janeiro, Rio de Janeiro 21941-902, RJ, Brazil
Adriana Bastos Carvalho, Antonio Carlos Campos de Carvalho, Carlos Chagas Filho Institute of Biophysics, Federal University of Rio de Janeiro, Rio de Janeiro 21941-902, RJ, Brazil
Abstract Cardiovascular diseases represent the world’s leading cause of death. In this heterogeneous group of diseases, ischemic cardiomyopathies are the most devastating and prevalent, estimated to cause 17.9 million deaths per year. Despite all biomedical efforts, there are no effective treatments that can replace the myocytes lost during an ischemic event or progression of the disease to heart failure. In this context, cell therapy is an emerging therapeutic alternative to treat cardiovascular diseases by cell administration, aimed at cardiac regeneration and repair. In this review, we will cover more than 30 years of cell therapy in cardiology, presenting the main milestones and drawbacks in the field and signaling future challenges and perspectives. The outcomes of cardiac cell therapies are discussed in three distinct aspects: The search for remuscularization by replacement of lost cells by exogenous adult cells, the endogenous stem cell era, which pursued the isolation of a progenitor with the ability to induce heart repair, and the utilization of pluripotent stem cells as a rich and reliable source of cardiomyocytes. Acellular therapies using cell derivatives, such as microvesicles and exosomes, are presented as a promising cell-free therapeutic alternative.
Key Words: Stem cell; Cell therapy; Cardiac stem cell; Cardiovascular diseases; Progenitor cardiac cells; Pluripotent stem cells
INTRODUCTION
Cardiovascular diseases represent the world’s leading cause of death, and in this heterogeneous group of diseases, ischemic cardiomyopathies are the most prevalent, accounting for 17.9 million deaths per year[1]. Coronary artery occlusion or reduction of blood flow results in irreversible death of cardiac cells with consequent functional cardiac impairment. Despite improvements in the clinical-surgical management of these patients, the main effect of ischemic heart disease — the death of cardiomyocytes — is not reversed. Acute interventions can restore blood flow, avoiding the death of more cardiac cells. However, this neither contributes to the recovery of the function of the damaged tissue nor stops the progression of ischemic disease[2]. Due to the limited cardiac regenerative capacity, the lost cardiomyocytes are replaced by fibrotic scarring, leading to cardiac remodeling and heart failure. The therapeutic option for heart failure patients is an organ transplant, but the demand is far greater than the availability of such organs[3]. This represents a significant public health problem, pushing researchers to look for effective alternative treatments.
In the 1990s, the idea to replace the cardiomyocytes lost due to ischemic diseases with an external cell source paved the way for cell therapies for regenerative medicine in cardiology[4]. In this review, we will discuss the various cell therapies that were applied to treat cardiac diseases, the questionable existence of an endogenous cardiac stem cell (CSC) and their putative cardiac regenerative potential, and, finally, what we envisage as the future in the field which is using cardiomyocytes derived from pluripotent stem cells or their derivatives, such as exosomes and microvesicles (Figure 1).
THE FIRST STEPS OF CARDIAC CELL THERAPY: SKELETAL MYOBLASTS
Skeletal myoblasts (SM) were the first cell type used to treat ischemic cardiac diseases. This was a natural choice as SM were contractile cells with autologous availability, simple isolation, highin vitroproliferation, and resistance to ischemia[5-7].
Preclinical studies showed that SM could survive and engraft after cell transplantation into ischemic hearts and differentiate into myotubules[4,7-10]. Although electromechanical coupling was not observed between the SM and resident cardiomyocytes, generating arrhythmogenic foci, an improvement in cardiac function was observed[11-13].
Phase I and II clinical trials failed to demonstrate the functional benefits observed in experimental studies, and the presence of arrhythmias was also observed in some patients[14]. These unfavorable outcomes stimulated the search for other cell types for cardiac cell therapy.
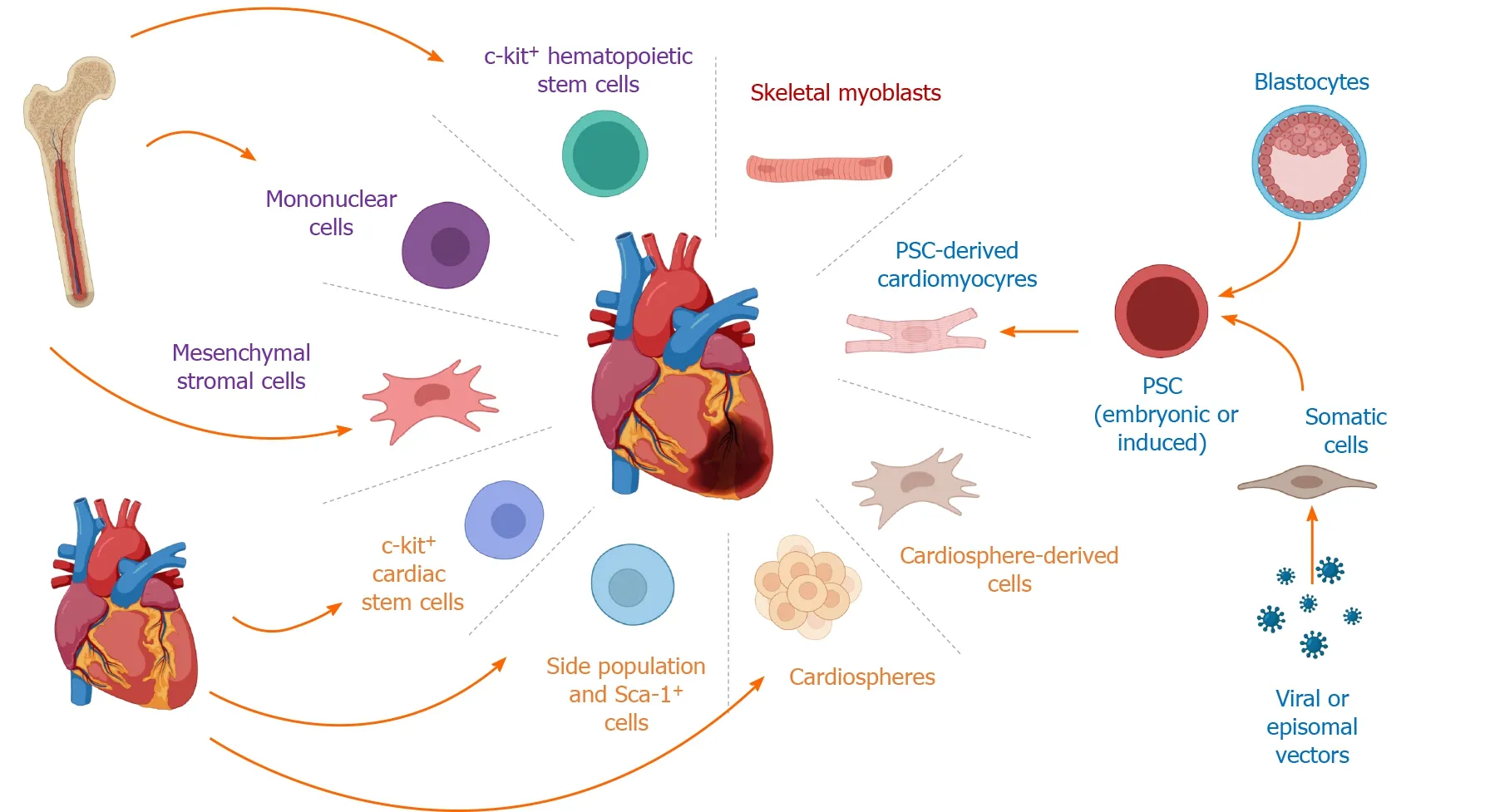
Figure 1 Cell-based therapies for ischemic heart disease.
THE NEXT STEP: BONE MARROW-DERIVED CELLS
Orlicet al[15], showed for the first time, that mouse c-kit+progenitor bone marrow cells could transdifferentiate into cardiomyocytes and improve cardiac function in ischemic hearts. The group injected c-kit+bone marrow-derived cells from a male GFP (green fluorescent protein) mouse into a wild-type infarcted female, claiming that the marrow-derived GFP cells expressed sarcomeric, endothelial, and smooth muscle proteins[15]. The same group achieved similar results by administering stem cell factor and granulocyte-colony stimulating factor into infarcted mice, showing bone marrowderived cell homing into the infarcted area, differentiation into cardiomyocytes, and increased cardiac function[16].
The capability of mononuclear bone marrow cells to transdifferentiate into cardiomyocytes was disputed by independent groups, which showed that, after cell transplantation, these cells could only differentiate into mature blood cells[17,18]. In addition, some authors attributed the improvement in cardiac function after mononuclear bone marrow cell transplantation into ischemic hearts to a fusion process between bone marrow cells and cardiomyocytes[19,20]. However, this fusion mechanism was refuted since it was an extremely rare event that could not account for the improvement in function observed after bone marrow-derived cell therapy[21]. The currently accepted mechanism — the paracrine effect — consists of bone marrow cells releasing factors that could stimulate cellular proliferation, angiogenesis, modulate the inflammatory response, and promote ischemic tissue protection[22].
Equally important to understanding the mechanism involved in cardiac repair was the search for the subset of bone marrow cells that could be more effective. The potential for cardiac repair of mesenchymal stromal cells (MSC), a subpopulation of mononuclear bone marrow cells, was starting to be investigated. Initially, MSC transdifferentiation into cardiomyocytes was also proposed, but these data were also disputed, and the benefits of MSC treatments were also attributed to paracrine effects[22-26].
The preclinical results with bone marrow-derived cells drove several clinical trials to pursue the same positive results in the clinical setting. The clinical trials conducted to treat heart diseases using bone marrow cells were safe and feasible, but the results were controversial regarding the functional benefits. A large number of diverse clinical studies of different designs made the comparison of results difficult. They differed on time to intervention and severity of the treated disease, subset of cells used, number of cells administered, injection pathway, endpoints, functional analyses methods, number of subjects, randomization, blinding, single or multicenter trials, among others. Due to this heterogeneity, even metanalyses were not able to draw definitive conclusions[27,28]. The only common aspect of these trials was that none could reproduce the highly significant preclinical improvements seen on cardiac function, nor could they demonstrate new cardiomyocyte generation. Therefore, scientists continued with their search for the ideal cell type to treat ischemic heart disease.
HEART REGENERATION
The regenerative capacity of the heart has been the subject of intense and controversial investigations for 150 years. However, the paradigm that the heart is a post-mitotic organ and therefore incapable of regenerating itself was challenged in 2009 when Bergmannet al[29] proved that the heart could regenerate itself during an individual’s lifespan. Many nuclear tests were performed during the Cold War, leading to an increase in atmospheric Carbon 14 (C14) concentration. The C14was absorbed by plants and entered our food chain, marking the DNA of dividing cells. After the interruption of the nuclear tests in 1963, the atmospheric concentration of this isotope decreased dramatically. Bergmann’s group compared the concentration of C14present in cardiomyocyte DNA to the atmospheric C14in the year that the individual was born, thus identifying the “age” of the cardiomyocytes. Using this elegant strategy, they showed that there was indeed cardiac cell renovation after birth and, by mathematical modeling, demonstrated that 50% of cardiomyocytes were renewed during the entire lifespan and that this renovation rate was age-dependent (1% per year at age 25, decreasing to 0.45% at age 75)[29].
This paradigm shift led researchers to pose a new question: what cell type was responsible for this cardiac regeneration capability? Hsiehet al[30] addressed this question using a double transgenic MerCreMer-ZEG mouse with the myosin heavy chain 6 promoter driving Cre-recombinase expression and constitutively expressing βgalactosidase (β-gal) flanked by loxP sequences followed by GFP. In this model, after administration of 4-hydroxy tamoxifen, most cardiomyocytes became GFP+ and noncardiomyocytes remained β-gal+. Since stem cells were not labeled with GFP, if the percentage of β-gal+ cardiomyocytes increased, the group would assume that the new cardiomyocytes originated from resident stem cells. The proportion of GFP+/β-gal+ cardiomyocytes was evaluated during aging and after ischemic heart injury. One year after the tamoxifen pulse, the GFP+/β-gal+ ratio was not altered, suggesting that nonmyocytes (possibly stem cells) have no contribution to heart regeneration with aging. On the other hand, after myocardial infarction, the group observed a decrease in GFP+ and an increase of β-gal+ cardiomyocytes, suggesting that regeneration was due to resident stem cells[30]. This article supported, at that time, the idea that cardiac repair was promoted by endogenous CSC.
ENDOGENOUS CSC: THE C-KIT+ CONTROVERSY
Beltramiet al[31] suggested that cardiac regeneration was driven by CSC located in special niches in the heart. These CSC had self-renewal properties, expressed the c-kit protein, had a clonal origin, and could differentiatein vitrointo cardiomyocytes, endothelial and smooth muscle cells. When injected into an infarcted border zone, the c-kit+CSC differentiatedin vivointo cardiomyocytes[31]. Human c-kit+CSC, when injected into immunodeficient mice and immunosuppressed rats, formed a chimeric heart improving the cardiac function of infarcted animals (increased ejection fraction and attenuated left ventricular dilation)[32]. The same group reported promising results of cell therapy using c-kit+CSC derived from rats, dogs, and humans[32-35].
They also reported an expressive cardiac renovation of the human adult heart dependent on c-kit+CSC. Contradicting the low regenerative rates demonstrated by Bergmann[29], Kajsturaet al[36] reported that cardiomyocyte renovation rate was greater in women and increased with aging (10%-20% per year at age 20 and 40% per year at age 100), suggesting that all cardiomyocytes were replaced 15 times in women and 11 times in men during one’s lifespan.
The regenerative potential of c-kit+CSC was evaluated in a phase I clinical trial coordinated by Bolliet al[37]. The c-kit+CSC were obtained from the appendage of the right atria of patients with chronic ischemic heart disease submitted to coronary artery bypass surgery. The cells were isolated and cultured three weeks before injection. An increase in ejection fraction (30.3% to 38.5%) and ventricular mass (24% to 30%) of the c-kit-treated group was reported after four months of follow-up[37]. In addition, after 12 mo, the group reported a reduction of the infarcted area in the c-kit+ treated group compared to placebo[38].
Independent groups raised a cautionary note about the regenerative potential of ckit+CSC. They failed to demonstrate the putative cardiac regeneration of c-kit+CSC in animal models and questioned the very existence of these endogenous cell populations in the adult heart[39,40]. Poulyet al[41] identified very few cells expressing c-kit on human atrial biopsies and these cells co-expressed CD45 and tryptase, indicating that they were not stem cells but rather mast cells. In transgenic mice expressing eGFP under the control of the c-kit promoter, c-kit-eGFP expression was observed during different stages of development in embryonic hearts, increasing in number until reaching maximum expression in the postnatal period (2 d of life). After this stage, the expression of c-kit-eGFP cells declined, and they were rarely found in adult hearts. In addition, Talliniet al[39] reported no evidence that adult c-kit+cells differentiated into cardiomyocytes, suggesting that the c-kit+expression shown by other groups after injury was only due to c-kit re-expression in preexisting cardiomyocytes. Jestyet al[42] evaluated the contribution of c-kit+CSC to cardiac regeneration by injecting c-kit+-GFP CSC into infarcted neonatal and adult mice. They observed differentiation of c-kit+-GFP CSC with cardiomyogenic fate only in neonatal animals. In adults, these cells did not contribute to tissue repair[42]. Zarubaet al[40] also showed that only c-kit+CSC derived from neonatal animals could differentiate into cardiomyocytes, promote cardiac repair and engraft in healthy hearts. They suggested that the c-kit+CSC potential to differentiate and repair has age limitations and was not present in c-kit+CSC derived from adults[40].
The controversy surrounding the existence of c-kit+CSC motivated groups to conduct lineage-tracing studies, considering that the main findings which sustained the cardiomyogenic ability of c-kit+CSC cells were based on immunofluorescence assays. These assays used an indirect strategy (primary and secondary antibodies) and therefore could produce false-positive results.
Ellisonet al[43] used a lentiviral system that expressed cre-recombinase under the control of a c-kit promoter to evaluate myocardial repair in yellow fluorescent protein (YFP) reporter mice. After isoproterenol injury, the group showed the presence of new YFP+myocytes (3.4% to 7.7%). Since only cells infected by the virus and that expressed c-kit became YFP+, the presence of YFP+cardiomyocytes led the group to state that adult c-kit+CSC were necessary and sufficient for functional cardiac regeneration and repair. They postulated that the key mechanism of cardiac regeneration after isoproterenol-induced heart injury was by c-kit+CSC differentiation into cardiomyocytes[43]. However, this study was criticized due to methodology issues, including the fact that the partial c-kit promoter used could not properly recapitulate c-kit regulatory elements[44,45].
Other groups used a Cre knockin c-kit transgene approach to identify the contribution of cardiac c-kit+cells for differentiation into cardiomyocytes during mouse development, aging, and after cardiac injury. van Berloet al[46] showed that endogenous c-kit+CSC contributed minimally to generate cardiomyocytes (approximately 0.003% or less if cellular fusion was considered) and concluded that this rare phenomenon could not significantly impact cardiac function. Sultanaet al[47] used multiple reporter genes in mice to show that there was no c-kit co-localized with Nkx2.5 — a gene expressed by cardiac progenitor cells — nor cardiac troponin T — a gene expressed by cardiomyocytes. The group showed that c-kit+cells in murine hearts are not cardiac progenitors but rather endothelial cell progenitors[47]. Liuet al[48] studied the cells immediately after Cre-recombinase induction using an instant ckit lineage tracing model. Using this system, they described that 50% to 70% of labeled cardiomyocytes expressed c-kit 24 h to 48 h after myocardial injury. In this short time, it was unlikely that a progenitor could differentiate into a cardiomyocyte. Moreover, the group showed that c-kit was expressed by cardiomyocytes in adult hearts and concluded that new cardiomyocytes generated after injury were derived from preexisting c-kit+cardiomyocytes and not from CSC[48].
The technical limitations of the chosen tools and models to study c-kit biology, fate, and function are still subject to intense debate[49,50]. To add to the controversy, Vicinanzaet al[51] argued that the previously reported Cre knockin c-kit models had major limitations, such as the inability to identify cells that express c-kit in low levels and the fact that these animal constructions caused haploinsufficiency of the c-kit gene, impairing normal biological regulation and causing a severe defect in CSCdependent myogenesis[46-48,51].
To bypass these technical issues, Heet al[52] developed a system that used two new kit-Cre drivers. This system allowed labeling of all c-kit+cells (even those with low expression) and did not affect the endogenous c-kit gene expression. In agreement with previous studies, the group showed that neither in homeostasis nor after an injury did c-kit+CSC contribute to the generation of new cardiomyocytes[46-48,52].
Thus, despite the many studies describing c-kit+cardiac cells published in the last 20 years, the role of c-kit+CSC in cardiac regeneration and even their existence in adult hearts is highly questionable. Therefore, cardiac regeneration by other endogenous CSC candidates would have to be explored.
CARDIOSPHERES AND CARDIOSPHERE-DERIVED CELLS
Cardiospheres and cardiosphere-derived cells (CDC) are a heterogeneous cell population obtained from explant culture of heart biopsies. Cardiospheres are originated from small phase bright cells, which detach spontaneously from the explants. These structures mimic the tridimensional tissue architecture and preserve the concept that resident stem cells are located in niches inside the organs. Messinaet al[53] described them as clonal, with c-kit in the core and MSC at the periphery. Cardiospheres were obtained from mice and humans and, when injected in the periinfarcted area in mice, induced cardiac regeneration, cell coupling, and improvement of cardiac function[53].
The expansion of cardiospheres as an adherent cell culture showed that a large number of CDC could be obtained from a small piece of heart biopsy, an important goal to translate CDC therapy to the clinical setting. The group reported that CDC differentiated into cardiomyocytes and presented spontaneous beatingin vitroafter ten days of co-culture with rat neonatal cardiomyocytes. When injected at the periinfarcted heart zone, CDC improved the ejection fraction of infarcted animals by directly differentiating into cardiomyocytes and by paracrine effects[54-57]. CDC obtained from children, or newborn cardiac biopsies showed a superior regenerative cardiac capacity to treat infarcted animals compared to those obtained from adults[58,59].
Clinical trials using autologous CDC were conducted to treat patients who suffered recent myocardial infarctions (30 d), aiming to reverse ventricular dysfunction. The intracoronary administration of autologous CDC was safe and showed a discrete decrease of the fibrotic scar when analyzed by magnetic resonance imaging, accompanied by an increase in left ventricular mass with no effect on ejection fraction or end-diastolic or end-systolic volumes at four months and one year after treatment[60,61].
Negative results after administration of cardiospheres or CDC were also observed when utilizing this cell therapy to repair the heart. Liet al[62] failed to observe mouse cardiac improvement after administration of CDC derived from adult hearts. Takeharaet al[63] treated four groups of infarcted pigs: (1) Treated with human CDC (hCDC); (2) Treated with a low release β-fibroblast growth factor (FGF) gel; (3) Treated with both; and (4) Treated with placebo. Group 2 showed cardiac regeneration and improved cardiac function. Group 3 had the same effects reported for group 2 but with higher magnitudes. The group that received isolated hCDC did not exhibit cardiac improvement[63]. In agreement with this, other groups showed that autologous CDC did not improve cardiac function in small or large animal models of myocardial infarction, with CDC, at most, attenuating cardiac remodeling[64-67]. Lineage-tracing of the cardiac explant-derived cells (EDC) was performed using the ventricular myosin light chain MLC2v-Cre/ZEG model. Transgenic EDC were analyzedin vitroby morphology and immunofluorescence for cardiac proteins andin vivoby engraftment and cardiac differentiation. EDC were engrafted into murine hearts but failed to generate cardiomyocytes, suggesting that the strategy to identify cardiac progenitor cells exclusively by morphology was inadequate[68].
Cardiospheres and CDC, similar to c-kit+cells, generated controversial preclinical results and failed to demonstrate robust cardiomyocyte differentiation or improved cardiac function in clinical trials. Thus, the search for the identity of cardiac progenitors continued.
SIDE POPULATION
The capacity to extrude Hoechst33342 and the expression of cell surface ATP binding cassette sub-family G member 2 (ABCG2) are characteristics shared among stem cells present in various tissues, called side population (SP) cells. These cells were also considered putative cardiac progenitor cells with cardiac, endothelial, and smooth muscle differentiation capabilities[69]. However, lineage-tracing using an ABCG2 CreER model demonstrated that this differentiation capacity was present only in embryonic phases and was lost in adulthood, refuting that these cells were CSC candidates responsible for homeostasis and injury response[70,71].
STEM CELL ANTIGEN-1+
The presence of stem cell antigen-1 (Sca-1) in heart cells was first described by Ohet al[72]. This surface marker, also called lymphocyte activation protein-6A (Ly-6A), consists of a glycosylphosphatidylinositol-anchored cell surface protein (GP-AP) of theLy6gene family that is a popular marker used to enrich samples with murine adult hematopoietic stem cells[73]. Sca-1+adult mouse heart cells were negative for CD45, CD34, c-kit, GATA-2, Lmo2, and Flk-1, presenting a distinct phenotype from hematopoietic stem cells, progenitor endothelial, and muscle satellite cells. In addition, Sca-1+ cultured cells were clonal, expressed contractile proteins, and presented spontaneous beating[72,74,75].
Transplantation of Sca-1+cardiac cells showed improved heart function in infarcted mice, promoted by direct differentiation into cardiomyocytes and the release of cytokines such as a soluble VCAM-1, which stimulated angiogenesis, migration, and survivalin vivo[76].
The Sca-1+population was described as a heterogeneous cell population. The subpopulation that expressed Sca-1 in high levels did not differentiate into multiple cell types, while the Sca-1lowshowed direct differentiation into endothelial cells and cardiomyocytesin vitroandin vivo,decreased infarct size, and preserved ventricular function in infarcted mice[77]. Another subpopulation, the Sca-1+CD31-, also showed cardiomyogenic potential in co-culture through a process mediated by cellular coupling with adult cardiomyocytes. This cell population could home to an ischemic heart area using the SDF-1α/CXCR4 pathway and attenuated post-infarct structural ventricular remodeling by direct endothelial and cardiomyocyte differentiation[78-80]. Similar properties were observed for Sca-1+CD45-Isl1+cells obtained from cardiospheres derived from middle-aged mice[81]. The Bmi1+cells, another Sca-1 subpopulation, demonstrated remarkable cardiac regeneration after cell therapy in infarcted mice. Approximately 14% of new mouse cardiomyocytes were observed after myocardial infarction[29,82], contradicting the low rates of cardiac regeneration previously demonstrated in humans.
Nosedaet al[83] further refined the study of Sca-1+murine heart subpopulations using a single-cell expression profile to identify a definitive phenotype for the cardiac stem/progenitor cells. They evaluated the expression of Sca-1, CD31, PDGFRα, and the ability to extrude Hoechst33342 and identified that only SP+Sca-1+CD31-PDGFRα+cells were clonogenic cardiac progenitors.
Even though Sca-1 is not expressed in humans, Goumanset al[84] isolated cardiac progenitor cells from human fetal and adult cardiac biopsies using an antibody that recognized a mouse Sca-1 epitope as a target. These progenitor cells also differentiatedin vitrointo spontaneous beating cardiomyocytes and endothelial cells. Furthermore, when Sca-1+cells derived from fetal tissue were injected in immunodeficient infarcted mice, they also improved cardiac function by direct differentiation into cardiomyocytes[84].
Baileyet al[85] studied a Sca-1 knockout mouse to understand the role of Sca-1 in heart development and cardiac regeneration. These animals showed defects in ventricular contractility and repair, suggesting that the genetic deletion of Sca-1 compromised resident progenitor cells responsible for cardiac repair[85]. A triple transgenic mouse based on the Tet-off Cre system showed that Sca-1+cells played an important role in the generation of cardiomyocytes during homeostasis and after heart injury[86]. These data were not confirmed by independent groups using lineagetracing and fate-mapping studies by multiple sophisticated tools used to genetically trace Sca-1+cells. They proved that these cells did not contribute to cardiac homeostasis or tissue repair by generating cardiomyocytes since Sca-1+cells mainly differentiated into fibroblasts and endothelial cells in response to stress[87-91].
Considering all these studies, it is unlikely that the beneficial results promoted by therapy using Sca-1+cells could be attributed to direct cardiomyocyte differentiation. It is more likely they are linked to angiogenesis stimulated by a paracrine effect. These data reinforce the current leading theory that the generation of new cardiomyocytes during adult life is derived from the proliferation of preexisting cardiomyocytes than from progenitor cells, as already demonstrated in neonatal mice and zebrafish[92-94]. For more details, we suggest the review written by Heet al[95].
IF RESIDENT CARDIAC STEM/PROGENITOR CELLS CANNOT BE FOUND: WHAT NOW?
All putative CSC listed in this review have been discarded as true cardiac stem/ progenitor cells by detailed lineage tracing experiments using sophisticated transgenic models. Furthermore, controversial results surrounded their proposed benefits in preclinical studies, and none resisted the test when applied in the clinical setting. The central idea to replace cardiomyocytes lost due to ischemic or chronic injury persists, but the efforts in the field are now redirected towards obtaining these cardiomyocytesin vitro,from pluripotent stem cells, and then transplant them into the injured heart.
CARDIOMYOCYTES-DERIVED FROM PLURIPOTENT STEM CELLS
Pluripotent stem cells are self-renewing cells that can differentiate into the three embryonic germ layers upon specific stimuli. Until 2006, the sole available source of pluripotent stem cells were embryonic stem cells (ESC). ESC was obtained from the blastocyst's internal mass and could be culturedin vitroas an immortalized lineage[96]. In 2006, Takahashi and Yamanaka[97] made a revolutionary discovery showing how to generate pluripotent stem cells from a somatic cell. They reprogramed fibroblasts, first from mice and, in the next year, from humans, by overexpressingOct-3/4, Sox-2, Klf-4,andc-Myctranscription factors and obtained induced pluripotent stem cells (iPSC), which share the same unique ESC properties[97,98]. Furthermore, the differentiation protocols of pluripotent stem cells into adult cells were improved based on lessons learned from developmental biology[99]. These advances allowed pluripotent stem cells to be efficiently differentiated into cardiomyocytesin vitroby modulating the Wnt pathway, representing an almost inexhaustible source of animalspecific, including human, cardiomyocytes to be used for cell therapy[100].
Transplantation of cardiomyocytes derived from human ESC (hESC-CM) engrafted into infarcted hearts, partially remuscularized myocardial infarctions, improved cardiac performance, and attenuated the remodeling process in infarcted rats and guinea pigs[101,102]. Yuet al[103] proposed that the anti-inflammatory effect promoted by the administration of hESC-CM on immunodeficient female mice submitted to permanent ischemia was a therapeutic mechanism by which these cells improved cardiac function. Human ESC-CM were also evaluated in a non-human primate model submitted to ischemia and reperfusion injury to evaluate the safety, feasibility, and efficacy in a large animal model. As shown in small animals, the administration of 1 billion hESC-CMviaintramyocardial injection improved cardiac function through remuscularization in the non-human primate model[104]. Arrhythmias are a significant concern in cardiac cell therapies. They can result either from the fetal-like phenotype of cardiomyocytes derived from pluripotent stem cells or from a dysfunctional electromechanical coupling between the graft and host cells — as seen with SM[14,105-107]. In this context, the maturation of grafted cells after three months of follow-up, the presence of electromechanical junctions leading to synchronic regular calcium transients between transplanted and host cells, and the absence of fatal ventricular arrhythmias were important observations by Chonget al[104].
The administration of human cardiomyocytes derived from iPSC (hiPSC-CM) also showed promising results in murine, porcine, and non-human primate ischemic cardiomyopathy models[108-110]. Kawamuraet al[108] showed that hiPSC-CM therapy improved cardiac function and attenuated ventricular remodeling of immunosuppressed minipigs submitted to permanent occlusion of the left anterior descending coronary artery after eight weeks of administration of 25 million purified hiPSC-CM. This work suggested that a paracrine mechanism was responsible for the observed results instead of the direct muscularization observed by Chonget al[104]. This conclusion was based on blood flow increase at the infarcted myocardium border due to angiogenesis, probably induced by basic FGF and vascular endothelial growth factor secreted by hiPSC-CM. Also, the majority of the iPSC-CM survived for a short time in infarcted hearts (2 wk), even though some were identified eight weeks after treatment. Therefore, the authors hypothesized that low engraftment was due to insufficient immunosuppressive therapy[108]. The low engraftment and paracrine activity of hiPSC-CM (release of proangiogenic and antiapoptotic cytokines) in the acute myocardial infarction model was corroborated by Onget al[109], who also demonstrated improved cardiac function by magnetic resonance imaging.
A major concern in regenerative medicine is how to avoid immunologic rejection and how to induce immunologic tolerance to cells used in cell therapy, considering that autologous transplantation has to meet manufacture temporal challenges in a clinical setting. The use of major histocompatibility complex (MHC)-matched cells between host and graft was addressed by Shibaet al[110]. They treated five immunosuppressed infarcted heterozygous MHC HT4 monkeys with intramyocardial injection of 4 × 108iPSC-CM per animal. The iPSCs were generated from an MHC haplotype (HT4) homozygous animal. After cell therapy, four of the five animals presented sustained ventricular tachycardia, peaking on day 14 post-transplant and decreasing spontaneously over time. The grafted cells coupled with host cells were still found in infarcted hearts 12 wk after cell treatment with no evidence of immune rejection. The authors suggested that cardiac contractile function was improved by the direct effect of adding new-force generating units. Still, they did not exclude a paracrine mechanism on heart regeneration promoted by allogeneic transplantation of iPSC-CM[110].
During myocardial infarction, aside from cardiomyocytes, other important cell types are also lost, such as endothelial, smooth muscle cells, and fibroblasts. Cell therapy using these cell types was successfully performed in a porcine model of acute myocardial infarction. Endothelial cells (hEC-ESC) and smooth muscle cells derived from human ESC (hSMC-ESC) were transplanted in a fibrin-gel patch into five infarcted pigs submitted to ischemia and reperfusion. Resonance magnetic imaging showed left ventricular functional improvement after a 4-wk follow-up period. The success of vascular cell therapy was attributed to direct neovascularization[111]. The combined use of cardiomyocytes, endothelial and smooth muscle cells derived from hiPSC to treat porcine myocardial infarction was also performed by Yeet al[112]. They showed that this cell combination resulted in cardiomyocyte engraftment and coupling to the host tissue, increased angiogenesis, improved left ventricular function and myocardial metabolism while reducing apoptosis and infarct size with no ventricular arrhythmias observed after four weeks of follow-up.
The end product of all these preclinical experiments with cardiomyocytes derived from pluripotent stem cells was that, for the first time, cardiomyocytes could be producedin vitroon a large scale to meet the requirements for cell therapy, presented robust engraftment of newly-generated units, and promoted angiogenesis, an important additional effect for cardiac regeneration, resulting in improved cardiac function. Despite these significant advances, there are still obstacles to overcome before moving to the clinic, such as managing the risk of arrhythmias, improving cell viability and consequently engraftment, eliminating immune rejection, and identifying the combination of secreted factors that could be responsible for the paracrine effect. Another major problem, not yet completely solved, is the maturation of the PSCderived cardiomyocytes. Cardiomyocytes derived from pluripotent stem cells exhibit an immature phenotype that can be shifted to a more mature state by different approaches such as:in vivografting, three-dimensional constructs using scaffolds and electrostimulation, and manipulation of metabolic pathways. A detailed analysis of such maneuvers is beyond the scope of this review and can be found in Karbassiet al[113].
FUTURE POSSIBILITIES
Cardiac cell therapy has changed significantly in the past 30 years. We have lived through the rise and fall of the endogenous CSC paradigm, whose existence is still subject to debate. Cardiomyocytes derived from pluripotent stem cells emerged as a promising therapeutic alternative, and this cell technology should continue to be investigated to meet the required conditions for clinical application. IPSC technology in human diseases has already been demonstrated to be safe, feasible and showed exciting first results in a patient with macular degeneration[114].
As the paracrine effect has also been suggested as a possible cardiac function improvement mechanism produced by cell therapy, the soluble factors secreted by cells have been investigated. Microvesicles and exosomes — collectively known as extracellular vesicles — were described as small carriers of bioactive products (such as RNA, DNA, proteins, lipids, and cytokines). They are released by cells in different contexts, exerting modulatory effects in diverse biological processes[115]. In ischemic cardiac diseases, extracellular vesicles derived from iPSC-cardiac progenitors and MSC have shown cardioprotective effects in infarcted mice by modulating the inflammatory response and promoting tissue regenerationviamicroRNAs[116-118].
Another emerging approach in cardiac therapy is thein situdirect reprogramming of fibroblasts into cardiomyocytes. The injection of transcription factors Gata4, Mef2c, and Tbx5 directly into mouse myocardium generated new induced cardiomyocytes (iCM), which coupled with the host’s heart cells and decreased the infarct area. When the transcriptomes of iCM were compared to the adult heart cells, they were found to be more similar to those heart cells than to cardiomyocytes generated by the same methodin vitro[119-120]. The direct reprogramming of human cells, which involves another molecular cocktail and epigenetic modulation, is still under investigation[121-124].
Strategies to stimulate the proliferation of endogenous cardiomyocytes are another possibility to regenerate infarcted hearts. MicroRNAs, such as the miR-15 family, regulate the cell cycle of cardiomyocytes. Treatment of infarcted mice with inhibitors of this family (anti-miR-15) has resulted in mitotic cardiomyocytes and improved cardiac function[125-127]. The potential of anti-miR-15, commercially named MGN-1374, is being evaluated in a clinical trial[128]. The small molecule MSI-1436 has also shown interesting results, accelerating heart regeneration in zebrafish and mouse infarction models by the same mechanism — stimulation of preexisting cardiomyocyte proliferation[129].
CONCLUSION
Thirty years of research have taken us a long way in the understanding of cardiac regeneration mechanisms. Unfortunately, this long journey has not yet resulted in the salutary effects of cell-based therapies in the clinical setting. Nonetheless, the long and winding road up to here has shown, as mentioned above, that many options still remain to be explored. Undoubtedly, learning from previous mistakes, we will reach efficacious cell-based therapies to repair and regenerate the injured heart.
杂志排行

World Journal of Stem Cells的其它文章
- Effects of living and metabolically inactive mesenchymal stromal cells and their derivatives on monocytes and macrophages
- Stem cells' centrosomes: How can organelles identified 130 years ago contribute to the future of regenerative medicine?
- Effects of storage media, supplements and cryopreservation methods on quality of stem cells
- Recent advances in stem cell therapy for neurodegenerative disease: Three dimensional tracing and its emerging use
- Current evidence on potential of adipose derived stem cells to enhance bone regeneration and future projection
- Neural stem cell therapy for brain disease