Recent advances in stem cell therapy for neurodegenerative disease: Three dimensional tracing and its emerging use
2021-10-11IlKwonKimJunHeeParkBomiKimKiChulHwangByeongWookSong
Il-Kwon Kim, Jun-Hee Park, Bomi Kim, Ki-Chul Hwang, Byeong-Wook Song
Il-Kwon Kim, Jun-Hee Park, Bomi Kim, Ki-Chul Hwang, Byeong-Wook Song, Institute for Bio-Medical Convergence, Catholic Kwandong University International St. Mary’s Hospital, Incheon Metropolitan City 22711, South Korea
Il-Kwon Kim, Ki-Chul Hwang, Byeong-Wook Song, Institute for Bio-Medical Convergence, College of Medicine, Catholic Kwandong University, Gangwon-do 25601, South Korea
Abstract Neurodegenerative disease is a brain disorder caused by the loss of structure and function of neurons that lowers the quality of human life. Apart from the limited potential for endogenous regeneration, stem cell-based therapies hold considerable promise for maintaining homeostatic tissue regeneration and enhancing plasticity. Despite many studies, there remains insufficient evidence for stem cell tracing and its correlation with endogenous neural cells in brain tissue with threedimensional structures. Recent advancements in tissue optical clearing techniques have been developed to overcome the existing shortcomings of cross-sectional tissue analysis in thick and complex tissues. This review focuses on recent progress of stem cell treatments to improve neurodegenerative disease, and introduces tissue optical clearing techniques that can implement a threedimensional image as a proof of concept. This review provides a more comprehensive understanding of stem cell tracing that will play an important role in evaluating therapeutic efficacy and cellular interrelationship for regeneration in neurodegenerative diseases.
Key Words: Cell tracing; Neurodegenerative disease; Stem cells; Three-dimensional imaging; Tissue clearing
INTRODUCTION
Most brain disorders lead to irreversible consequences in intra- and inter-cellular responses depending on their severity, which commonly causes deterioration of physical or intellectual function. In general, it is known that the adult central nervous system is not capable of neurogenesis, but recent research on stem cells has negated this precept[1,2]. To improve neural regeneration to replace damaged neural cells and/or re-establish dendritic connections, two basic strategies have been established over the past two decades[3-5]. First, endogenous neural stem cells (NSCs) participate in the self-repair process in the subventricular zone (SVZ) lining the lateral ventricles and the subgranular zone within the dentate gyrus of the hippocampus, despite limitations in cell number and regenerative ability[6]. Even if symptomatic treatment is performed when the boundaries of endogenous regeneration in the brain are crossed, they have limited implications, including sustainability and efficiency in the repair of neurodegenerative diseases. Second, exogenous transplantation of stem cells is expected to become a source of neurogenesis. Stem cells derived from pluripotent ‘embryonic’ stem cells (ESCs), which are more lineage-committed reprogrammed ‘embryonic-like’ pluripotent stem cells (PSCs) have been used as a therapeutic source in neurodegenerative diseases[7,8]. The fundamental mechanism underlying all therapies is a positive regulation of progressive loss of brain structure, function, or neuronal survival. Although stem cell-derived NSCs or neural progenitors can affect cell replacement therapy, direct transplantation of stem cells or stem cell-free therapy is mostly known to be exerted through paracrine effects, including cytokines, chemokines, and neurotrophic factors at the molecular level[9-11]. Unfortunately, the therapy currently available does not fully account for the mechanism of stem cell functionin vivo, and it does not prove the relationship between exogenous stem cells and existing brain cells from the site of neurodegenerative disease[12]. To analyze an interconnected network with a molecular biological approach, an integrative description of the microenvironment needs to comprehend the three-dimensional structure of the brain containing exogenous stem cells.
This review will focus on recent improvements of stem cell therapy for neurodegenerative disease, the methodological approach of cell tracking for the definition of stem cell proof-of-concept (PoC), and on the advanced technique of cell tracking for a threedimensional structure description after stem cell treatment. This clarification will influence future studies by providing insights into the three-dimensional structure approach of stem cell tracing for many therapies of neurodegenerative diseases.
STEM CELL THERAPY IN NEURODEGENERATIVE DISEASE
Neurodegenerative diseases are mainly classified by clinical characteristics which are based on major symptoms and the site of involvement, including Alzheimer’s disease (AD), Parkinson’s disease (PD), and Huntington’s disease (HD). AD and HD are caused by neuronal loss in the brain, and PD is known to involve a specific local loss of dopaminergic neurons in the substantia nigra of the brain[13]. In fact, AD is the most common neurodegenerative disease, usually chronic and progressive, showing a decline in intellectual function, such as memory, judgment, and language skills, and impairments in daily life ability, personality, and behavior. HD is also accompanied by abnormal behavioral movements and cognitive impairment. PD is a degenerative neurological disease with the second highest incidence after Alzheimer’s disease, which is chronically manifested, as well as movement disorders such as tremors or paralysis of the arms, legs, and face, stiffness, stiffness, and postural instability, as well as pain, depression, and dementia. Although there are limited treatment options, the viability of cell therapy treatment has been the focus of recent research.
Stem cells were discovered in the early 1960s and are generally capable of continuous self-renewal and have the ability to differentiate into several types of cell lineages[14,15]. Stem cells include ESCs, progenitor cells, mesenchymal stem cells (MSCs), and PSCs, and are classified as totipotent, pluripotent, or multipotent according to their differentiation ability. Totipotent stem cells such as ESCs and PSCs can be isolated from the four cell stages of the embryo and can differentiate into all types of cells in the body, including tissues outside the embryo. Multipotent stem cells such as MSCs and progenitor cells can be isolated from various tissues in the adult human body and can differentiate into various cells, but only those of a closely related family type of cells. In recent years, the development of stem cell technology has expanded to many human body tissues, including treatment for degenerative neurological diseases using stem cells. The application fields of stem cells used in the treatment of neurodegenerative diseases are shown in Figure 1[16-23]. Because organizations with ineffective recovery systems cannot easily return after injury or extensive degenerative events, it is important to understand the characteristics of the available stem cell type and the specific mechanisms of neurodegenerative diseases, including AD, PD, and HD[12].
Stem cells and AD
The 2018 Global Alzheimer's Disease Report stated that 50 million people worldwide have the disease, and it is the most common cause of dementia, accounting for 50%-70% of cases of dementia cases[24,25]. AD has been shown to cause intracellular formation of nerve fiber tangles caused by the deposition of β-amyloid (Aβ) peptides on the extracellular matrix between neurons and the accumulation of hyperphosphorylated tau proteins in neurons[26].
MSCs play a major role in the treatment of AD, such as immune regulation, reduction of Aβ plaque burden through internalization and Aβ degradation of endosomal–lysosomal pathway oligomers and neurotrophic/regenerative potential[25,27]. Injection of green fluorescent protein (GFP)-labeled bone marrow (BM) MSCs in the hippocampus of an AD animal model has been shown to reduce the size of Aβ plaques and regulate functional immunity[28]. Transplantation of MSCs was shown to increase neurogenesis as demonstrated by immunostaining brain sections with an antipolysialylated form of the neural cell adhesion molecule and doublecortin antibodies[29]. It was also confirmed that MSCs labeled with PKH26-111 were injected into AD mice through the tail vein to reach the brain, and the radioactivity of BMSCs was significantly higher in the AD model than in the control group in the gamma counter and gamma camera imaging[30]. The effect of intravenous injection of BM MSCs in a mouse model of Alzheimer's disease was confirmed through β-amyloid positron emission tomography imaging, memory function studies, and histopathological evaluation[31]. Another technique for tissue repair involves paracrine effects using the secretion of extracellular vesicles from MSCs. The secretion of MSC extracellular vesicles can target Aβ deposition and is being studied as an important method for AD treatment, including siRNA and enzymes[29,32,33]. MSC-derived cytokines and vascular endothelial growth factors also showed regenerative effects in an AD model[34].
The mammalian brain has the capacity to repair itself through neurogenesis and gliogenesis to a limited degree; however, endogenous neurogenesis and gliogenesis decrease significantly with age and are unable to regenerate enough brain cells alone. Research using NSCs that express a phenotype similar to that of brain cells has great potential in the treatment of AD. Several recent studies have shown that NSCs can increase the survival and regeneration of endogenous neurons by producing neurotrophic factors, vascular endothelial growth factor (VEGF), and vessel density in the cortex[35], and that NSC-derived cholinergic neuron-like cells can also support a significant improvement in learning and memory ability with choline acetyltransferase (ChAT) activity[36]. Using a human source, NSC function was confirmed to significantly reduce cerebral Aβ42 Levels[37]. Parket al[38] found that a human NSC line encoding the ChAT gene was also transplanted into the amyloid precursor protein (APP) swe/PS1dE9 AD model mice and induced the proliferation of endogenous NSCs and the production of growth factors and neurotrophic factors.
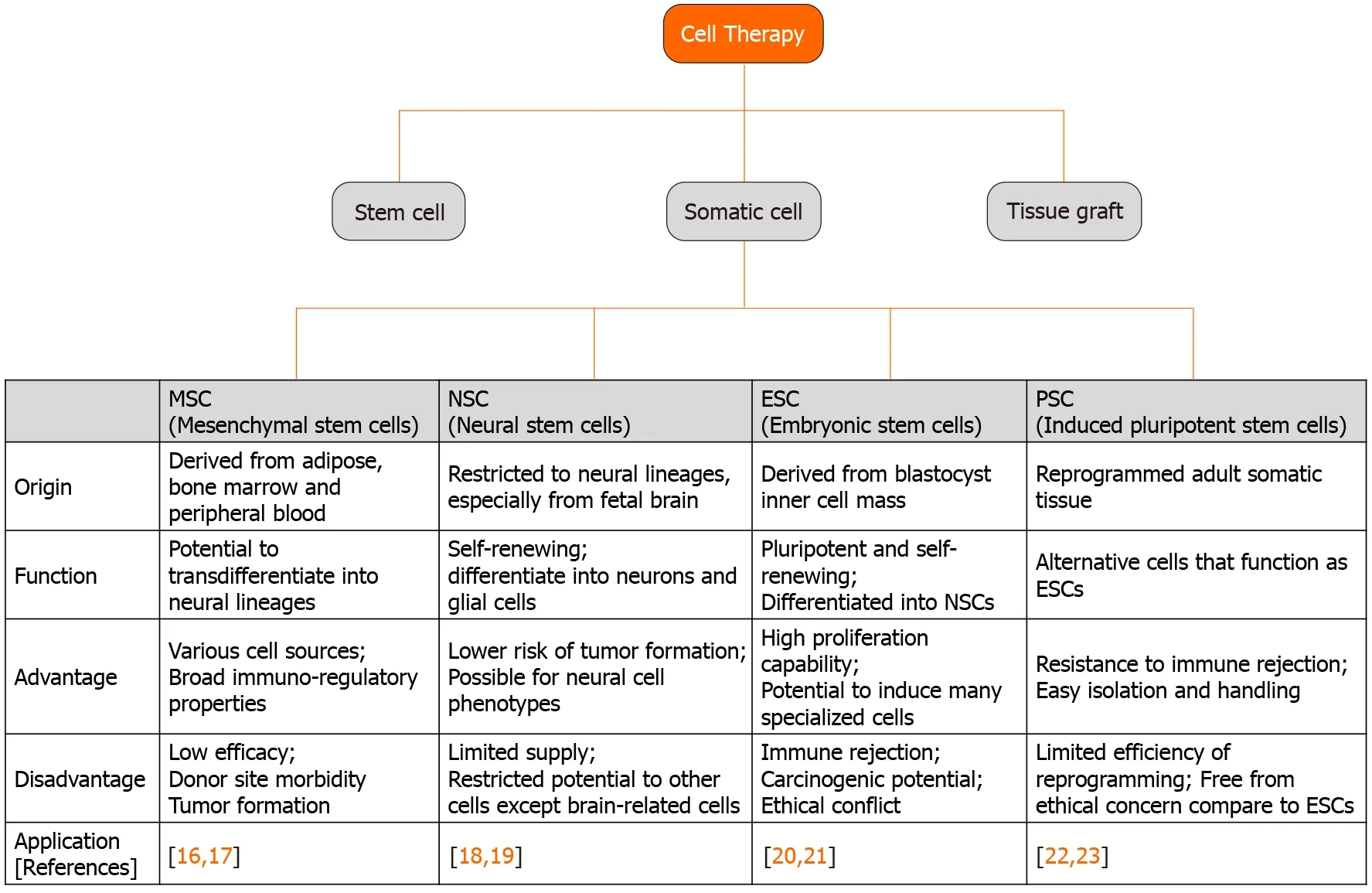
Figure 1 Stem cell types and their application to neurodegenerative diseases.
Significant experimental and clinical progress has been made with PSCs since they were discovered 10 years ago. They are now widely used in the treatment of AD to regulate endogenous neurogenesis, neuronal loss, and pathological changes. Administration of PSCs derived from mouse skin fibroblasts by treating protein extracts of ESCs has been shown to mitigate plaque deposition and cognitive dysfunction in a 5XFAD transgenic mouse model[39]. From human sources, PSC-or PSC-derived cells have been used to ameliorate degenerative disorders. Human iPSC-derived macrophage-like cells genetically modified to express neprilysin-2 or to mutate Tau Ex10+16, Aβ-degrading activity, differentiated into functional neurons, and reduced A β levels after xenograft administration to the 5XFAD or APP PS1 tg/wt NOD-SCID transgenic AD mouse model[40,41].
Stem cells and PD
PD is a common neurodegenerative disease characterized by impaired motor function, which is known to be caused by the selective loss of dopamine (DA) neurons in the human midbrain. Various studies have been conducted extensively on both motor and non-motor deficits. Cognitive impairment begins to develop motor impairment at an early stage and continues to progress. Non-motor symptoms are also a cause of deterioration in the quality of life of patients and treatments that can resolve cognitive impairment and dysfunction may be possible. Stem cells are generally used to consider neuroprotection, neuroplasticity, and immunomodulatory properties in PD pathogenesis.
Transplantation of human MSCs into 6-hydroxydopamine (6-OHDA)-induced lesions protected dopaminergic neurons and induced neurogenesis, resulting in therapeutic effects due to the release of soluble factors such as brain-derived neurotrophic factor (BDNF)[42]. MSCs were also found to regulate DA neuron apoptosis and oxidative stress and to improve motor function in the early stages of PD[43]. Intravenous administration of MSCs improved dysfunction and protected tyrosine hydroxylase-positive fibers in the striatum and substantia nigra pars compacta. It has been shown that the cytokine stromal cell-derived factor (SDF)-1α increases dopamine release from cells by inhibiting apoptosis in PC12 cells exposed to 6-OHDA, and confirmed that it is a principal component of the MSC-derived secre-tome[44]. To evaluate the effects of stem cell homing, Cerriet al[45] examined the intra-arterial infusion of MSCs in collaboration with a transient blood-brain barrier disruption by mannitol pretreatment. After 28 d of 6-OHDA induction, the progression of the damage site did not change; however, normalization of the pathological responsiveness of striatal neurons to dopaminergic stimulation was induced by MSC infusion. Furthermore, secretomes or exosomes derived from MSCs have been confirmed to have potential use in stem cells, including neuroprotective factors[46]. Conversely, the inhibition of cell-to-cell transmission molecules, namely α-synuclein, in MSCs led to functional improvement of motor deficits based on a pro-survival effect on cortical and dopaminergic neurons[47].
NSC transplantation allowed parkinsonian rats to be recovered through the regulation of SDF-1/chemokine receptor 4 (CXCR4) expression. Intraperitoneal injection of the CXCR4 antagonist, AMD3100, increased mRNA and protein expression of SDF-1 and CXCR4 in the NSC-transplanted site of the right substantia nigra. Furthermore, apomorphine-induced rotational behavior was reduced significantly in a rat model of PD[48]. In the xenograft model, the characterization of PD sites was examined using a high-throughput quantitative proteomic approach at the SN, striatum, olfactory bulb, and SVZ after human NSC treatment. These effects demonstrated that the rescue of SVZ function and the elicitation of endogenous response were induced by an increase in neurotrophic factors[49]. For three-dimensional microengineered cell therapy, NSCs were cultivated in the Nichoids microscaffold, and then transplanted into lesions of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Mechanical reprogramming of NSCs and recovery of PD symptoms produced an enhanced therapeutic effect in a murine experimental model of PD[50].
In a study conducted 20 years ago, transplantation of low-dose undifferentiated mouse ESCs into mice increased the proliferation of differentiated DA neurons and restored cerebral function in a PD animal model[51]. Another study demonstrated that a highly enriched population of midbrain neural stem cells derived from ESCs improved the electrophysiological and behavioral properties in a rodent model of PD[52,53]. Recently, studies on ESC-derived DA neurons capable of translational use have been actively conducted. Using a two-step WNT signaling activation strategy, human ESCs were induced to midbrain DA neurons at the clinical-grade level, and engraftment of these cells upregulated their behavioral recovery of amphetamineinduced rotation in a 6-OHDA model[54]. Furthermore, clinical-grade midbrain DA neurons, named MSK-DA01, safely demonstrated survival of the transplanted cells and behavioral amelioration in parkinsonian rats under GLP conditions without adverse effects[55]. PSCs were also examined for the improvement of PD at the preclinical level. In a primate PD model, hPSC-derived DA neurons showed an improvement in long-term survival of cells and spontaneous movement, dopaminergic progenitors derived from a clinical-grade human PSC line were produced, and their therapeutic effects were confirmed in 6-OHDA-lesioned rats[56,57].
Stem cells and HD
HD is a common degenerative brain disease with autosomal dominant inheritance. It is the least researched of the three major neurodegenerative diseases[58]. HD, characterized by progressive neuronal death, has various symptoms such as cognitive decline, behavioral changes, motor dysfunction, weight loss, sleep disturbance, and mental disorders[59]. This situation begins in the striatal part of the basal ganglia by increasing the number of CAG repeats in exon 1 of the huntingtin (HTT) gene, which encodes the huntingtin protein, leading to an atypically long polyglutamine region at the protein N-terminus[60]. Numerous therapies are aimed at slowing disease symptoms; however, stem cell therapy has been continuously studied by many researchers to restore the atrophied tissue, which can replace some abilities of degenerated cells[61].
MSCs are a promising HD treatment because they are not only simple to acquire and cultivate, but also have unique nutritive activity and immunomodulatory functions. Simple treatment of human MSCs has been demonstrated to enhance neural differentiation capacity, neurotrophic factor stimulation, and anti-apoptotic effects using the R6/2-J2 animal model. Transplanted MSCs can integrate with host cells to increase the level of secretory factors such as von Willebrand factor, SDF-1, and CXCR4[62]. Both intranasal deliveries showed the possibility of improving the therapeutic efficacy of HD. MSCs pre-treated with lithium and valproic acid (VPA) enhanced that open motor function improved walking distance and average speed in N171-82Q transgenic mice. Lithium and VPA have been used as mood stabilizers to boost cell survival and efficacy[63]. The second study demonstrated that MSCs significantly increased the survival rate of R6/2 mice, simultaneously with improved sleep disturbances and motor performance. The PoC of MSCs was explained by detecting GFP-expressing cells in the olfactory bulb, midbrain, and striatum on day 5. Furthermore, increased expression of DARPP-32 and decreased expression of inflammatory modulators were found in the striatum for 7.5 wk[64]. Treatment methods using various molecules released from MSCs have also been proposed. MSC-secreted neurotrophic factors were injected with quinoline acid (QA), which functioned as excitotoxicity in the striatum, and changed striatal volume with QA lesion of the brain in the rat HD model[65]. Two other studies demonstrated that MSCs that releaseHTT142mutant and shRNA or aquaporin 4 antisense oligonucleotides can expand the therapeutic abilities for HD therapy[66,67]. Recently, MSC-conditioned medium (CM) derived from the amniotic membrane has been reported to have a protective effect on striatal degeneration and motor deficits in the R6/2 mouse HD model. In this recovery process, a significant decrease in microglial activation and nitric oxide synthase induced by MSC-CM was observed, resulting in the modulation of inflammatory cells[68].
Pluripotent cell lines targeting HD have been developed by several research groups. The QA-lesioned HD rat model was monitored using an apomorphine-induced behavioral test and immunohistochemical staining after implantation of BDNF-overexpressing human NSCs (HB1.F3.BDNF) on the opposite side of the striatum. NSC PoC confirmed that the transplanted cells were moved to the QA lesion site with striatal GABAergic medium spiny neurons, containing DARPP-32 in the host brain[69]. A clonal conditionally immortalized NSC line (CTX0E03), which already showed safety and efficacy signals in patients with chronic ischemic stroke, was examined using the QA-lesioned HD model. Thirteen weeks post-transplantation, CTX0E03 survived in the striatum and cortex of the brain with QA lesions, differentiated into striatal neurons, and showed progenitor-palatal connections with the host tissue. Survived CTX0E03 reduced gliosis and host immune responses, but increased endogenous neurogenesis and angiogenesis[70]. The same research group also demonstrated the therapeutic potential of PSCs in a rodent model of HD. NSCs derived from a human PSC line (1231A3-NPCs) also showed reconstruction of the damaged neuronal connections and behavioral improvement for 12 wk post- transplantation[71]. Human embryonic stem cell-derived NSC lines were also reported in a therapeutic study in the striatum of R6/2 mouse HD fragment model (first confirmation) to confirm the efficacy of improving motor deficits and rescue synaptic alterations. The second confirmation for improving motor and late cognitive impairment was done using the Q140 knock-in mouse HD model[72]. Combination therapy related to transduction ofHTTgene-regulated PSCs was also conducted. PSCs derived from fibroblast/dental pulp of wild or HD rhesus monkeys were transfected with shRNA targeting theHTTtranscript and transplanted into the N171-82Q mouse model. The mutantHTT-PSCtransplanted group was encouraged in their lifespan counterpart, with motor function and pathological changes, including integration and differentiation[73].
STEM CELL TRACING IN DEGENERATIVE BRAINS
Histological analysis
To track transplanted stem cells, many preclinical studies on brain injury use bromodeoxyuridine, PKH26, and 4,6-diamidino-2-phenylindole as fluorophores[74-76]. Stem cells pre-labeled with fluorophores can be identifiedviaimmunohistochemistry in fixed tissue using anti-fluorescent-tagged antibodies or staining methods that use color-changing substrates. At present, histology is the gold standard to test whether transplanted stem cells survive or differentiate into tissue cells in an animal model study[77]. However, this requires the sacrifice of numerous animals and provides no longitudinal or whole-body monitoring. With a lack of information on stem cell behavior,in vivolongitudinal, non-invasive, and repeatable methods have been developed to monitor transplanted cells. In addition, it is crucial to track the capabilities of transplanted stem cells to reconstruct brain functions and biological roles.
Imaging technology
Stem cell imaging methods can be divided into direct and indirect cell labeling depending on the possibility of re-imaging over a long period of time. Direct cell labeling is the most frequently used method and consists of incubation prior to implantation and labeling cellsin vitrousing reporter probes containing fluorophores, radiotracers, or paramagnetic nanoparticles (Figure 2)[78,79-81]. These reporter probes can bind to specific epitopes on the cell membrane, such as copper-64-labeled antibody or zirconium-89-desferrioxamine-NCS (89Zr-DBN)[82,83], or can be absorbed by passive diffusion or transporters such as indium-111- and 89Zr-oxine or 2-[18F]-fluoro-2-deoxyglucose (18F-FDG)[30,79,84]. After incubation, cells are injectedin vivofor monitoring by magnetic resonance imaging (MRI), positron emission tomography (PET), single photon emission computed tomography (SPECT), and optical imaging.
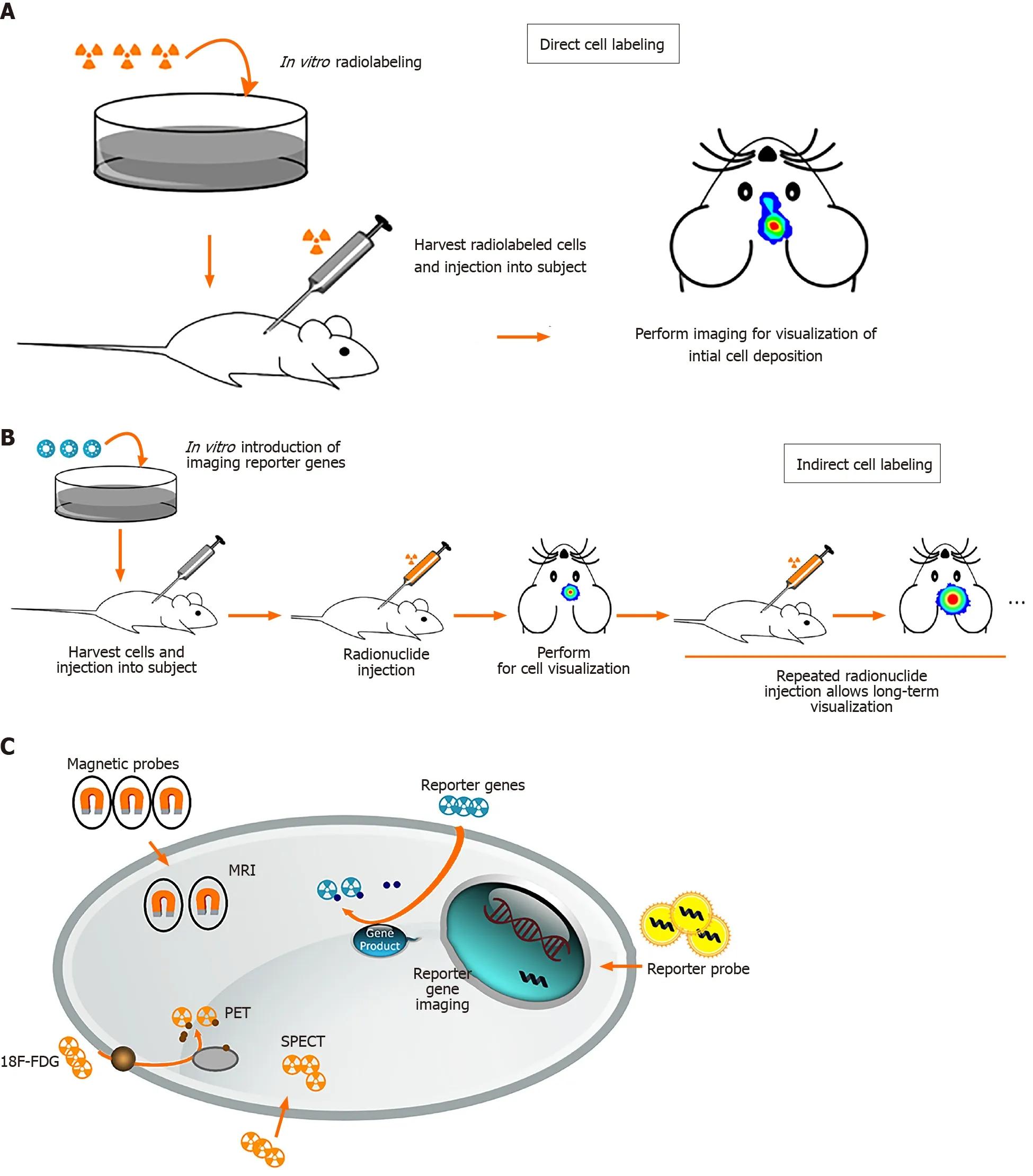
Figure 2 Schematic overview of different methods for stem cell labeling.
The first study of MR tracking of progenitor cells transplanted into the central nervous system was reported in 1992, and a superparamagnetic contrast agent was used for imaging rat brain cells[85]. Direct tracking through MRI offers benefits such as morphological characterization, high spatial resolution, lack of radiation, and longterm stem cell monitoring[86,87]. MRI requires the use of a contrast agent to visualize cells. For example, superparamagnetic iron oxide nanoparticles (SPIONs) have been shown to allowin vivomaintenance of neural progenitor cell viability, phenotype, proliferation, and differentiation[88,89]. Successful labeling of MSCs and progenitor cells with SPION has also been demonstrated in long-term, multimodal imaging and found no consequences on viability, differentiation capacity, or biological characteristics[90-92].
However, there are two limitations in labeling stem cells with magnetic contrast agents. After transplantation, the label was diluted because the stem cells continued to proliferate rapidly. Therefore, the MR signal decreased and lost rapidly over time because of cellular proliferation. In addition, SPIONs can be deposited in extracellular tissues when dead transplanted cells are entrapped by immune cells such as microglia in the central nervous system, leading to false signals in MRI[93].
PET and SPECT are nuclear medicine imaging techniques that represent promising imaging modality for tracking stem cells widely used in experimental trials. Before stem cells were transplanted into the host, radiotracers such as18F-FDG, lipophilic 99mTc-D,L-hexamethylene-propyleneamine oxime, and111In-oxine are required to label the stem cells to detect the transplanted cellsviaPET or SPECT scanner[79,94-96]. There was no difference in viability or differentiation ability after labeling with radiotracers. No microstructural changes were observed. The positron emitted from the radioactive isotope rapidly loses kinetic energy while traveling through the surrounding tissue, and then interacts with the electron to emit two high-energy photons of 511keVat (high-frequency photons) moving in almost the opposite direction. PET camera scanners can detect and image these photons. SPECT is very similar to PET in the use of radioactive tracers and the detection of gamma rays. These methods provide sensitivity in the picomolar range and the ability to use the same tracer across multiple species. SPECT imaging has the added advantage of having a lower false-positive signal compared to MRI. However, they do not provide anatomical information and must be used in conjunction with MRI, computerized tomography, or X-rays[97].
Optical imaging, compared to MRI, PET, and SPET, has the benefits of lower cost, rapid acquisition, no radiation toxicity, and relatively high sensitivity[98]. Semiconductor nanocrystals, also known as quantum dots (QDs), are a new class of biocompatible fluorescent materials that are relatively photostable and have a narrow luminescence band used for cell tracking. Near-infrared-emitting QDs may be particularly useful for tracking transplanted cells in the human brain, because longer wavelengths allow easier penetration of tissues such as bone and skin[99]. Bioluminescence imaging (BLI) has been widely applied in preclinical studies of stem cell imaging in the brain for several years. BLI has also been used to quantify gene expression and stem cell localization in mice and rats[100,101]. BLI is only limited to small animals, but not to large animals, because BLI can only penetrate a few centimeters of tissue.
Indirect cell labeling was modified by inserting an exogenous reporter gene into the cells. These reporter genes can produce specific proteins that function as radioactive probes, so the probe signal is not limited to the half-life of the tracer used and can be detected by PET, SPECT, and MRI for a long time. It not only allows long-term noninvasive imaging of stem cells, but also distinguishes between viable and nonviable cells. The most widely used imaging reporter gene is luciferase from firefly Photinus pyralis (Fluc), which catalyzes the oxidation of D-luciferin to oxyluciferin by emitting visible light (the principle of BLI) in the presence of oxygen, magnesium, and adenosine triphosphate. Integration of Fluc into NSCs and MSCs has been successfully performed for cell monitoring using BLI[102-105]. Another example of an indirect method of tracking consists of an exogenous reporter gene delivered to cells of interest, such as a ferritin heavy chain, which can act as a powerful MRI reporter to track cell distribution and migration in acute ischemic stroke[106]. However, the major drawback of reporter gene-based imaging is the incorporation of genomic material into the cell.
THREE-DIMENSIONAL IMAGING FOR STEM CELL TRACKING
Tissue optical clearing technique
Tissue imaging techniques for the depiction of three-dimensional structures and their molecular information are a growing trend that researchers need to facilitate volumetric imaging rather than the two-dimensional section used as the standard procedure. Due to the three-dimensional scalability of individual neurons and their interrelationships with brain cells, this imaging technique, namely Clear, Lipidexchanged, Acrylamide-hybridized Rigid, Imaging/immunostaining compatible, Tissue hYdrogel (CLARITY), was developed by the Stanford University Research Group in 2013[107]. According to the tissue-reagent reaction principle, three major techniques are now available: hydrogel-, solvent- (hydrophobic), and aqueous-based (hydrophilic) approaches[108]. Descriptions of these techniques are provided in Table 1[107,109-113].
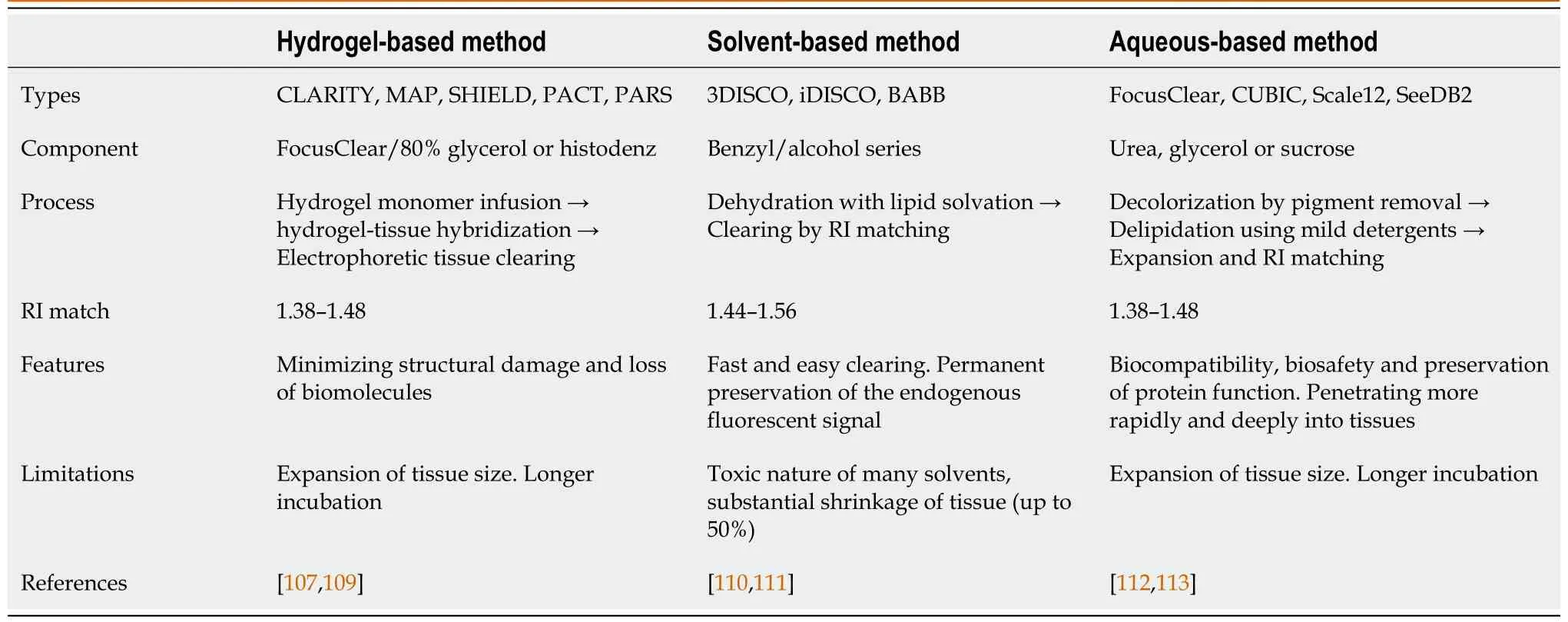
Table 1 Comparison of major clearing techniques
Intact tissue clearing methods continue to grow for three-dimensional imaging of the brain, centered on labeling options and imaging analysis tools. It is expected that this process may prove the discovery of novel physiological and pathological mechanisms based on three-dimensional molecular information for neurodegenerative diseases. In the beginning, disconnected axons with APP accumulation and swelling were found in the traumatic mouse brain, revealing novel insights into threedimensional axon degeneration of temporal progression after axonal injury[114]. To identify specific brain regions with early susceptibility to AD progression, Canteret al[115] created a spatiotemporal map of Aβ deposition by using whole-brain systemwide control of interaction time and kinetics of chemicals immunolabeling in the 5XFAD model, suggesting an understanding of the mechanisms of brain dysfunction and progressive memory loss[115]. Furthermore, neurons and mitochondrial proteins in the cerebellum of mouse and human brain tissues were first optimized by revealing mitochondrial disease. It has been demonstrated in a three-dimensional network that vascular, dendritic, or axonal networks finely determine the interrelationships between complex vascular structures and vasogenic factors in patients with mitochondrial disease[116].
Challenges and approaches for stem cell PoC
There are no PoC studies of three-dimensional stem cell tracing for treatment of neurodegenerative diseases. The only research on the existence of stem cells represented the spatial relationship with endogenous Gli1 positive MSCs in adult calvarial bones during postnatal craniofacial development, and indicated the osteogenesis mechanism for craniofacial research using the bone specific poly (ethylene glycol)- associated solvent system tissue clearing method[117].
The identification of the transplanted stem cells that can participate in the specific circuit and the host neurons that provide inputs to them may be critical for successful cell tracing for stem cell-based therapies for neurological disorders. To trace full or limited area projections in the brain, researchers need to be complemented by labeling or genetic manipulationin vivobefore stem cell transplantation and by using highresolution image system includingin vivomulti-photon or light-sheet microscopy[118]. Neuro-specific proteins, DNA/RNA-conjugated fluorescent dyes, and viral/non-viral constructs have been used to explore the connectivity between reciprocal hosts and stem cell grafts[119]. For reliable three-dimensional analysis, the membrane-bound protein-specific phenotype of stem cells and the target circuits with strong can be set and verified in a genetic animal model of neurodegenerative disease[120,121]. Furthermore, stem cell labeling based on gene delivery can be important to define the correlation analysis in three dimensions between PoC phenotypes of endogenous or exogenous stem cells, state-modified/unmodified proteins, and statealtering genes to understand the physiology and pathology of degenerative brains[108,122].
CONCLUSION
For many decades, appropriate cell tracing strategies for PoC and the connectivity between host neurons and grafted stem cells have been observed using traditional two-dimensional tracing techniques. Through the development of tissue optical clearing techniques and their convergence technologies, however, it is possible to demonstrate tracing in three dimensions and to analyze the molecular pathological changes associated with endogenous cells functioning in neurodegenerative diseases. Studies on the ability of three-dimensional host-graft integration in diseases will help to serve from the basic application to the clinical monitoring of the potential strategies of stem cell therapy. An understanding of the three-dimensional imaging of stem cells may also help to approach fundamental questions regarding the cell conditions, that is, dose, time, phase, and disease mechanism, when regenerating naturally or therapeutically in neurodegenerative disease.
杂志排行

World Journal of Stem Cells的其它文章
- Effects of living and metabolically inactive mesenchymal stromal cells and their derivatives on monocytes and macrophages
- Stem cells' centrosomes: How can organelles identified 130 years ago contribute to the future of regenerative medicine?
- Effects of storage media, supplements and cryopreservation methods on quality of stem cells
- Stem cell therapies in cardiac diseases: Current status and future possibilities
- Current evidence on potential of adipose derived stem cells to enhance bone regeneration and future projection
- Neural stem cell therapy for brain disease