微藻碳减排与生物质利用技术研究进展
2021-10-11荣峻峰宗保宁
李 煦,荣峻峰,宗保宁
(中国石化石油化工科学研究院,北京 100083)
“能源”和“环境”是人类社会可持续发展面临的重大问题,工业化和人类活动造成温室气体在大气中的累积是引起气候变化的一个重要因素[1]。化石能源的使用一方面支撑了人类社会的高速发展,另一方面也造成了严重的环境污染和碳排放危机。2018年全球能源消耗比上年度增长2.3%,仅使用化石燃料排放的CO2就达到了33.1 Gt[2]。大气中CO2质量分数也从1980 年的340 μg/g上升到2020年的412 μg/g[3]。CO2约占温室气体总量的50%,但对环境变化所产生影响的比例却达到了82%,带来了包括海平面上升、食品安全危机、物种灭绝与生物多样性危机等诸多问题[4]。国际社会也在不遗余力地制定各种政策,希望能够减缓CO2排放量逐年上升而引起的全球变暖趋势。在技术方面,CO2的减排、封存与利用成为降低CO2净排放量的有效手段。
光合作用是生物利用光能将CO2吸收并转化为有机物的过程,是自然界经过数十亿年进化形成的高效CO2固定手段。微藻是能够进行光合作用的单细胞或细胞聚集体生物,根据其结构的不同可分为原核微藻与真核微藻。原核微藻以蓝藻为主,又称为蓝细菌,包括螺旋藻、颤藻、念珠藻等种类。原核微藻细胞内含有光合色素,但不形成叶绿体,细胞结构与革兰氏阴性细菌相近。真核微藻具有细胞核、叶绿体等细胞器,包括绿藻、硅藻、轮藻等诸多种类[5-6]。微藻通常生活在水环境中,不具有组织与器官的分化能力。与陆生高等植物相比,微藻通过光合作用进行CO2的吸收有其独特的优势。首先,微藻的生长速率远高于高等植物。由于微藻细胞不产生分化,理论上所有的细胞都可以进行光合作用生成有机物,因此微藻生物质积累的速率远高于高等植物,生物质产率可达50~100 t/(ha·a)[7]。其次,微藻以细胞为单位进行营养物质的吸收与利用,不存在远距离的运输过程,因此固碳效率高于依赖维管束系统进行物质运输的高等植物。最后,微藻种类繁多,可以在淡水、海水等多种条件下生长,扩大了微藻固碳技术的适应性。以下综述微藻固碳领域近年来的研究与实践进展,包括微藻碳固定过程与微藻生物质的利用等方面,以期为今后的技术开发与应用提供思路。
1 微藻固碳的原理

1.1 光能捕捉与传递
光系统是微藻进行光吸收的功能单位,是由色素、脂肪和蛋白质组成的复合物。每一个光系统含有捕光复合物和光反应中心复合物两个主要成分。捕光复合物中的光吸收色素可形象地比喻为一种天线,捕获光子并将能量传递给反应中心的色素,光反应中心复合物利用这一能量裂解水分子,形成O2与H+,并激发出一个自由电子进入光合电子传递链,这一过程又称为水的光解[8]。H+的不断产生形成了跨生物膜H+浓度梯度。ATP(腺苷三磷酸)合成酶在跨膜H+浓度梯度的驱动下形成ATP,作为后续有机物合成的能源;而自由电子则经过光系统中复杂的转递后与NADP+(氧化型烟酰胺腺嘌呤二核苷酸磷酸,氧化型辅酶Ⅱ)形成NADPH(还原型烟酰胺腺嘌呤二核苷酸磷酸,还原型辅酶Ⅱ),作为后续有机物合成过程中还原力的来源[9]。
1.2 无机碳浓缩机制
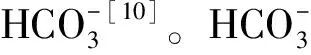
1.3 光合碳代谢
CO2在核糖-1,5-二磷酸羧化酶(Rubisco酶)的催化下与核糖-1,5-二磷酸生成3-磷酸甘油酸,是无机碳固定成为有机碳的第一步。以光能捕捉与传递步骤中产生的ATP与NADPH为能量和还原力的来源,3-磷酸甘油酸经过多步酶催化生成果糖-1,6-二磷酸,随后转变为蔗糖、淀粉等糖类分子,以生物质的形式储存下来,最终实现CO2到生物质的转化。Rubisco酶是叶绿体中含量最高的蛋白质,可能也是地球上最丰富的蛋白质,是光合作用碳固定的核心,也是受到严密调控的位点[13]。
2 影响微藻固碳效率的因素
2.1 藻种
以CO2为原料合成有机物的过程发生在微藻细胞中,不同种类的微藻固定CO2的效率也存在差异。研究者估计地球上存在约30万种微藻,而被人们发现并记录的仅有约3万种,不同微藻物种之间的差异远大于高等植物之间的差异[14]。因此选择合适的微藻藻种就成为提高固碳效率的首要手段。性状优良、CO2吸收固定效率高的藻种既可以从自然界筛选得到,也可以通过基因工程等手段对天然藻种进行改造获得。
2.1.1 藻种筛选微藻在天然水环境中生长,对于空气的CO2浓度有较好的适应性。然而工业烟气中的CO2浓度远高于空气,在利用微藻进行烟气CO2减排时需要筛选能够耐受高浓度CO2并快速生长的藻种。Zhao Bingtao等[15]使用体积分数15%的CO2进行了微藻养殖试验,结果显示小球藻(Chlorellasp.)的效果优于等鞭金藻(Isochrysissp.)与强壮前沟藻(Amphidiniumcarterae),小球藻生物质积累效率可达到0.192 g/(L·d),CO2固定速率为0.353 g/(L·d)。Radmann等[16]评价了不同藻种在以电厂烟气CO2为碳源的养殖条件下的生长情况,结果显示螺旋藻、普通小球藻、斜生栅藻与聚球藻的生物质质量浓度分别可达到1.59,0.98,0.68,0.41 g/L。不同藻种在CO2固定方面具有较大的差异,因此需要针对待减排的烟气特征筛选合适的藻种。
2.1.2 藻种改良定向进化是一种以环境条件作为选择压,模拟自然界进化过程实现生物体性状快速改良的手段。通过人为施加比自然界更为严格的筛选条件,能够大大提高有益突变积累的速率,从而在较短的时间内实现生物性状快速进化。Li Dengjin等[17]通过逐步提高CO2浓度进行培养,使小球藻逐渐适应了电厂烟气中CO2的高浓度,解除了CO2体积分数大于10%时对小球藻生长的抑制作用;进而发现,即使CO2体积分数高达30%,也不会对定向进化筛选所得藻种的生长产生明显的不利影响。Cheng Dujia等[18]通过46轮的筛选,获得了最大CO2固定速率达到1.20 g/(L·d)的藻种,这一结果远高于天然筛选得到的藻种。Aslam等[19]使用电厂烟气对混合微藻种群进行了定向进化,筛选得到了对CO2与NOx浓度较高的烟气具有良好耐受性的链带藻藻种。
随着对微藻生长与生物质积累以及光合作用研究的深入,这些过程的分子机制越来越明晰,分子生物学技术也正在日新月异地发展。这些都为通过合成生物学和基因工程手段对天然藻种进行改良、提高微藻固碳效率奠定了基础。通过藻种遗传改造提高微藻吸收CO2效率可能的途径,包括提高CO2固定效率、增强光能捕捉系统以及引入新的CO2固定通路以减少能量损失。微藻细胞固定CO2的过程有两个关键环节,分别为Calvin循环与碳浓缩机制(CCM)。Hazra等[20]通过构建杂合的Rubisco酶提高了莱茵衣藻对CO2/O2的特异性。而过表达Rubisco活化酶则能够在不改变CO2浓度的条件下使微拟球藻的生物量产率提高46%[21]。提高景天庚酮糖-1,7-二磷酸酶的表达量能够改善多种藻类的生物质积累和光合作用效率[22-23]。而在普通小球藻中,过表达果糖1,6-二磷酸醛缩酶能够使光合作用速率提高1.2倍[24]。绿藻细胞中吸收光能的色素主要是叶绿素a和b,它们能够吸收的日光波长范围在400~700 nm之间。向细胞中引入其他色素能够扩大吸收光的波长范围,从而改善光合作用的能量供应,并更加充分地利用自然光。Tros等[25]在聚球藻PCC 7002中表达了叶绿素f合成相关基因,使细胞吸收光的波长上限提高到750 nm,从而使细胞吸收的光能提高了19%。
2.2 养殖装置
微藻生物质的组成与积累速率在很大程度上受到养殖条件的影响。微藻的规模养殖流程和光生物反应器形式是微藻生物技术的核心环节,也是目前该领域的技术瓶颈[26]。微藻的养殖系统根据具体形式的不同可分为开放式系统和封闭式系统两类[27]。目前,通过养殖微藻进行CO2减排时最常用的养殖装置是跑道池。这种开放式的养殖装置具有结构简单、可扩展性较好、成本低的优势;但由于混合程度低、光利用效率低且藻液直接与外界环境接触,对微藻生物量积累、CO2固定效率以及微藻品质都带来了不利影响[28-29]。
为了解决上述问题,研究者开发了封闭式光反应器用于微藻养殖。封闭的养殖系统隔绝了微藻与外界环境,避免微藻受到灰尘、细菌、原生动物等的污染,大大提高了微藻产品的品质。同时,封闭式光反应器可以通过在传质传热、补光导光等方面进行改进,提高CO2利用效率和微藻生长速率,从而改善微藻固定CO2的能力。更优秀的养殖系统意味着要在效率、成本、占地、能耗等多个因素中取得更好的平衡。众多研究者在微藻养殖系统形式方面进行探索,得到了微藻在封闭式反应器中的固碳效率,结果如表1所示,而这方面的探索还远没有结束。
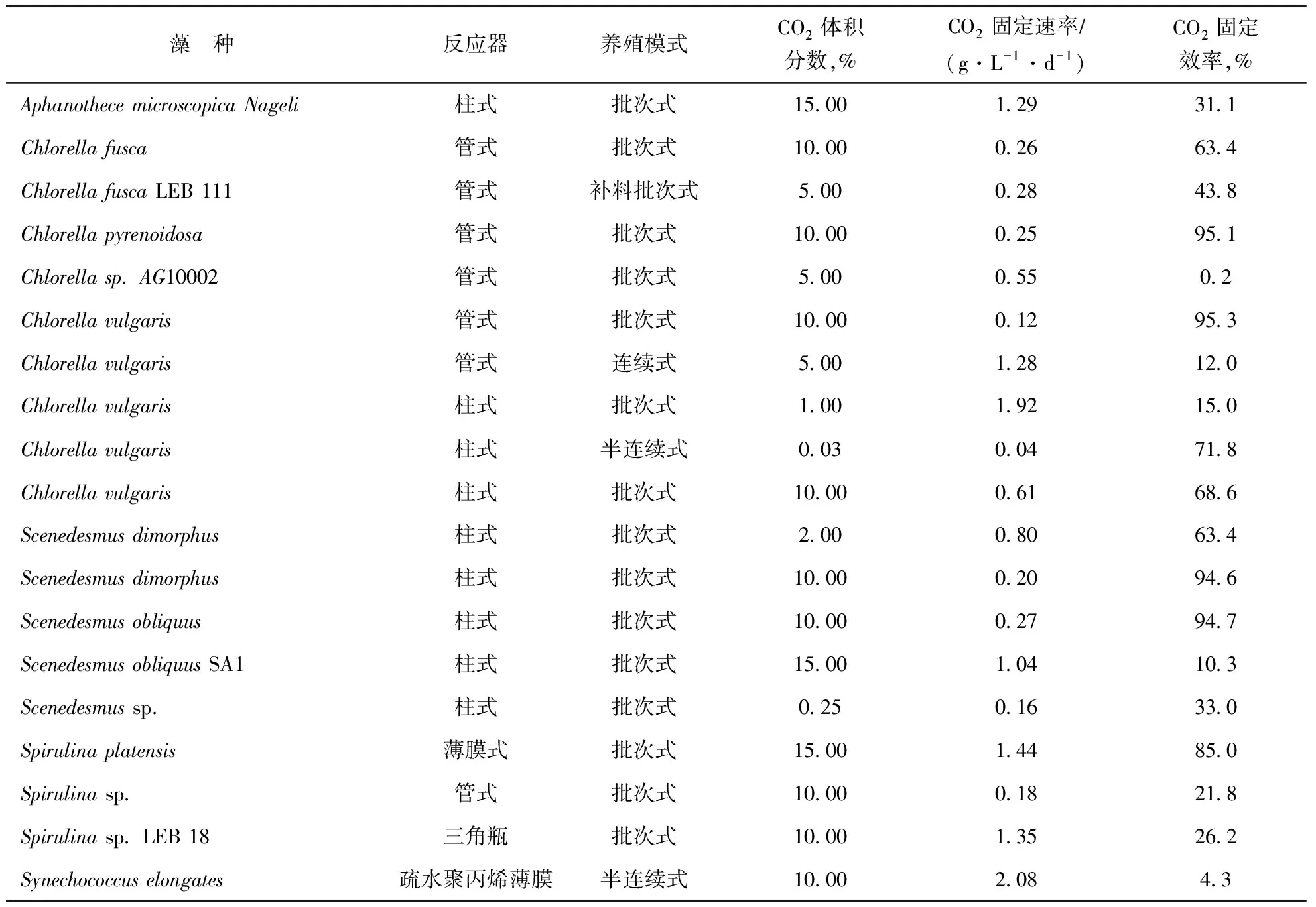
表1 微藻在封闭式反应器中的固碳效率[3]
2.3 养殖条件
2.3.1 光照光能是微藻光合作用生产有机物的能量来源,因此光照是微藻固碳的重要影响因素。总体上,微藻的生长速率随着光照强度的增强而提高[30]。当光强超过特定的光饱和点后,继续提高光强则会引起生长速率的下降,称为光抑制现象。这是由于过量光照产生的自由电子数量超过了碳固定反应的需求,产生了大量活性氧,对细胞造成了氧化损伤[31-32]。除光照强度外,光周期(一天中光照/黑暗的时间比例)也是影响微藻生长与生物质积累的重要因素,通常每天12~18 h的光照能够实现良好的效果[33]。但具体的光照参数根据实际的藻种与光照条件而又有所不同。例如,Wahidin等[34]发现:对于微拟球藻,在光量子密度为100 μmol/(m2·s)的中等光照条件下,光周期为18 h/6 h时可获得最好的效果;而在光量子密度为200 μmol/(m2·s)的高光强条件下,最佳的光周期改变为12 h/12 h。太阳光是光能最廉价的来源,大规模的微藻养殖目前也以利用自然光为主。但随着人工照明技术的发展,研究者也开发了用于微藻培养的人工光源,以提高微藻生长与固碳的效率。Koc等对发光二极管(LED)、卤素灯、荧光灯与白炽灯等多种光源进行了比较,结果显示采用了AlIn GaP技术的LED光源在促进微藻生长方面具有最好的效果与经济性[35]。光伏与人工光源技术相结合,有望提高微藻对能量的利用效率。
2.3.3 CO2浓度在传统的微藻养殖模式下,CO2在水中的溶解是一个较慢的过程,过高或过低的CO2浓度都会对微藻吸收CO2的效率产生影响。CO2浓度过低会使细胞不能得到足够的CO2;而CO2浓度过高一方面会由于溶解不完全而产生逸散,另一方面则会因为CO2在水中的解离导致培养体系pH过低[38]。因此,需要控制合适的CO2浓度,使CO2的供给与微藻对无机碳的需求相匹配,并维持合适的环境条件[39]。Razzak等[40]使用混有体积分数2%~12% CO2的空气进行小球藻的养殖,结果显示CO2体积分数为4%时微藻固碳效率最高,CO2吸收速率为0.2 g/(L·d)。在通入CO2体积分数为10%的空气培养时,斜生栅藻的比生长速率为(0.887±0.012) d-1,比CO2体积分数为20%时提高13.7%[41]。因此,在利用微藻进行CO2减排时,需要结合烟气种类与CO2浓度、藻种特性以及养殖条件,确定合适的烟气通入方式。
3 微藻生物质的利用
在绿色经济的背景下,资源的循环利用已经成为支撑人类社会可持续发展的重要力量。将固定CO2获得的微藻生物质进行高价值利用,就成为微藻固碳技术不可或缺的一环。微藻的生物特性具有多样化的特征,这就为微藻在不同领域的广泛利用提供了可能。
3.1 微藻生物质的组成
与其他生物体一样,微藻在生长过程中能够积累大量的蛋白质、糖类、脂肪、核酸及其他生物活性物质。不同的微藻具有不同的生物化学组成,而微藻养殖方式的不同也会对生物质组成产生影响。表2为典型藻种的生物质组成。从表2可

表2 不同微藻生物质的化学组成(以干物质计)[42] w,%
以看出,为了适应不同领域的应用需求,需要选择具有不同生物质组成特性的藻种,才能发挥出微藻生物质的最大价值。
3.2 微藻生物能源
脂肪在微藻细胞中主要作为能量储存物质和细胞结构的组成成分。其中,甘油三酯是细胞主要的能量储存形式,而由磷脂和糖脂构成的生物膜则是微藻细胞赖以存在的结构基础[43]。从微藻的总脂中获取脂肪酸,再通过酯化反应获得脂肪酸酯,就能够作为燃料加以利用。微藻细胞中的脂肪酸链长度通常在C12~C18之间,获得的脂肪酸酯性质与柴油相近,因此也被称为微藻生物柴油[44]。微藻总脂中含有较高比例的不饱和脂肪酸,这些不饱和的碳链会对生物柴油的品质产生影响,因此在生产过程中需要进行加氢处理,以改良油品品质[45]。除生物柴油外,微藻生物质还可以用于生产甲烷、合成气、氢气、乙醇、喷气燃料等多种能源产品[46-47]。
微藻生物柴油生产的第一步是从收获的微藻生物质中提取油脂。传统的油脂提取是利用相似相容原理,使用有机溶剂对干燥的微藻生物质进行萃取。极性溶剂(例如异丙醇、甲醇等)与非极性溶剂(例如己烷、氯仿等)都可以实现微藻油脂的提取,但两类溶剂混合使用通常能够得到更高的提取率[48]。有机溶剂的使用通常会带来环境与安全方面的风险,近年来研究者将超临界流体萃取技术应用于微藻油脂的提取,大大降低了溶剂的使用与泄漏带来的风险。超临界流体萃取技术具有较强的灵活性,且溶剂回收过程不需要专门提供能量,效率也较高。例如,使用超临界CO2进行微藻油脂的提取,总脂收率通常在60%以上,甚至可以高达98%~100%[49]。为了获得更高的总脂收率,除了提取压力、温度、时间等常规提取参数外,还有一些因素也有改良的空间。微藻生物质在提取前通常需要脱水,将原料水质量分数降低至20%以下能够显著提升提取效率[50]。通过超声、球磨或微波等手段在提取前对原料进行预处理,破坏微藻细胞壁,能够将提取率提高16%~20%[51]。由于CO2极性较低,在超临界CO2提取过程中加入少量极性溶剂乙醇,能够在不增加提取过程安全风险的前提下将提取率提高20%~90%[52-53]。
获得微藻总脂后,传统的生物柴油生产方式是将甘油三酯转化为游离脂肪酸,再用甲醇酯化获得脂肪酸甲酯。这种分步式的生产方式工艺较为复杂,各级提取纯化过程的成本也较高。研究者开发出原位转酯反应工艺,将脂肪提取与酯交换一步进行,使用甲醇同时作为提取溶剂和酯交换反应原料,从而大大降低了工艺的复杂程度和成本[54]。Patil等[55]对小球藻藻油进行了原位转酯反应,使用质量分数20%的硫酸作为催化剂,在60 ℃下反应4 h,脂肪酸甲酯收率最高可达98%。将超声处理与原位转酯反应相结合,可以使藻油生产生物柴油的得率达到98.9%[56]。
3.3 高蛋白微藻生物质
蛋白质是微藻的重要生化组成成分之一,在细胞结构组成、物质与能量代谢、信号传递与营养储存等方面都具有基础性的作用。某些微藻的蛋白质含量很高,例如蛋白核小球藻与螺旋藻等,生物质中蛋白质质量分数可以达到40%~70%,高于常见的蛋白质来源[45,57]。蛋白质的营养价值很大程度上取决于其氨基酸的组成,特别是其中必需氨基酸的含量。植物来源的蛋白质通常缺少一种或几种必需氨基酸,大大影响了其营养价值。而微藻蛋白质的氨基酸组成丰富,其营养价值也优于多种常见的动物蛋白来源(如畜肉、禽肉与奶制品等)[58]。根据世界卫生组织/联合国粮农组织的推荐,以小球藻和螺旋藻为代表的微藻生物质中必需氨基酸的含量非常均衡,能够满足人类膳食所需[59]。因此不管是作为人类食物还是作为水产畜禽饲料,高蛋白的微藻生物质都具有非常好的应用前景。
微藻作为饲料的研究开始于20世纪50年代。近年来,越来越多饲料中出现了微藻生物质的身影,包括大型牲畜饲料、禽类饲料与水产饲料等[60]。饲料中添加微藻,不仅能作为蛋白质的来源替代饲料中传统的蛋白质来源(例如豆粕、鱼粉等),还能够改善动物的生长性状,从而提高养殖业的收益。Holman等[61]在绵羊饲料中添加了质量分数为10%的螺旋藻,试验动物的平均体重出现了显著增长。螺旋藻完全替代饲料中的鱼粉不会降低鲤鱼的生长性能,且更有利于蛋白质的沉积[62]。螺旋藻替代50%的鱼粉能够显著提高西伯利亚鲟鱼的增重率和饲料转化率[63]。目前,我国已将裂壶藻、螺旋藻、拟微绿球藻与小球藻等列入了饲料原料目录,允许在饲料中使用上述藻种作为原料,为微藻在饲料领域的应用进行了规范。
在微藻作为食品方面,数十年以来也得到了长足的发展。美国FDA已经将小球藻、螺旋藻、杜氏藻、裂殖壶藻等多种微藻认定为公认安全的(GRAS)食品原料[64]。在螺旋藻最初的发现地非洲乍得湖,天然生长的螺旋藻本身就是当地人民传统的食物之一,食用螺旋藻已有数百年的历史。随着食品工业的发展,目前市场上也有多种添加了小球藻或螺旋藻成分的食品、饮料等出售,如面包、酸奶、冰激凌、饼干等,为追求营养和健康的消费者提供了新的选择[65-66]。
3.4 高价值微藻代谢物
除了脂肪、蛋白质与糖类等大量营养成分外,微藻还含有多种具有生物活性的代谢物,能够在保健品、医药产品与美容产品中开展应用。微藻中含有多种人体不能合成的长链多不饱和脂肪酸(PUFA),例如十八碳三烯酸(亚麻酸,ALA)、二十碳五烯酸(EPA)、二十二碳六烯酸(DHA)等。这些PUFA在人体内具有促进神经细胞发育、调节细胞功能、抗炎症、调节血糖血脂等多种生物活性,具有极高的经济价值。EPA和DHA的传统来源是深海鱼油。事实上,海鱼本身合成这些PUFA的能力较低,体内的EPA和DHA主要来源于食物链捕食,因此这些PUFA的最终来源正是海洋食物链的原初生产者——微藻。部分微藻细胞内含有高浓度的PUFA,例如微绿球藻和三角褐指藻总脂中EPA的质量分数可高达39%,而裂殖壶藻总脂中DHA的质量分数也可达到30%~40%[67]。通过微藻大规模培养与提取直接获得PUFA,不但降低了成本,还能够避免重金属等污染物经食物链在海鱼体内富集引起的安全风险,是非常理想的PUFA来源。
类胡萝卜素是一类长链的类异戊二烯聚合形成的萜类化合物。微藻作为光合生物,细胞内含有多种类胡萝卜素作为吸收光能的色素,例如β-胡萝卜素、叶黄素、虾青素、岩藻黄素等。这些类胡萝卜素分子中具有共轭双键,在人体内能够发挥视觉维持、抗氧化、抗炎症等多种功能,是微藻源保健品开发的热点。雨生红球藻(Haematococcuspluvialis)在其生长过程中的红色细胞阶段能大量积累虾青素,质量可达藻体干基质量的2%~5%,是理想的天然虾青素来源[68]。金色奥杜藻生物质中含有质量分数约2%的全反式岩藻黄素,具有很强的抗氧化功能[69]。
藻蓝蛋白是蓝藻中广泛存在的捕光色素蛋白,由脱辅基蛋白和作为辅基的开链线性四吡咯(藻胆色素)以硫醚键共价结合而成。藻蓝蛋白是天然的水溶性色素蛋白,作为一种食品色素添加剂被广泛应用于食品工业中[70]。藻蓝蛋白还是功能强大的广谱抗氧化剂,能够清除体内的自由基,具有抑制癌细胞、抗氧化、促进细胞再生、提高机体免疫力等功能,在化妆品与医药保健等行业具有应用前景[71]。螺旋藻是目前生产藻蓝蛋白的主要原料。尽管藻蓝蛋白的市场需求很大,但受提取纯化技术所限,目前产量还不能满足需求,成本也较高。研究者正在通过开发新的提取纯化技术提升藻蓝蛋白的生产能力,并降低成本,扩大藻蓝蛋白的市场规模[72]。
4 微藻固碳的经济性
微藻的人工养殖现在仍然是一个规模较小的产业,全球每年养殖的微藻总量仅为20~30 kt,且大部分微藻生物质用于食品与保健品等领域,生产成本与价格均较高[73]。例如,使用封闭式反应器生产的微藻,成本通常在1万美元/t以上,仅有保健品、医药产品、美容产品等少数应用途径能够接受如此高的成本。使用开放式养殖装置并结合CO2废气与含氮磷废水的应用,能够将微藻的生产成本降低到0.2~0.3万美元/t,具有在饲料等大宗蛋白领域实现商品化的潜力[74-75]。利用微藻固定CO2的成本受到原材料、能耗、土地占用等多个因素的影响,目前仍然处在较高的水平。微藻在饲料与环保产业中的应用也受限于较小的生产规模和产品产量[76]。为了提高微藻固碳的经济效益,势必要通过技术开发与大规模养殖的实践来降低微藻养殖的成本,并做大微藻生物经济的规模。目前全球微藻产业的市场总价值在5 000万美元左右,到2025年预计将增长至7 000万美元[7]。随着碳税、碳汇与碳交易制度的实施,CO2的排放成本将会越来越高,而CO2的吸收利用势必成为一项有利可图的工作。如果将微藻养殖与CO2减排相结合,则能够在很大程度上减少微藻养殖的原料成本;而大规模的微藻养殖也会使人工成本、管理成本等得到最大程度的降低,从而使微藻产业得到迅猛发展,并开辟更多新的应用途径,实现市场规模与应用领域的正反馈,使碳元素循环利用的新经济模式成为可能。
5 最新研究进展
微藻固碳作为生物技术、新能源与绿色环保等战略性新兴产业的交叉点,用自然的手段解决自然的问题,能够在循环经济的逻辑上实现环境保护和可持续发展。中国石化石油化工科学研究院(简称石科院)在微藻环保减排领域开展了十余年的工作。石科院微藻生物技术研发团队将“CO2/NOx吸收固定技术”、“微藻养殖技术”与“微藻生物质利用技术”集成,在烟气吸收、藻种筛选、新型光反应器开发、养殖技术开发与微藻产品加工利用方面进行了大量创新,建立了微藻用于碳氮减排和高蛋白生物质生产的新技术。这一技术利用微藻将石化行业排放的CO2和NOx进行固定和转化,将污染物转变为高价值的微藻生物质,为成功构建循环经济体系奠定了基础。目前已经在中国石化催化剂长岭分公司、中国石化湖北化肥分公司与中国石化石家庄炼化分公司(简称石家庄炼化)等地开展工业放大试验,在治理碳氮排放的同时生产微藻生物活饵料、微藻水质改良剂、鱼虾及畜禽饲料原料、高品质饲料添加剂等高附加值产品,取得了良好效果。图1为石家庄炼化微藻减排炼油厂CO2示范装置的照片,图2为其微藻养殖池的照片。

图1 石家庄炼化微藻减排炼油厂CO2示范装置照片

图2 微藻养殖池照片
6 总结与展望
微藻生物技术被认为是能够解决人类社会发展所面临诸多问题的有效手段之一。过去数十年来,微藻的商业化生产与应用也为实现这一目标带来了曙光,但展现在研究者和产业界面前的路还很长,发展进程中也需要继续解决技术、资金、政策监管与发展方式等诸多问题。展望未来,微藻生物技术的开发与应用仍需要坚持创新驱动发展,坚持绿色可持续的发展概念,兼顾碳氮减排社会价值与微藻产品经济价值体系的实现,为社会转向高质量发展阶段保驾护航。
