Stem cells' centrosomes: How can organelles identified 130 years ago contribute to the future of regenerative medicine?
2021-10-11AndreasGoutasVarvaraTrachana
Andreas Goutas, Varvara Trachana
Andreas Goutas, Varvara Trachana, Department of Biology, Faculty of Medicine, University of Thessaly, Larisa 41500, Biopolis, Greece
Abstract At the core of regenerative medicine lies the expectation of repair or replacement of damaged tissues or whole organs. Donor scarcity and transplant rejection are major obstacles, and exactly the obstacles that stem cell based therapy promises to overcome. These therapies demand a comprehensive understanding of the asymmetric division of stem cells, i.e. their ability to produce cells with identical potency or differentiated cells. It is believed that with better understanding, researchers will be able to direct stem cell differentiation. Here, we describe extraordinary advances in manipulating stem cell fate that show that we need to focus on the centrosome and the centrosome-derived primary cilium. This belief comes from the fact that this organelle is the vehicle that coordinates the asymmetric division of stem cells. This is supported by studies that report the significant role of the centrosome/cilium in orchestrating signaling pathways that dictate stem cell fate. We anticipate that there is sufficient evidence to place this organelle at the center of efforts that will shape the future of regenerative medicine.
Key Words: Centrosome; Primary cilium; Asymmetry; Self-renewal; Differentiation; Stemness
INTRODUCTION
Stem cells are undifferentiated cells characterized by two unique properties, the capacity to self-renew and maintain a robust stem cell pool, and the ability to differentiate into all types of specialized cells[1]. The differentiation potential of stem cells divide them into four different cell types: totipotent stem cells, which include the zygote and the cells produced by the first few divisions that give rise to the embryo plus extra-embryonic tissues[2]; pluripotent embryonic stem cells (ESCs) present in the inner cell mass of the developing blastocyst that differentiate to form cells in all three germ layers and gametes[3]; multipotent adult stem cells (ASCs) found in many tissues of the adult body, which can differentiate into several cell types that belong to a particular lineage[4], and unipotent stem cells that can only differentiate into one cell type[5]. In 2006, another category of pluripotent stem cells, occurred from disturbing the stable state of differentiated cells and induced them to revert to the level of pluripotency of ESC, was described by a group of Japanese investigators. These induced pluripotent stem cells (iPSC)[6] have increased the hopes for personalized stem cell therapies, as they are derived from patients, divide indefinitely in vitro, and potentially differentiate into any mature cell type. Moreover, their use is not accompanied by the ethical concerns associated with the use of ESCs[7].
At the heart of stem cell therapies is the hope of repair or replacing damaged tissues or whole organs. Donor scarcity, poor quality donor organs, and transplant rejection are the major difficulties faced by regenerative medicine, and exactly those that stem cell-based therapies promise to eventually overcome[8]. Efforts have been directed toward both repair and replacement, and spectacular advances have been reached in the last 20 years[9]. Repair of damaged tissues or organs mainly depends on injection of isolated stem cells that, either because of their proper differentiation or the secretion of biologically active molecules or both, result in tissue/organ structural regeneration and functional improvement[10]. In those attempts, different types of cells have been used, including ESCs, ASCs, and more recently iPSCs. In addition to the ethical issues accompanying the isolation of human (h)ESCs[11], attempts to use ESCs or iPSCs in the clinic have been problematic because of difficulties in achieving full differentiation and function, risk of tumorigenesis, and significant genomic instability[12,13].
ASCs, on the other hand, have been proven safe, and therefore have high expectations of therapeutic potential[14]. As they are scattered throughout the body in bone marrow, adipose tissue, myocardium, skin, umbilical cord blood, and skeletal muscle, ASCs are relatively easily isolated and expandedin vitro[15]. For 50 years, hematopoietic stem cells (HSCs) have been successfully used for the treatment of blood diseases like leukemias and autoimmune disorders[16]. The success of hematopoietic transplantation has raised hopes of the use of other ASCs to treat conditions, such as heart infarction, stroke, spinal cord injury, macular degeneration, diabetes, and skin burns[17]. Despite initial enthusiasm, clinical trials have identified problems such as undesired immune response[18], virus contamination[19], and difficulties with stem cell transport[20]. In addition, the therapeutic use of ASCs requiresin vitroexpansion,which is not free of concerns. Numerous studies demonstrated that ASCs rapidly reach replicative senescence in culture, and that impairs their regenerative potential[21,22].
The only stem cells in routine clinical use are HSCs, as the complications associated with the use of other stem cells have proven greater that originally imagined. It should also be mentioned that the benefits of HSCs and other mesenchymal stem cells (MSCs) depend not only on theirin situdifferentiation to functional tissue cells, but also on their broad repertoire of secreted growth factors, cytokines, chemokines, and other bioactive components, as well as small circular membrane fragments or extracellular vehicles (EVs), enriched in mRNA, microRNA, bioactive lipids, nucleotides, and proteins[23-25]. Paracrine secretion has regenerative properties and has restore confidence in stem cell therapy. The mechanisms are far from being understood, and additional effort is needed to achieve effective, safe, and powerful regenerative approaches that involve the MSC secretome[26].
In addition to exploiting the self-renewal and differentiation properties of stem cells to repair cell and tissue damage or injury, growing tissues or entire organs in the laboratory is a long-term objective of regenerative medicine. The scientific and clinical community is coming closer to this ultimate goal with advances in our knowledge of the factors essential to directing stem cell differentiation and progress in tissue engineering. In fact, specific tissues and even whole organs generated in the laboratory have been transplanted into patients. These include relatively simple laboratorygrown organs, such as skin, bladder, and windpipe[8]. Encouraging advances have been made in the development of bone, cartilage, heart, nerve, and other tissues[27]. New multidisciplinary advances in organ bioengineering based on advances in cell biology, material science, chemistry, molecular biology, engineering, and medicine, include fabrication of synthetic or natural three-dimensional scaffolds used with stem cells and/or bioactive molecules[28]. However, it is evident that reaching the era of off-the-shelf organs awaits a deeper understanding of organogenesis.
Understanding organogenesis requires a detailed description of the decisionmaking machinery that controls the ability of stem cells to balance self-renewal and differentiation, while establishing and maintaining cell fate in the right place at the right time. The role of the orientation of stem cell division has emerged as an important mechanism for determining cell fate. A stereotypical asymmetric cell division (ACD) gives rise to one daughter stem cell with the exact same level of potency as the mother cell and another daughter cell that has acquired a more differentiated state. This unique asymmetry allows the stem cell to self-replicate and maintain the stem cell pool, while at the same time produce numerous differentiated progeny. For ACD to occur, cells must previously establish asymmetry/polarity, which is guided by a variable balance of intrinsicvsextrinsic cues. Several studies over the last 20 years have revealed the previously unappreciated, multifaceted role of centrosomes in interpreting signals from the extracellular as well as the intracellular environment that govern cellular asymmetry[29-36].
CENTROSOMES
More than a century ago, Theodor Boveri portrayed the centrosome as the dynamic center that governs cell division. He was remarkably accurate in describing its basic organization and function as an organelle that consists of a core structure, the centrioles, and an outer centroplasm, now known as the pericentriolar matrix (PCM), that organizes and anchors the “astral rays” that consist of microtubules (MTs)[37]. These nonmembranous organelles function as the MT organizing center (MTOC) of animal cells and therefore regulate vital processes for cell cycle progression, such as mitotic spindle assembly, chromosome segregation and cytokinesis. We now know that this core structure described by Boveri[38] near the end of the 19thcentury,i.e.the pair of centrioles (mother and daughter) and the surrounding PCM consists of around 200-300 proteins, governs MT nucleation, and also regulates cell cycle checkpoints[39]. The centrosome composition is not fixed, as the PCM materials use the MTs anchored to the centrosome as exchange routes. Cycling cells tightly regulate the centrosome cycle, allowing only one duplication round per cell cycle, so that two centrosomes are present in each mitosis. To ensure that, duplication and segregation of centrosomes is coregulated with the chromosome duplication-segregation cycle[40]. The basis of coregulation is the dependence of both key S phase events on cyclin-dependent kinase 2 (Cdk2) activation[41]. The robustness of the coregulation is ensured by the localization of cyclin E-Cdk2 at the centrosomes during G1/S phase when the initiation of DNA synthesis takes place[42].
Centrosome structure and duplication cycle
In proliferating cells, the centrosome needs to duplicate just before or at the onset of S phase so that it forms two new centrosomes that will orchestrate the assembly and organization of the mitotic spindle. Each centrosome consists of two centrioles, a mature mother centriole, and an immature daughter centriole that was assembled during the previous cell cycle, and is about 80% of the length of the mother centriole. Except for length, mother and daughter centrioles are structurally distinct, as the distal surface of mother centrioles is associated with two types of outgrowths, the distal and subdistal appendages that are missing from daughter centrioles. The mother and daughter centrioles are in tight orthogonal association with each other[43]. Disorientation or disengagement, with the loss of the tight association, occurs before completion of cytokinesis, and requires the activity of separase, the protease that is also responsible for the separation of sister chromatids before anaphase[44]. Disengagement is necessary for the initiation of centriole duplication, which takes place before, or at the onset of S phase, where the formation of a new centriole (procentriole) starts at the proximal end of each of the already existing centrioles. The next step is elongation of the procentriole that starts during late S phase. The centriole reaches full length during the following cell cycle. Elongation is followed by maturation in G2, with the recruitment of additional PCM material[45]. Complete maturation of a procentriole into a mother centriole extends over one and a half cell cycles, culminating with the acquisition of distal and subdistal appendages[41]. After duplication of centrosome is complete, the fibrous link between parental centrioles is dissolved to allow centrosomes separation and their migration to opposite poles during prophase of mitosis. As the cell exits mitosis, each new cell inherits one centrosome carrying a mother and a daughter centriole, ready to begin the next centrosome-chromosome duplication cycle[46] (Figure 1).
In interphase, centrioles take on another life; the mother centriole matures and docks below the plasma membrane, where it forms the basal body, which serves as a template for the formation of the axoneme that assembles the primary cilium[37]. The appendages that distinguish the mother centriole from the daughter, drive this process, called ciliogenesis. While the subdistal appendages are involved in organizing the interphase MT cytoskeleton, the distal appendages promote membrane docking and are essential for the formation of the primary cilium. In cycling cells, the cilium cycle follows the cell cycle. The cilium is reabsorbed when cells enter mitosis, which allows the formation of centrosomes and the mitotic spindle assembly [47]. The role of the primary cilium, the antenna-like extension present on the majority of nonproliferating or quiescent cells, has been neglected for many years. It has recently become evident that this organelle has both sensory and signaling functions[48,49] that are of key importance for normal development and health. Highlighting this significance, defects in ciliogenesis are characteristic of a set of ciliopathies that affect organs such as the kidneys, eyes, liver and brain[50-52].
Perturbations of centrosome function have also been linked to carcinogenesis, as they compromise the fidelity of chromosome segregation and can result in aneuploidy. That was the basic premise of Theodore Boveri’s famous theory of cancer development[53], and is still considered an important hallmark[54]. In addition to compromising chromosome segregation by affecting spindle geometry, it has been established that centrosomes contribute to carcinogenesisviaseveral mechanisms that include cellular polarity[55,56], asymmetric centriole inheritance in stem cell lineages[57,58], and ciliary function[59-61]. It is no surprise that centrosome aberrations that might lead to tumorigenesis are related to asymmetries that are intrinsic to their structure and the duplication cycle. The relationship highlights the crucial importance of the asymmetric nature of the centrosome for stem cell physiology. In the following paragraphs we review evidence that supports the maintenance of stem cell renewal and differentiation potential by centrosomes, which direct (1) asymmetric division and distribution of cell fate determinants; and (2) primary cilium-dependent signaling that orchestrates cell fate.
CENTROSOMES AND ASYMMETRIC STEM CELL DIVISION
Stem cells can not only divide symmetrically to expand the stem cell pool, but also asymmetrically. ACD produces one identical stem cell with self-renewal ability and one differentiating cell to produce daughter cells with different fates. This ability of stem cells is the mechanism that balances the need for maintaining the stem cell population with the demand for more differentiated cells, and is vital for tissue homeostasis[62,63]. ACD refers to a polarized/asymmetrical mode of division orchestrated by extrinsic and intrinsic cues that determine the fate of the daughter cells.
ACD depends on cellular polarization
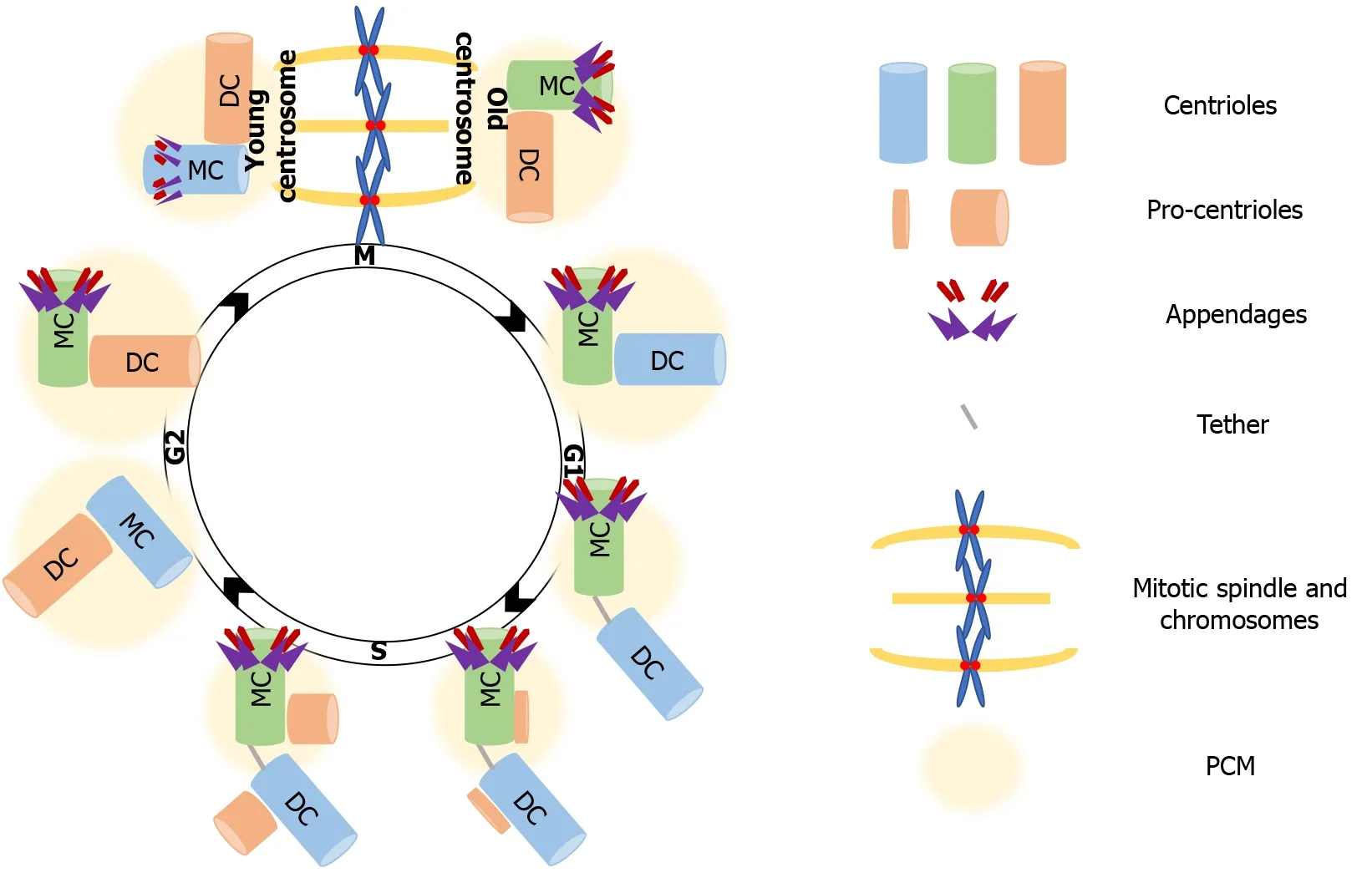
Figure 1 The centrosome cycle.
Extrinsic cues consist a molecular signal repertoire that originates in the extracellular environment that stem cells reside in, called the niche[64]. The asymmetry defined by the niche depends upon the concept of the delivery of self-renewal signals from the niche to the cells within range. For instance, inDrosophila melanogastertestes and ovaries, the niches consist of certain types of postmitotic cells that secrete critical selfrenewal ligands to neighboring cells[65]. The cell fate determinants can even reverse the phenotype of partially differentiated cells to become stem cells again[66]. However, that is not a universal phenomenon. HSCs maintain their stemness even after leaving their niche[1], and neural stem cells can also divide symmetrically outside their niche to produce identical self-renewing progeny[67].
The latter evidence highlights the importance of intrinsic cell fate determinants and turns the focus to intracellular cues that are characterized by or provide asymmetry. In other words, the simplest way of producing two different daughter cells, is to distribute fate regulators asymmetrically by polarizing the interphase intracellular environment and ensuring that the determinants will be inherited asymmetrically by properly orienting the mitotic spindle during division. The proper orientation of the spindle to ensure asymmetric division of cell fate determinants, can also been seen as the process by which extrinsic and intrinsic cues merge. Drosophila male germline stem cells (GSCs) are attached to their niche, which provides the signaling ligands necessary for retaining stem cell identity[64]. The asymmetry provided by the niche to the attached progeny is guaranteed by the GSCs orienting their spindle perpendicular to the hub cells, so that one daughter cell maintains the attachment to the cell hub, whereas the sibling cell initiates differentiation because of losing the attachment[68]. The latter emphasizes the fact that ACDs are achieved by polarization of fate determinants coupled with proper spindle orientation[69]. The best described example of polarization of cell fate determinants is also provided by studies in Drosophila. It has been shown that Drosophila neuroblasts produce fate determinants that remain inactive during interphase, are distributed in a polarized manner, and during division the mitotic spindle is oriented in such a way so that these determinants are inherited by only one of the two daughter cells, which will become the ganglion mother cell[70,71].
Centrosomes serve as means of cellular polarization
Importantly, both cellular polarization as well as orientation of the mitotic spindle depend upon centrosome function. For polarization to occur the most essential requirement is the existence of MTs, and in essentially all non-terminally differentiated somatic cell as well as male germline animal cells, MT organization is governed by centrosomes[72]. For instance, it is well established that centrosome positioning governs the localization of several subcellular compartments, such as the Golgi apparatus, by controling of the nucleation and anchoring of MTs. It has recently been shown that the centrosome might also promote actin filament assembly. As MTs and actin filaments are the two main cytoskeleton networks supporting cell polarity, the latter makes the centrosome the master regulator of intracellular architecture[73].
It is clear that, besides having this fundamental effect on cell geometry, the centrosome determines the position at which the spindle poles will form and how the mitotic spindle will be oriented. Correct orientation of the mitotic spindle ensures not only faithful segregation of chromosomes but also proper segregation of molecules defining cell fate[74]. Various MT subpopulations,i.e.kinetochores, interpolar and astral MTs, are involved in controlling the process[75]. For instance, astral MT nucleation, stability, and dynamics as well as anchoring at the cortex are of particular importance in order to achieve proper orientation of the mitotic spindle[74].
More than building polarity/asymmetry, centrosomes provide a mechanism that maintains and transmits differential cell fate information, which also explains how cellular memory is passed on from one cell to the next during division[76]. As mentioned, because of the way centrosomes duplicate, the centrioles within each centrosome can be distinguished by age, one is formed in the preceding cell cycle, and the other is assembled at least one cycle earlier. After duplication, the centrosome that retains the most recently built centriole is the young centrosome and the other one is the old centrosome (see Figure 1). The old centrosome consists of the older mother centriole, which harbors completely mature distal and subdistal appendages. Because the subdistal appendages are the major site for MTs anchoring, the old centrosome typically has higher MT organizing activity (MTOC) than the young centrosome, which contains the recently formed mother centriole. Even more, several proteins such ninein (NIN), Cep164 and outer dense fiber protein 2 (ODF2) were found to be localized to the mother centriole, whereas centrobin (Cnb) localizes only to the daughter centriole[77-80]. This differential protein composition enhances the asymmetry of the old and young centrosomes, which in turn ensures that the daughter cells arising following division are “born differently”, as one receives the young centrosome and the other receives the old centrosome. Also, this asymmetry carries the intriguing assumption that it would be also functionally relevant to stem cell ACD.
The asymmetric segregation of the centrosomes that defines the fate of progeny has been best described in GSC, Drosophila neuroblasts, and mouse neural progenitors[81-83]. Those studies provided a narration of the asymmetric centrosome cycle, which depends on differential MTOC activity and in turn on centrosome age, as already mentioned. Specifically, as described for Drosophila neuroblasts, the young centrosome -the one containing the younger mother centriole- maintains its MTOC ability throughout interphase, whereas the old centrosome -containing the older mother centriole- downregulates its ability to nucleate MTs as the neuroblasts enter interphase. That helps orient the mitotic spindle along the neuroblast apical-basal polarity axis, as the young centrosome with active MTOC ability, remains tethered to the apical neuroblast cortex. The inactive old centrosome is displaced form the apical cortex as its centriole downregulates MTOC activity through the “shedding” of its PCM content. In that way, the apical centrosome will always be the young centrosome, and will be inherited by the self-renewed progeny, while the old centrosome segregates into the more differentiated daughter cell. Biased centrosome segregation also takes place in male Drosophila GSCs; but in this cell type is the old centrosome that retains the MTOC activity and therefore maintains its localization near the stem cell niche, ensuring that the self-renewal ability is passed on to the proximal progeny[34,36,82,84-87].
Even though it is true that the above mentioned mechanism of biased centrosome inheritance was originally described for a few cell types, several studies have shown that human cells are probably not an exception. For example, studies have revealed the dependance of MTOC ability on specific centrosome components in human cells. The human daughter centriole-associated ciliopathy protein, Cep120, has been shown to have a critical role in MTOC activity, as its depletion results in accumulation of PCM components. Elevated PCM levels result in increased MTOC activity at the centrosome, which is crucial for centrosome homeostasis, potentially underlies the pathogenesis of ciliopathies, and provides further evidence of the dependence of ACD on centrosomes[88].
Strong evidence has been provided for involvement of the Wnt pathway in the determination of cell fate in humans. Several important Wnt pathway components, such as disheveled 2, which actually transmits the Wnt signal, was found to localize at the centrosome and to regulate spindle orientation[89]. The latter is critical for determining the plane of cell division and defining whether a cell remains within a particular environment, such as the niche, therefore controlling cell fate. The importance of Wnt signaling in the ACD of human skeletal stem cells (hSSCs) was highlighted in a recent study. It was shown that covalently immobilizing Wnt factors onto synthetic materials can polarize single dividing hSSCs, orient the spindle, and simultaneously generate a Wnt-proximal hSSC and a differentiation-prone Wnt-distal cell[90]. The study emphasizes the importance of deciphering the nature and function of centrosomes for the development of promising approaches for tissue repair.
Moreover, recent advances of centriole biology support the universality of biased centrosome segregation[91-95]. Firstly, the novel concept of PCM as a molecular assembly formedvialiquid-liquid phase separation[96] is an outstanding paradigm that makes the asymmetric nature of youngvsold centrosomes even more pragmatic and relevant[97,98]. Even more, targeted cotranslation is another concept that adds to the fascinating idea of centrosome-dependent ACD. In zebrafish and various human cell types, the mRNAs of key centrosome scaffold proteins such as pericentrin (PCNT) and nuclear mitotic apparatus protein 1 (NUMA1), among others were found to be located on the centrosome where they are translated during mitosis[99-101]. Thein situtranslation (1) optimizes centrosome maturation, as its core proteins are manufactured at their destination compartment, and (2) adds a sophisticated layer of regulation of centrosome asymmetry that could prove critical for ACD.In situtranslation provides insights to the mechanism(s)viawhich mutations in PCNT, for example, cause primary microcephaly phenotypes that are thought to arise from proliferation defects in neural progenitors[102]. Moreover, the above studies[99-101], identified eight mRNAs that localize in the centrosomes of human cells. These mRNAs code for centrosome proteins PCNT, NUMA1, CCDC88C, NIN, BICD2, HMMR, CEP350, and ASPM that regulate centrosome maturation, spindle positioning, and MT dynamics. Given the importance of these proteins in centrosome biology, which is indicated by theirin situtranslation, it would be interesting to elucidate their specific role in centrosome-dependent ACD, which will in turn open new horizons in manipulating the determination of cell fate.
Old vs young centrosome: Asymmetries are functionally relevant
The differential segregation of old and young centrosomes in asymmetrically dividing cells is accompanied by functionally relevant consequences. The old centrosome carries ciliary membrane when it is internalized before mitosis. Because of that, the daughter cell that inherits this centrosome will form a primary cilium before its sibling does[103]. The consequence is that the cell that inherited the old centrosome accumulates primary cilium-associated smoothened (SMO) and experiences higher hedgehog (Hh) signaling, that has been demonstrated to promote stem cell identity. On the contrary, the sibling cell that inherits the young centrosome loses self-renewal ability in response to lower Hh signaling and commits to differentiation[103].
As earlier mentioned, the old and young centrosomes differ not only in age and their ability to organize MTs, but also in their molecular composition,i.e.proteins and mRNAs that could serve as fate determinants[104,105]. A well-designed study by Lambert and Nagy[104] showed that fate-determining mRNAs are attached to one of the centrosomes during cell division in mollusk embryos. Those mRNAs are inherited by only one daughter cellviathe asymmetric segregation of the centrosomes, and define the embryonic patterning during mollusk development. A recent study demonstrated that Mindbomb1, a Notch ligand activity regulator, was found to localize onto the daughter centriole in chick neural progenitors, and that the daughter cells that receive this centriole after ACD differentiate into neurons[106].
Another interesting asymmetry that is associated with centrosomes has been reported to accompany cytokinesis. At the end of cytokinesis, the midbody ring is inherited by one of the daughter cells, which studies in HeLa cells have shown, is the cell that inherits the old centrosome[107]. Interestingly, studies with stem cells revealed a correlation between midbody inheritance and self-renewal ability[107-109]. Even though that is an interesting notion, the exact role that the midbody or midbodyassociated molecules have in regulating self-renewal is missing. Similarly, the aggresome, a large structure that accumulates damaged or misfolded proteins, was also observed to be associated with centrosomes and to be inherited, together with the young centrosome, during ESC division by the differentiated progeny[110]. Again, it was implied that the aggresome acts as a cell fate determinant without providing a mechanistic insight that would explain such a function.
Interestingly, a recent study provided even more direct evidence linking the old centrosome’s composition to pluripotency maintenance. It was shown that NANOG, the protein that, together with SOX2 and OCT4, has a fundamental role in defining stemness, localizes in the cytoplasm on the appendages of the mother centriole in human tumor cell lines, fibroblasts and hESCs[111]. Even though this important study clearly demonstrated the association of NANOG with the old centrosome, the assumption of its role in centrosome maturation is lacking direct evidence. Nevertheless, these observations gave birth and provide support of the immortal centrosome hypothesis, which proposes that the daughter cell that retains the older centrosome has self-renewal properties[112]. The hypothesis is elegant, but not globally supported. In Drosophila neuroblasts and female GSCs, the cell that inherits the young centrosome maintains self-renewing[109,113]. A similar pattern of asymmetric inheritance of the centrosome was found in human cancer stem cells. It was shown that in human neuroblastoma cell lines, the young centrosome is inherited by the NUMA-retaining cell, which also possesses greater proliferation ability[114]. In any case, the fact that either the young or the old centrosome is associated with selfrenewal progeny means that it is the asymmetric nature of the centrosome that contributes to determination of cell fate. Recent studies of the molecular composition of centrosomes provide additional evidence supporting asymmetry. Several proteins reported to be associated with stem cell centrosomes could prove to be functionally relevant to asymmetry and have crucial roles in determining cell fate. For example, NIN is a protein reported to be specific to the mother centriole[79,115] and to accompany inheritance of the old centrosome in radial glial progenitor cells that retain pluripotency in mice[83]. It was also demonstrated that Klp10A, an MT-depolymerizing kinesin, is located on the centrosomes of stem cells but not the centrosomes of differentiating germ cells in Drosophila male germline. Moreover, depletion of Klp10A resulted in an abnormally elongated old centrosome compared with the younger one, which in turn gave rise to asymmetric daughter cell size, with a bigger GSC and a smaller differentiating gonial blast. Even though these results did not reveal the exact significance of centrosome asymmetry, they imply that the old centrosome (or older mother centriole) possesses an ability that is missing from the young centrosome[116]. A recent study identified another mouse neural stem cell centrosome protein, AKNA, that is found only on the subdistal appendages of the mother centriole, and has a vital role in promoting MT nucleation and growth[117].Alms1a, the Drosophila homologue of the gene responsible for the Alstrom syndrome, a characteristic human ciliopathy, is another mother centriole-specific protein that was recently identified and found to interact with both Klp10A and Sak[118], the Drosophila homologue of Polo-like kinase 4 (Plk4), the master regulator of centriole duplication[119]. In addition to centriole duplication, Plk4 was recently reported to support the establishment of centrosome asymmetry. In Drosophila neural stem cells, Plk4 phosphorylates Spd2 on the mother centriole, which is important not only for centrosome asymmetry but also for proper mitotic spindle positioning[120]. These findings add further knowledge of the unique characteristics of old and young centrosomes that will help to elucidate how centrosomal asymmetry drives asymmetric stem cell division, by identifying the centrosome molecules and their functions that drive self-renewalvsdifferentiation. And by deciphering how cell fate determination is controlled, new research directions will open, aiming at producing cells specific for use in tissue repair and regeneration.
PRIMARY CILIA AND STEM CELL DIFFERENTIATION
Among the centrosome asymmetries, no difference is more remarkable than the unique ability of the mother centriole to dictate the formation of the primary cilium. The primary or nonmotile cilium is an organelle consisting of MTs surrounded by a specialized membrane that carries signal receptors. It extends from the apical surface of nearly all vertebrate cells, and forms when the basal body docks on the membrane[121].
Mother centriole and ciliogenesis
Ciliogenesis, is known to be entirely dependent on the mother centriole appendages, with the distal appendages promoting mother centriole to basal body maturation and membrane docking. The subdistal appendages direct cilium positioning (Figure 2)[37].

Figure 2 The primary cilium.
The process is rather complex and involves multiple steps that include (1) transport of preciliary vesicles to the basal body, associating with the distal appendages, and fusing to form a larger vesicle; (2) enrichment of the ciliary vesicle with membrane proteins that promote ciliary membrane expansion and selective trafficking of proteins to the cilium; (3) extension of the centriole/basal body MTs to form the axoneme of the cilium; and (4) the forming of a transition zone that partitions the cilium from the cell body[47,122]. Two distinct ciliogenic pathways have been described. The intracytoplasmic and plasma membrane associated pathways, differ in the position of the formation of the preciliary vesicles,i.e.the cytoplasm or plasma membrane, and not in the basic steps[61]. Ciliogenesis results in the construction of a distinctive microenvironment within the primary cilium that facilitates the transduction of extracellular initiated signals. Many components of the cilium, both regulatory and structural, participate in receiving and interpreting a variety of different extracellular cues[123]. As already mentioned, aberrations in ciliogenesis or dysfunction of primary ciliaassociated signaling is linked to several human pathologies, These ciliopathies highlight the functional significance of the mother centriole that was neglected for a long time and has recently experienced a renaissance. This renaissance is believed to be attributed to accumulating evidence that support its role as the cell’s "antenna", which receives and integrates signals from the extracellular environment that regulate development, cell polarity, and importantly, cell identity[124].
The primary cilium inherits asymmetries
Many studies have reported the presence of primary cilia in a variety of stem and progenitor cells. Aberrations or alterations in their structure/expression, length, and/or protein composition highlight their significance for stem cell function[125-129]. It is becoming evident that the function of the primary cilium in signaling is of crucial importance when it comes to the determination of stem cell fate. As already mentioned, in dividing mouse radial glial progenitor cells, the primary cilium is not completely disassembled prior to cell division. Fragments stay attached to the old centrosome, which includes the older mother centriole (see Figure 1). The latter is believed to result in more rapid formation of a primary cilium in that daughter cell than in its sibling, as its old centrosome had controlled the formation of a primary cilium in a previous cycle. Even more, it was also demonstrated that the old centriole responds to signaling that promotes stemness[103]. A recent study, using live-cell imaging analysis, demonstrated that cilia grow faster from older centrosomes associated with a ciliary remnant than from “naked” centrosomes that lack a remnant[130]. Additionally, this study clearly demonstrated that the remnants were associated with the distal appendages of the mother centriole and that overexpression of active Nek2A kinase prematurely displaced distal appendages from interphase centrosomes. As previously noted, inheritance of the ciliary remnants seems to give to the cells a temporal advantage in reforming the cilium and therefore becoming responsive to Hh ligands[103]; the latter could prove essential in paving ways to manipulate cell fate determination.
Given the above, a recent study by Vestergaard and colleagues [131] that showed that transcription factors known to be associated with pluripotency, such as SOX2 and NANOG, are located on the primary cilium of hESC did not come as a surprise. Specifically, the study describes the technique used to examine the spatiotemporal regulation and localization of those transcription factors and revealed that in addition to the expected nuclear location, SOX2 and NANOG were associated with a subset of hESC primary cilia. Even though the study is important for indicating a functional relationship between primary cilia and differentiation and/or self-renewal processes, it lacks an explanation of why SOX2 and NANOG were found in some hESC primary cilia and not in all of them. It remains to be shown whether that was a technical limitation of the study or if it is functionally relevant.
Primary cilium-associated signaling pathways determine cell fate
A variety of signaling pathways that are crucial for cell fate determination and differentiation have been reported to be associated with or mediated by the primary cilia of human stem cells[132-135]. The most relevant are the Hh and Wnt signaling pathways, which have already been mentioned[136]. Hh signaling has been shown to be of critical importance in organogenesis, as it promotes stem cell proliferation and migration[139-142]. Two of the most important proteins for Hh signaling, Patched (PTCH) and SMO, were found to be located in the primary cilium. Briefly, when Sonic Hh is present it binds PTCH, thus allowing SMO to move into the ciliary axoneme and activate the glioma-associated oncogene transcription factor (GLI). For a detailed description see Kopinkeet al[143]. Briefly, the cilium is believed to act as a mediator of the trafficking and accumulation of SMO and GLI proteins in the context of Hh signaling during development and regeneration.
Like the Hh signaling pathway, Wnt signaling is considered extremely crucial for cell fate determination[144]. Wnt signaling includes both canonical and noncanonical Wnt pathways. Canonical Wnt signaling controls cell proliferation and cell fate, and defects have been associated with cancer development. Noncanonical Wnt signaling is thought to give shape to tissues by control of cell migration and orientation driven by cell polarization and ACD. In noncanonical Wnt signaling the receptor of Wnt signals, Frizzled protein, was found to be located on the membrane of the primary cilium. The downstream activity of the Wnt proteins inversin and disheveled is also located at the base of the cilium[144,145]. Recent evidence suggests that components of noncanonical Wnt signaling interact or are associated with the primary cilium[146].
Recent findings also report the dependance of the platelet-derived growth factor (PDGF)[137] and transforming growth factor beta (TGF-β) signaling pathways on the primary cilium[133,138]. The PDGF signaling pathway is considered to be of major importance for wound healing and cancer development, and has been implicated in cell migration and differentiation[147]. PDGF signaling depends upon the interaction between PDGF-AA ligand and its receptor PDGFRα, which was found to occur on the primary cilium membrane. This interaction may be the best described example of the function of the primary cilium as a chemical antenna, as its orientation depended on the concentration of the PDGFRα receptor[148]. TGF-β signaling is also linked to cell proliferation and differentiation. It is particularly important in epithelial-mesenchymal transition (EMT), a procedure that is mediated by shear stress activating TGF-β that is located on the primary cilium[149,150]. The downstream proteins of TGF-β signaling, SMAD 2/3 and extracellular signal-regulated kinase 1/2 (Erk1/2), have also been found at the base of the cilium[133], further supporting its importance in this pathway.
The significance of cilia-mediated signal transduction was further emphasized by a study that investigated hESC mesendoderm and neuroectoderm (NE) fate decisions. It was demonstrated that a specific ciliation pattern occurred within the first 24 h that, coupled with G1 phase lengthening, induced NE lineage specification before any other neural markers were expressed. Notably, it was further shown that cilia formation in NE precursors was accompanied by increased autophagy that resulted in NRF2-mediated transcription inactivation and repression of the expression of pluripotency genes OCT4 and NANOG that allow lineage commitment toward NE[151].
The critical significance of the above signaling molecules being present at the primary cilium was further supported by reports of its fundamental role in defining the offspring of different progenitors. When muscle-resident fibro/adipocyte progenitors (FAP) are injured or aged, proliferation is shifted towards the production of adipocytes, which causes muscle to be replaced by fat. A recent study demonstrated that the process was directed by ciliary Hh signaling[152]. The same study also demonstrated that preventing ciliation in FAP resulted in inhibition of intramuscular adipogenesis and enhanced myofiber regeneration after injury in a Duchenne muscular dystrophy mouse model. A study of electrical field stimulation (EFS)-enhanced osteogenesis of human adipose-derived stem cells (hASC) demonstrated that if the molecular composition of the primary cilium was disrupted, the ability of hASC to detect electrical field signals was compromised. The same study also reported evidence of the primary cilium as a key calcium-signaling module during EFSosteogenesis[153]. Another recent study added to the above by demonstrating that calcium induction triggered ciliogenesis and adipogenic differentiation of human MSCs by negatively regulating Wnt5a/β-catenin signaling[154].
Another study of hASCs showed the potential implication of ciliary signaling in the pathogenesis of obesity. It was reported that obese hASCs had shortened cilia, and were unable to respond properly to stimuli[155]. Interestingly, another study in obese patients showed that treatment with inhibitors of Aurora A kinase or Erk1/2 rescued both the length and functionality of primary cilia and increased the expression of genes related to self-renewal/stemness. The findings have clinical importance for autologous MSC-based therapies[156]. Further studies revealed that the above cilia aberrations were associated with a deficiency in Hh signaling that affected hASC differentiation capacity. The data support the potential of novel therapies for obesity and associated pathologies[155]. The impact of ciliary Hh signaling in tissue regeneration and tumorigenesis was described in a recent study of the importance of epithelial-EMT programming in stemness. The stemness of both mammary stem cells and their neoplastic counterparts, mammary tumor initiating cells, in the mammary epithelium seem to depend on the EMT program, which in turn relies on primary cilia formation and Hh signaling[157].
Stemness dependance on the primary cilium was also investigated in a recent study in which the authors silenced the expression of two of its components, the ciliary proteins IFT172 and KIF3A, in MSCs. The outcome of siRNA-based knockdowns was the production of fewer and shorter cilia, increased proliferation ability of MSC and reduction of the expression of the stem cell markers OCT4, NANOG, and SOX2[158]. The results suggest the dependance of stemness maintenance on proper cilia function and signaling. Similarly, a recent study reported the dependance on cilia-specific genes of hematopoietic stem and progenitor cell (HSPC) function in the hemogenic endothelium (HE) of zebrafish embryos. The authors described the role of ciliamediated Notch signaling in HSPC asymmetric division in the production of mature blood cells as well as self-renewing progeny[159].
Not only structure and signaling but also proper disassembly of the primary cilium was reported to be an important factor in stem cell function. It is known that a mutation in the centrosomal-P4.1-associated protein (CPAP) is linked to Seckel syndrome microcephaly[160] and possibly to neural progenitor cell (NPC) dysfunction. It was demonstrated recently that CPAP serves as a scaffold protein that promotes timely cilium disassembly, and mutation results in retarded cilium disassembly as well as delayed cell cycle re-entry and therefore premature differentiation of NPC[161]. The latter further emphasizes the important role that the primary cilium has in ACD that maintains tissue homeostasis.
A recent study investigated the function of cilia-dependent signaling in regeneration and repair of fractured bone. Interestingly, the authors showed that delayed fracture healing in smokers might be attributed to dysfunctional ciliarymediated TGF-β signaling in MSC[162]. Besides TGF-β, Hh signaling and intraflagellar transport (IFT) were reported to be essential for bone development. IFT moves nonmembrane-bound particles from the cytoplasm to the tip of the cilium and is considered crucial for cilium assembly and maintenance[163,164]. A recent study showed that IFT proteins regulated Hh signaling in osteoblasts (OBs), and their silencing resulted in impaired OB differentiation and subsequent craniofacial and skeletal abnormalities[165].
As previously mentioned, MSCs have been accepted as vital for tissue homeostasis and regenerative medicine, as they are present in almost all tissues, are easily isolated, can differentiate into almost any cell lineage, and can be cultured on specific scaffolds used for tissue reconstruction[4,136,166]. Even though initial studies of MSC-based regenerative approaches focused on the musculoskeletal system, studies have recently been expanded to include other tissues, like the nervous system, heart, liver, cornea, and trachea[136]. Many studies have used this type of adult stem cells to explore the role of the primary cilium in directing regeneration and repair. For example, in one of the first studies, Corbitet al.[167] demonstrated that knocking down the cilia protein Kif3a resulted in disruption of the proper structure of the cilium and enhanced canonical Wnt signaling. Similarly, siRNA knockdown of IFT88, another primary cilium-associated protein, was also demonstrated to compromise the osteogenic, chondrogenic and adipogenic differentiation potential of MSCs[126]. Knockdown of another cilia-associated protein, polycystin-1 in human adipose tissue-derived MSC (hASC) resulted in a downregulation of osteocalcin gene, diminished calcium accretion, and reduced alkaline phosphatase activity that abrogated hΑSC-dependent bone regeneration and repair abilities[168]. MSC cilium structure and the activity of its associated proteins in the control of cell differentiation were investigated in a study analyzing changes in ciliary length. It was reported that MSCs cultured in adipogenic differentiation medium exhibited an elongation of their primary cilia with subsequent upregulation of nuclear PPARγ levels and recruitment of IGF-1Rβ to the cilium, thus contributing to expanding our knowledge of ciliary protein function[169].
Even more essential, the role that MSCs could have in tissue engineering and regenerative medicine was highlighted in a study that investigated the effect of substrate environment architecture on MSC phenotype determination[170]. It was shown that substrate architecture can induce changes in cytoskeletal tension that in turn influence primary cilium signaling. Specifically, it was demonstrated that MSCs cultured on grooved surfaces had more elongated and aligned cilia. It was concluded that the specific architecture enhanced ciliogenesis and suppressed MSC proliferationviainhibition of canonical Wnt signaling[171]. Another recent study described the dynamic sensory abilities of hASC primary cilia and the importance of manipulating those abilities. The authors found that hASC cilia length and cilia conformation varied in response to culture conditions (e.g., complete growth, osteogenic differentiation, or adipogenic differentiation culture medium) with the longest cilia expressed in cells differentiating into adipocytes. Importantly, they showed that cyclic tensile strain enhanced hASC osteogenic differentiation while suppressing adipogenic differentiation[172]. The study highlights the importance of the primary cilium in lineage specification and therefore its role as a novel target in attempts to manipulate hASC for tissue engineering applications.
CONCLUSION
Based on all the above, it is safe to say that cellular asymmetry and asymmetric distribution of cell fate determinants as well as ACD define stemness. It is becoming more than evident that the centrosome and the centrosome-derived primary cilium provide an excellent vehicle to serve this asymmetry. The centrosome and centrosome-derived primary cilium illustrates the extraordinary ability of stem cells to maintain the crucial balance between self-renewal and differentiation. As studies regarding stem cell centrosomes and cilia accumulate, we are reaching a better understanding of the requirement of the presence of these structures for orchestrating receiving, interpreting and transducing signals. Essentially, centrosome-dependent signaling -by directing changes in stem cell morphology, gene expression, and cytoskeletal organization- ultimately determine stem cell differentiation. Hence, it is tempting to envision procedures aiming to manipulate and change centrosome composition and/or cilium architecture and trafficking, as means of controlling the direction of differentiation in the context of tissue engineering and regenerative medicine. Already various methods that aim to guide cell phenotype, including chemical or mechanical stimulation as well as modulation of the architecture, composition and/or dimensionality of the substrate microenvironment, have been reported[136]. From what was presented here, it seems that those manipulations, intentionally or unintentionally, directly or indirectly, aimed at exploiting the functions of the centrosome/ cilium. For this reason, we dare to predict that this 130-year-old organelle, originally called the centrosome, in order to acknowledge its location near the geometrical center of the interphase cell, will be at the center of efforts that will shape the future of regenerative medicine.
ACKNOWLEDGEMENTS
The authors wish to thank Dr. Taylor M for editing the manuscript.
杂志排行

World Journal of Stem Cells的其它文章
- Effects of living and metabolically inactive mesenchymal stromal cells and their derivatives on monocytes and macrophages
- Effects of storage media, supplements and cryopreservation methods on quality of stem cells
- Recent advances in stem cell therapy for neurodegenerative disease: Three dimensional tracing and its emerging use
- Stem cell therapies in cardiac diseases: Current status and future possibilities
- Current evidence on potential of adipose derived stem cells to enhance bone regeneration and future projection
- Neural stem cell therapy for brain disease