镇江长江豚类省级自然保护区浮游动物群落结构特征及影响因素
2021-10-09谈金豪蔺丹清张家路
谈金豪,蔺丹清,代 培,张家路,刘 凯,*
1 农业农村部长江下游渔业资源环境科学观测实验站, 无锡 214081 2 中国水产科学研究院淡水渔业研究中心, 无锡 214081
浮游动物群落作为水生态系统中常见的生态类群,在水生食物网的物质和能量循环过程中起到了至关重要的作用[1]。浮游动物的存在不仅影响到微生物和浮游植物的群落结构变化,而且作为鱼虾类的饵料生物,会通过“上行效应”影响渔业资源的变动[2]。同时,研究表明浮游动物群落结构受水温、pH及氮磷等水环境因素的制约[1,3],对水体环境变化敏感。因此,对浮游动物群落结构进行研究和分析,可以客观地反映出水域生态环境的优劣。
江苏镇江长江豚类省级自然保护区位于长江镇江江段,范围自上而下包括润扬大桥以下世业洲洲尾、定易洲及焦北滩洲滩及主航道水域(南侧以焦北滩夹江为界)、和畅洲北汊洲滩、边滩及水域,总面积57.3 km2,其中核心区面积约14.9km2,缓冲区面积约22.8km2。保护区地理位置独特,生态环境良好,渔业资源丰富,历来是长江江豚活动较为频繁的水域[4- 5]。随着长江江豚栖息地的日趋碎片化,镇江长江豚类省级自然保护区已经逐渐成为长江下游长江江豚群体间基因交流的重要生态走廊之一,对于维持长江下游长江江豚栖息地的完整性具有重要生态学意义。从保护区建成至今,已有代梨梨等[6]在2010年对保护区浮游动物群落结构特征及长江江豚生存状况进行了评估,然而随着南京以下12.5m水航道整治工程的实施,保护区面临着更严峻的考验,开展浮游动物群落演替的长期跟踪监测和阶段性全面调查,对目前江豚栖息地的保护性研究至关重要。
本文于2016—2018年按季度对镇江长江豚类保护区浮游动物群落现状进行了调查,探究其群落结构和水环境因子的关系,结合过去的研究成果分析近年来浮游动物群落结构的演替特征,以期为镇江长江豚类省级自然保护区生境的保护和管理提供科学的依据与对策。
1 材料与方法
1.1 采样时间和采样断面
镇江长江豚类保护区浮游动物群落结构调查于2016年至2018年开展,每年按季节分别于春季(4月)、夏季(6月)、秋季(10月)和冬季(1月)各采样一次,三年共计采样12个频次。根据镇江长江豚类保护区水域特点,选取6个采样断面,具体采样点如图1所示。每个断面设置3个样点,即断面的南岸、北岸及中间(南、北岸样点距岸约50m,各调查时期样点水深0.6—15.6m,多数小于10m,最大水深出现在2018年夏季的断面3南岸;江中间样点水深均大于20m,介于22.8—32.9m之间),其中断面1至断面3位于保护区临近的主航道水域(图1),断面4和断面5位于保护区核心区,断面6则位于保护区的实验区和缓冲区。

图1 镇江长江豚类保护区采样截面Fig.1 Sampling sites in Zhenjiang Yangtze dolphin nature reserve
1.2 实验方法
1.2.1浮游动物样品采集与分析
原生动物和轮虫类样本的定量采集,使用5L有机玻璃采水器,在各样点水体表层(0.5m)和底层(离底0.5m)分别采水,取均匀混合水样1L后,加入15mL鲁哥试剂和4%的甲醛溶液固定,带回实验室静置48h后,用虹吸管吸去上层清液,浓缩并定容至50mL,保存等待镜检;枝角类和桡足类的样本则使用5L玻璃采水器,采集20L水样,经25#浮游生物网过滤浓缩,放入100mL样本瓶,加入4%的甲醛溶液固定,带回实验室等待镜检。
参照《淡水浮游生物研究方法》、《原生动物学》、《中国淡水轮虫志》、《中国动物志(淡水枝角类)》和《中国动物志(淡水桡足类)》等[7- 11]鉴定浮游动物种类,原生动物和轮虫均取1mL浓缩水样,采用1mL计数框计数,放置于10×10显微镜下每个样本重复计数3—4次,取其平均值进行数据统计;甲壳动物(枝角类和桡足类)则将浓缩水样中的个体全部计数。
浮游动物密度和生物量换算参考《淡水浮游生物研究方法》[7],浮游动物生物密度计算公式为
N=(Vs×n)/(V×Va)
式中N为1 L水中浮游动物的个体数(个/L),Vs为浓缩样品的体积(mL),n为计数所得的个体数,V为采样体积(L),Va为计算体积(mL)。
式中,B为1L水样中浮游动物总生物量(mg/L),Ni为1L水样中第i种浮游动物个体数,Wi为第i中浮游动物重量系数。
1.2.2水质理化因子测定
水温、酸碱度和溶解氧等指标使用携式水质检测仪(HANNA HI 98703、HACH-HQ30D&2100Q)现场测定,透明度使用塞氏盘测定;总氮、总磷和叶绿素a等水质指标,按水样采集规范采集,在各样点水体表层(0.5m)和底层(离底0.5m)分别采水500mL,水样均匀混合后,存放于玻璃瓶中,放置在4℃冷藏箱中带回实验室,在24小时内参照《水和废水监测分析方法》[12]进行测定。监测的水质数据按照《地表水环境质量标准》(GB3838—2002)进行分析。
1.3 数据分析和处理
运用Shannon-Wiener指数(H′)、Margalef指数(D)[13]评价保护区浮游动物群落结构的多样性,并评价保护区的水质状况[14],优势度指数(Y)确定浮游动物群落中的优势种。
(1) Shannon-Wiener(H′)指数
H′ =-∑ (Ni/N) log2(Ni/N)
(2) Margalef(D)指数
D=(S-1)/lnN
(3) 优势度指数(Y>0. 02 时,即为优势种)
Y=(Ni/N)×Fi
式中,N为样品中的总个体数量;Ni为第i种的个体总数量;Fi为第i物种出现的频率;S为样品中种类总数。多样性指数的水质评价标准为:0—1为多污型、1—2为α-中污、2—3为β-中污、3—4为寡污型、>4为清洁。
相关性分析:选取浮游动物的物种数、密度和生物量,以及水环境因子指标。对数据进行log(X+1)对数转化使之呈正态分布,利用Pearson相关性分析法分析浮游动物群落结构与环境因子的相关性。
冗余分析(RDA)分析:基于canoco4.5软件对浮游动物与环境因子进行RDA分析。在进行RDA分析前,进行去趋势对应性分析(DCA),若DCA排序结果小于3.0,则选取RDA分析。
渔产力评估:通过浮游动物生物量来计算镇江保护区的渔产潜力(F),从而评估保护区江豚容纳量。
F=(b×P/B×a)/K
式中,b为浮游动物平均生物量;P/B指浮游动物现存量与生物量之间的比值;a为鱼类对浮游动物的利用率;K为鱼类摄食的饵料系数。参考相关文献[6],确定P/B系数为40,a为30%,K为10。
2 结果
2.1 环境因子分析
对保护区水域2016—2018年的调查结果显示(表1),水温7.6—25.9℃,均值为18.2℃,均为夏季最高,冬季最低;pH变幅为7.42—8.49,均值为8.04,保护区水体总体偏碱性。总氮(TN)、总磷(TP)和叶绿素a(Chla)年度波动较大,其中TN变幅为0.52—2.84mg/L,均值为2.00mg/L;TP变幅为0.01—0.38mg/L,均值为0.14mg/L;Chl-a变幅为0.75—15.93 mg/m3,均值为3.88 mg/m3。各水环境指标季节性差异较为显著(P<0.05)。
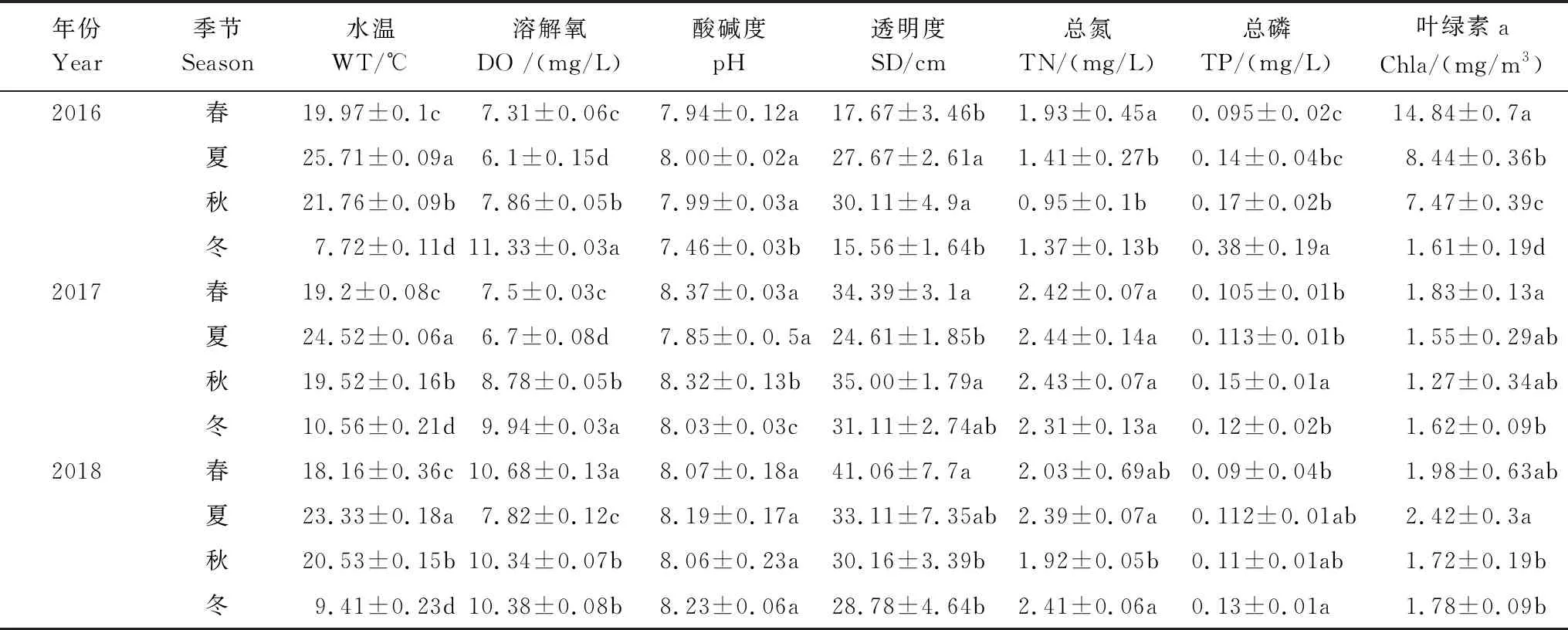
表1 镇江保护区不同季节环境参数
根据《地表水环境质量标准》(GB3838—2002)评价镇江保护区水质,结果表明保护区水域DO、pH和TP等指标符合Ⅲ类的水质标准,TN指标则大部分超出。综合而言,镇江保护区水质达到Ⅲ类水质的功能标准,TN成为主要制约因素。
2.2 浮游动物种类组成
2016—2018年保护区江段共鉴定出浮游动物90种,其中原生动物物种数最多,共40种,占浮游动物物种总数的比例为43.01%;其次为桡足类有22种,占比23.66%;枝角类15种,占16.13%;轮虫类最少,仅13种,占13.98%。其中2016年鉴定出浮游动物42种,夏季最多,冬季最少;2017年60种,秋季最多,冬季最少;而2018年共41种,春季最多,冬季最少。各年度浮游动物种类组成较稳定,无明显差异,总体表现为原生动物最多,其次为桡足类,轮虫类和枝角类占比较少。各年度种类数季节性差异较为显著(图2),而空间分布差异不明显(图3)。
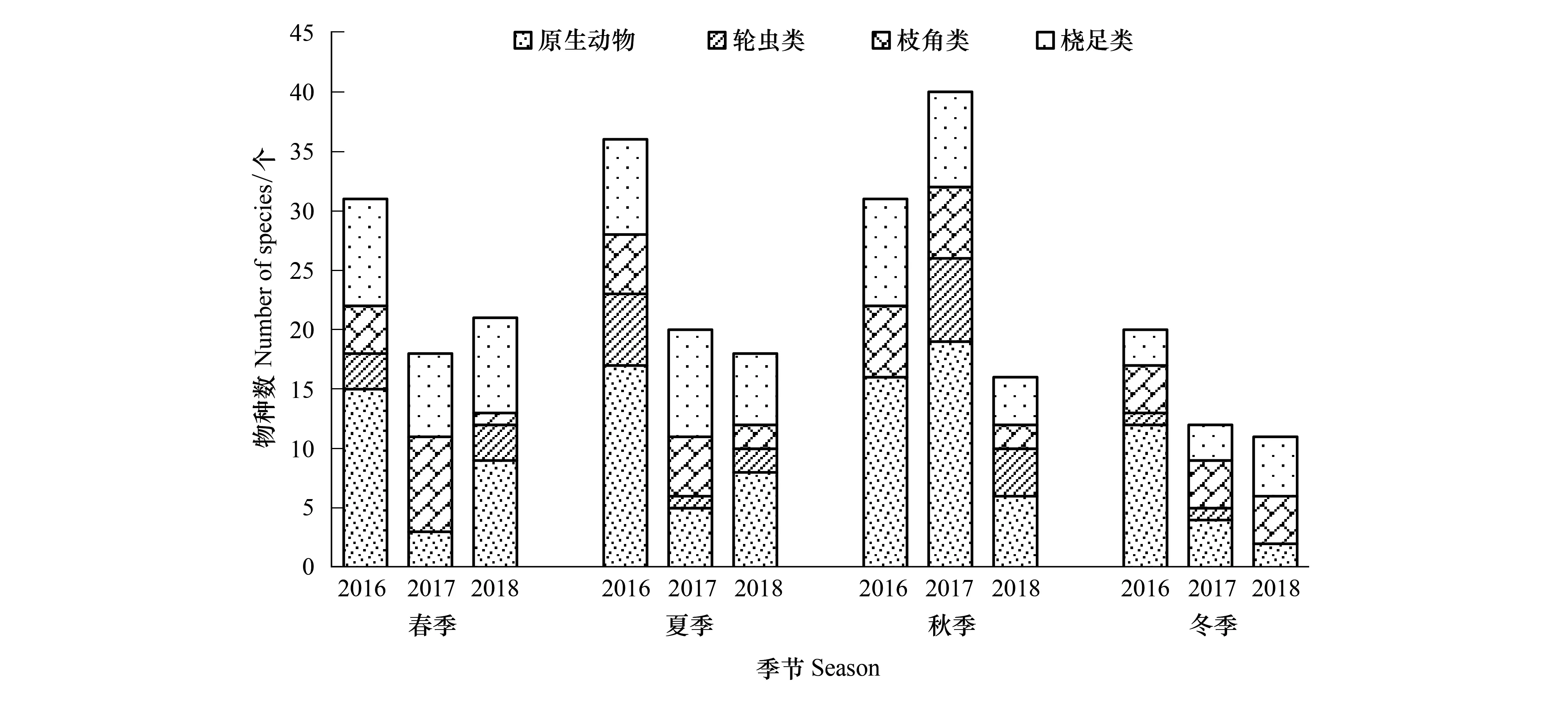
图2 2016—2018年保护区浮游动物物种数季节变化Fig.2 Seasonal variation in species composition of zooplankton in Zhenjiang nature reserve from 2016 to 2018
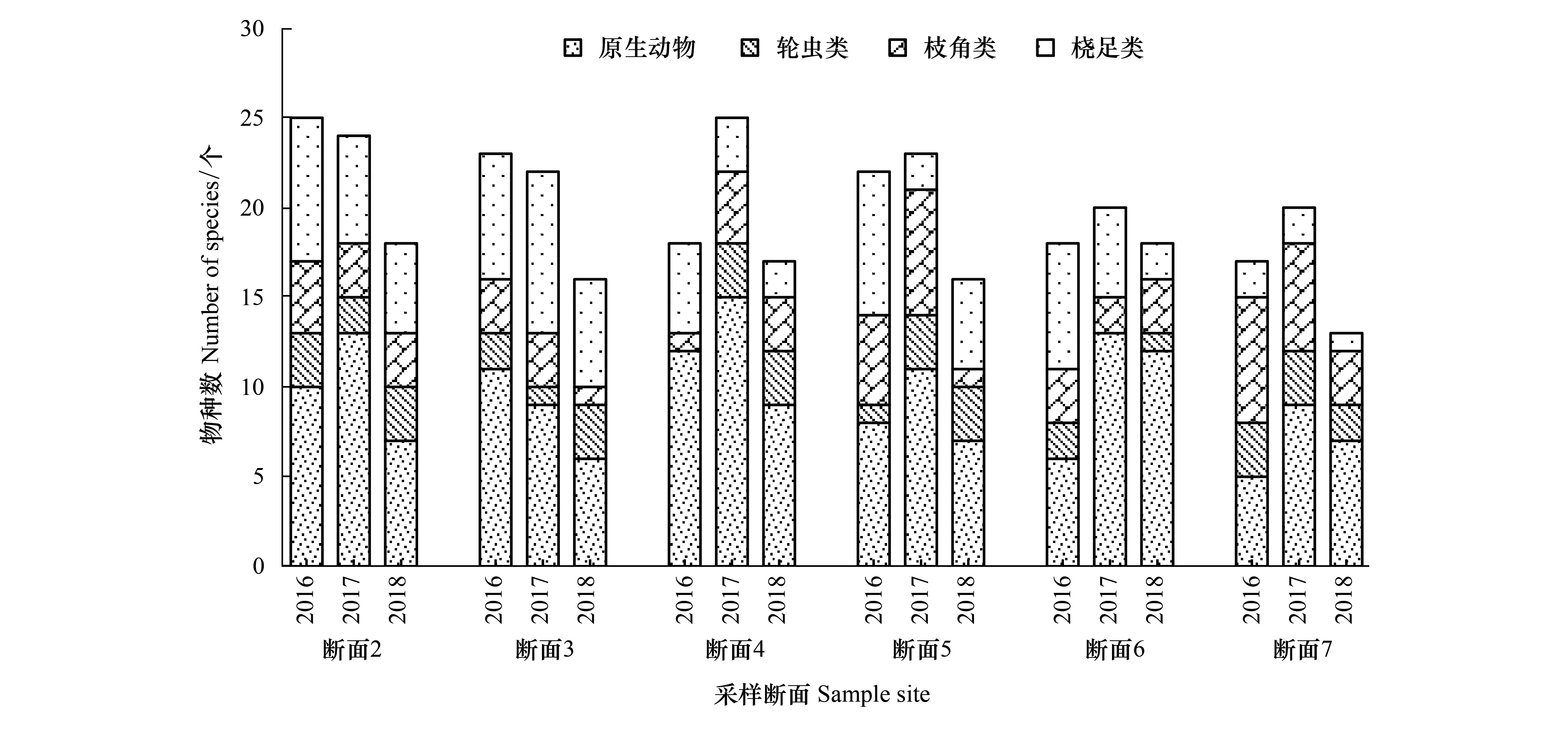
图3 2016—2018年保护区浮游动物物种数空间变化Fig.3 Spatial variation in species composition of zooplankton in Zhenjiang nature reserve from 2016 to 2018
2.3 优势种
2016至2018年镇江保护区浮游动物优势种类共计8种,包括7种原生动物和1种轮虫类。表2可见,从种类组成上看,原生动物中淡水麻铃虫 (Leprotintinnusfluviatile)、恩茨筒壳虫 (Tintinnidiumentzii)、王氏似铃壳虫(Tintinnopsiswangi)和江苏似铃壳虫(Tintinnopsiskiangsuensis) 连续三年都有出现,呈季节性变化,春季和冬季的主要优势种为王氏似铃壳虫和恩茨筒壳虫,而在夏季和秋季则是淡水麻铃虫和江苏似铃壳虫占优。根据原生动物和轮虫的水体营养类型指数种类表[15- 16],原生动物的优势种多为喜清洁水体类群,而2018年春季出现的轮虫优势种-针簇多肢轮虫,属于富营养水体指示种类。
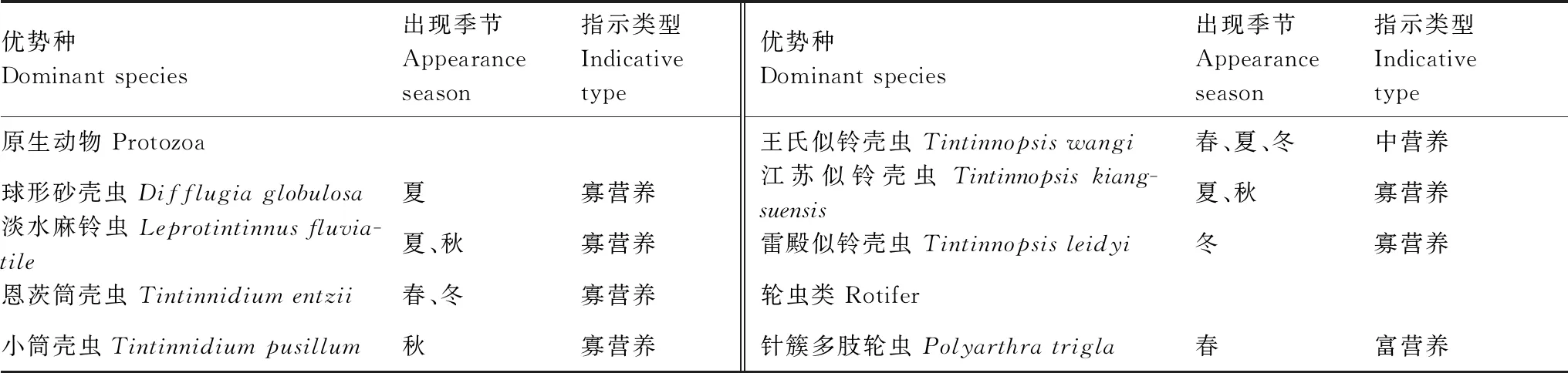
表2 保护区浮游动物优势种
2.4 浮游动物密度和生物量
镇江保护区2016—2018年浮游动物的密度和生物量的季节变化和水平分布见图4、图5。2016年浮游动物年均生物密度为557.66个/L,生物密度季节变化为秋季(861.91个/L)>夏季(778.92个/L)>春季(295.29个/L)>冬季(150.07个/L),存在极显著的季节性变化(ANOVA,F=48.138,P<0.01); 2017年年均生物密度均值为293.19个/L,季节变化与2016年相一致,秋季显著高于其它季节,季度间存在极显著差异(ANOVA,F=93.232,P<0.01);2018年年均密度最小,为115.6 个/L,季节性差异显著(ANOVA,F=3.296,P<0.05),生物密度的高峰在春季(150.07个/L),低谷出现在冬季(64.44个/L)。各年度生物密度样点间空间分布均无显著差异(P>0.05)。
2016年年均生物量为0.06mg/L,夏季最高(0.155mg/L),冬季最低(0.01mg/L),季度差异极显著(ANOVA,F=15.711,P<0.01);2017年生物量均值为0.06mg/L,最大值为0.18mg/L,出现在秋季,最小值出现在冬季为0.003mg/L,生物量存在极显著的季节性差异(ANOVA,F=30.216,P<0.01);2018年生物量的季节变化也较为显著(ANOVA,F=4.173,P<0.05),从高到低依次为夏季(0.067mg/L)>秋季(0.038mg/L)>春季(0.025mg/L)>冬季(0.006mg/L)。各年度生物量空间分布同样无显著差异(P>0.05)。

图4 2016—2018年保护区浮游动物密度和生物量季节变化Fig.4 Seasonal variation in density and abundance of zooplankton in Zhenjiang nature reserve from 2016 to 2018
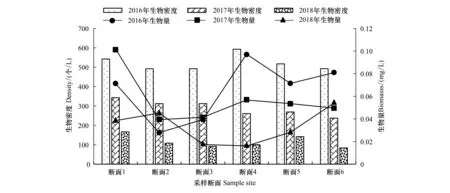
图5 2016—2018年保护区浮游动物密度和生物量空间变化Fig.5 Spatial variation of zooplankton density and abundance in Zhenjiang nature reserve from 2016 to 2018
总体来说,保护区浮游动物的密度组成中原生动物占绝对优势,占比超过91%,其次为轮虫,占8.5%左右,而枝角类和桡足类占比1%都不到。从2016年到2018年浮游动物的密度和生物量均呈现逐年下降趋势,且都存在显著的季节性差异,高峰一般都出现在夏秋季节,而冬季的密度和生物量均处于最低。从空间特征来看,无论是保护区核心区、缓冲区,还是主航道之间,密度和生物量的差异都不显著。
2.5 多样性指数
对保护区2016—2018年各年度四季的浮游动物生物多样性指数进行了比较(图6, 7)。结果表明,2016年Shannon-Wiener 多样性指数H′年均0.88,秋季最高(1.02),冬季最低(0.78),存在显著的季节性差异(ANOVA,F=5.344,P<0.01);2017年年均值0.68,季节性差异极显著(ANOVA,F=8.761,P<0.01),最大值出现在秋季,最小值在冬季;2018年Shannon-Wiener多样性指数H′全年变化范围为0.33—0.45,年均0.4,季节变化为秋季最高,春季最低。
2016年Margalef丰富度指数D变幅为0.66—2.33,年均1.13,季节变化特征为春季最高,冬季最低(ANOVA,F=3.251,P<0.01);2017年年均值为1.29,季节性变化与上年一致(ANOVA,F=4.313,P<0.01);2018年Margalef丰富度指数D年均2.37,最大值则出现在冬季(3.8),最低值在秋季(0.62)。
保护区浮游动物Shannon-Wiener多样性指数H′和Margalef丰富度指数D均存在显著的季节性差异,而各样点间的差异则不大。根据多样性指数来评价保护区的水质,春、冬季两属于中污-多污型,夏、秋两季多属于中污型。总体而言,保护区水质属于中污型。
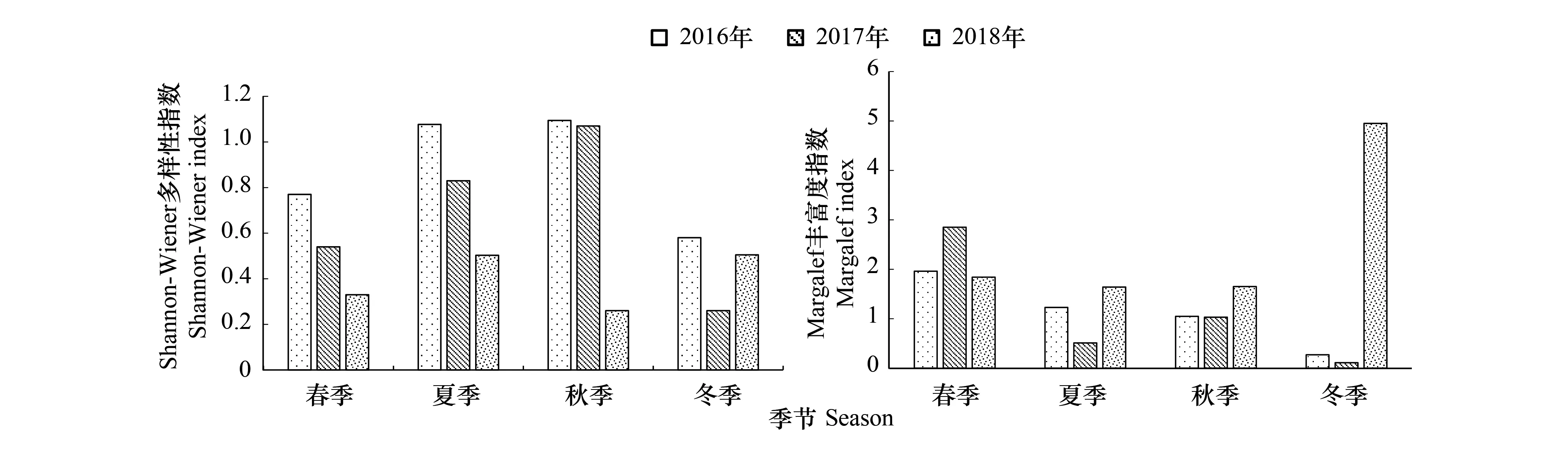
图6 2016—2018年保护区浮游动物多样性指数季节变化Fig.6 Seasonal variation of zooplankton diversity index in Zhenjiang nature reserve from 2016 to 2018
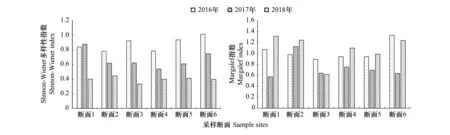
图7 2016—2018年保护区浮游动物多样性指数空间变化Fig.7 Spatial variation of zooplankton diversity index in Zhenjiang nature reserve from 2016 to 2018
2.6 浮游动物群落结构与环境因子的相关性分析
2.6.1浮游动物与环境因子的相关性分析
当环境变量间的相关程度较高,易导致系数估计的误差,会影响到Pearson相关性分析的结果。因此,在对保护区浮游动物群落与环境因子进行Pearson相关性分析之前,对本文选取的水温(T)、溶氧(DO)、总氮(TN)、总磷(TP)和叶绿素a(Chl-a)等7个环境数据进行共线性检验,VIF>10表现共线性显著,若VIF介于0—10之间,则表示共线性不显著。检验结果如下表所示(表3),结果显示选取的环境数据共线性并不显著,故本文选取的7个环境数据可以作为研究浮游动物密度变化的影响因子。
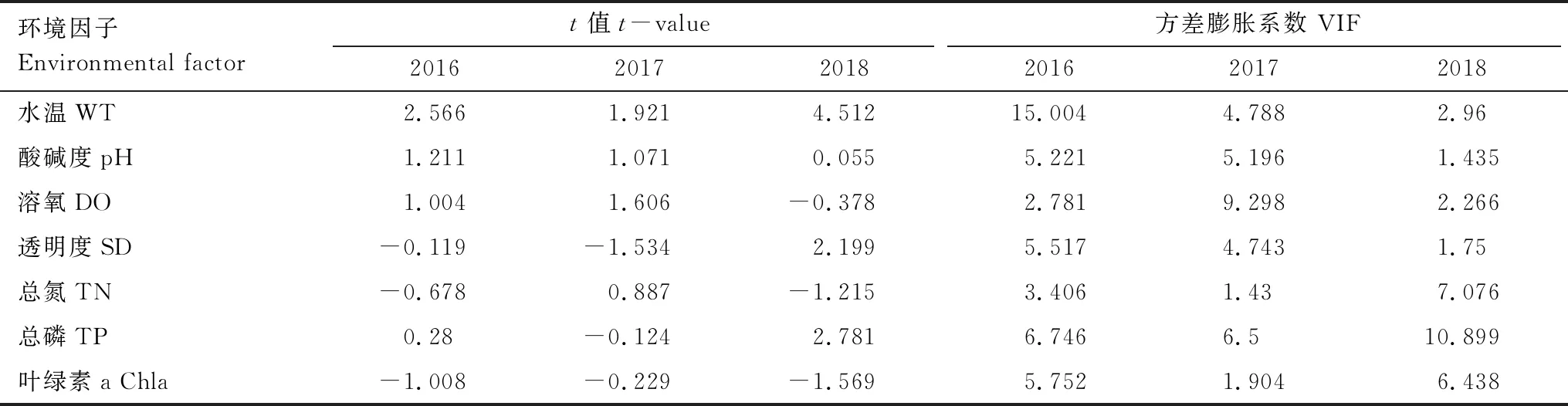
表3 环境因子共线性诊断
对镇江保护区环境因子与浮游动物的密度进行相关性分析,结果表明(表4),浮游动物的物种数、密度和生物量,与水温、透明度、TN、TP和叶绿素a呈显著正相关,与溶氧、pH呈显著负相关。由此可见,浮游动物的密度和生物量会随着水温的升高,或是营养盐浓度的增加而增加,同时,叶绿素a和透明度的提高也会对浮游动物的生长起到一定促进作用。而水体碱性的提高,以及过高的溶氧,可能会导致浮游动物的数量有所下降。
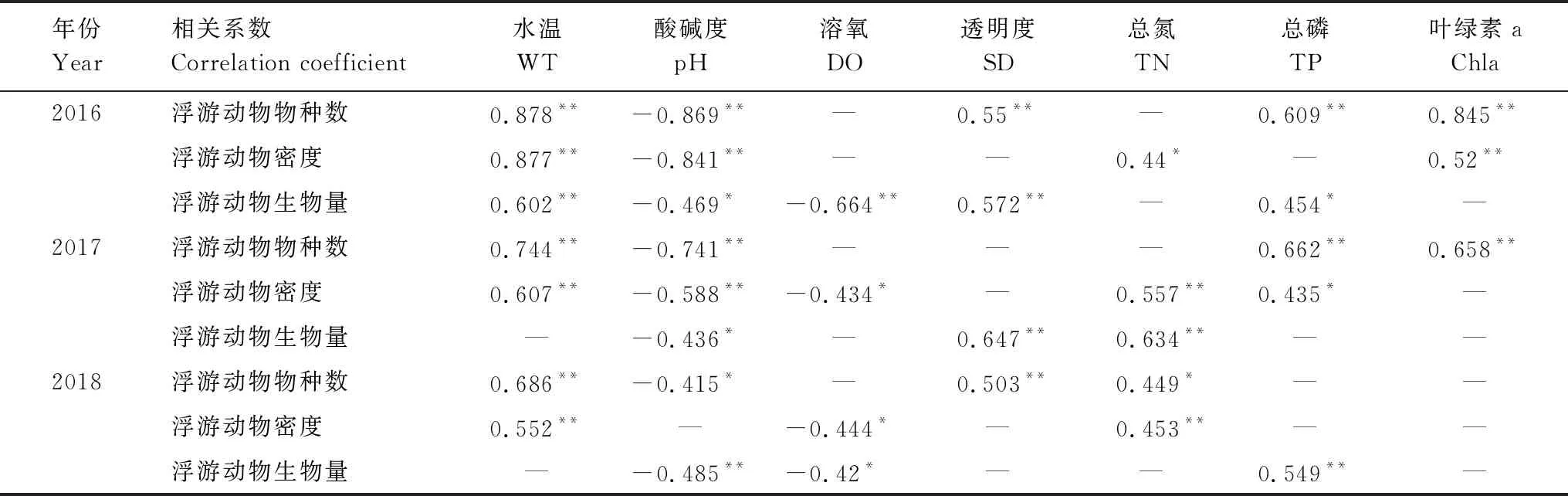
表4 保护区浮游动物与环境因子相关性分析
2.6.2浮游动物与环境因子的RDA分析
为了进一步探究镇江保护区环境因子对浮游动物的影响,本文综合各年度调查中常见的物种,根据浮游动物密度在一个样点中至少≥1%,且出现频率≥25%,筛选出14种浮游动物进行DCA分析(表5),结果显示,排序轴长度小于3,故选择RDA进行典范对应性分析。RDA分析结果显示,前两轴的特征值分别为24.9%和17.09%,共解释了浮游动物群落整体的74.78%,物种和浮游动物的相关性都达到0.9,表明RDA排序图可以很好地反映环境因子对浮游动物的影响。
表5 典范对应分析浮游动物代码
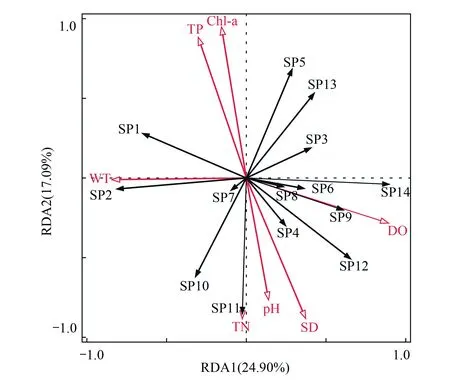
图8 镇江保护区浮游动物与环境因子RDA分析 Fig.8 RDA analysis of zooplankton in Zhenjiang nature reserveWT: 温度; DO: 溶解氧; Chla: 叶绿素a; SD: 透明度; TP: 总磷; TN: 总氮; SP1: 球形砂壳虫 Difflugia globulosa; SP2: 淡水麻铃虫 Leprotintinnus fluviatile; SP3: 恩茨筒壳虫 Tintinnidium entzii; SP4: 雷殿似铃壳虫 Tintinnopsis leidyi; SP5: 江苏似铃壳虫 Tintinnopsis kiangsuensis; SP6: 王氏似铃壳虫 Tintinnopsis wangi; SP7: 螺形龟甲轮虫 Keratella cochlearis; SP8: 小多肢轮虫 Polyarthra minor; SP9: 针簇多肢轮虫Polyarthra trigla; SP10: 长额象鼻溞 Bosmina longirostris; SP11: 小栉溞 Daphnia cristata; SP12: 无节幼体 Copepod nauplii; SP13: 如愿真剑水蚤 Eucyclops speratus; SP14: 球状许水蚤 Schmackeria forbest
物种与环境因子的RDA分析排序图显示(图8),淡水麻铃虫、球形砂壳虫受水温的影响比较大,与之呈现密切的正相关关系,其密度会在夏、秋季较高。小多肢轮虫和针簇多肢轮虫与溶氧的正相关性较大,反之螺形龟甲轮虫偏向于溶氧较低的水体。小栉溞喜好在氮营养程度高,且偏碱性的水体中生存,与TN和pH密切相关。雷殿似铃壳虫则喜欢生活在透明度高的水体中。因此,水温、pH、透明度、TN和溶氧是影响镇江保护区浮游动物群落的重要因素。RDA分析排序图不仅能解释环境因子对浮游动物群落结构的影响,而且能反映出不同物种的生态适应性,比如恩茨筒壳虫和螺形龟甲轮虫在排序图中处于对立位置,表现出显著差异的环境需求;而小多肢轮虫和针簇多肢轮虫在排序图中的位置较为接近,说明两者的生态习性较为相似。
2.7 渔产力及江豚容纳量评估
根据2016至2018年浮游动物的年均生物量,通过渔产力公式计算分析可以得出(表6),保护区各年的渔产潜力分别为1163.96kg/km2、1026.03kg/km2和610.22kg/km2,按每头长江江豚每年摄食1500kg计算,那么理论上保护区对长江江豚的容纳量范围在23—44头。然而,综合考虑到江豚适口鱼类资源的分布等环境因素的限制,实际上保护区内可以利用的渔产力将远远不足,江豚的容纳量也远小于最低值。同时,对三年的渔产力及江豚容纳量对比发现,两者呈现出逐年下降的趋势。
3 讨论
3.1 保护区浮游动物群落结构特征
通过2016—2018年连续三年对镇江保护区浮游动物群落的调查,结果显示3个年份浮游动物群落结构相对稳定,均以原生动物占优,而轮虫类、以及个体较大的枝角类和桡足类占比相对较少,发现的8种优势种也主要为食藻、食菌和碎屑功能类群原生动物。浮游动物密度和生物量构成中,原生动物是密度的主体,而生物量中则是枝角类和桡足类共同占有优势,密度和生物量存在显著的季节性变化,总体表现为夏、秋季高于春、冬季。相比于长江干流湖北段[17]、江苏段[18,19]而言,保护区江段物种数和生物量略少,其密度量要高,这可能与江心洲有关[20],和畅洲作为长江下游典型的江心洲滩,洲滩附近水生植被丰富,水质稳定,为一些特定种类提供了良好的栖息环境。本次调查结果与代梨梨等[6]对镇江保护区浮游动物群落结构特征的历史研究数据相比,原生动物、枝角类和桡足类占总数的比例结构类似,但轮虫类占比呈显著下降趋势。同时,与国内其他河流相比较,本研究与嫩江[21]、富春江[22]、松花江[23]以耐污、喜富营养水体的轮虫类群为主的群落组成也有所不同。造成这种差异的主要原因可能是水文条件和水体营养状况的联合作用。一方面,和畅洲沿岸为航道整治修建的丁坝,导致镇江段水流速加大,泥沙量增加,透明度降低,不利于环境敏感性轮虫的发展[24],比较而言,原生动物对此具有更强的适应性[20]。另一方面,保护区周边自然生态良好,水体营养水平不高,在一定程度上影响到轮虫的生长和繁殖[25],多数富营养化水体指示种无法形成优势类群。

表6 保护区渔产潜力及江豚容纳量估算
3.2 浮游动物群落结构变化与环境的关系
影响河流湖泊浮游动物群落结构的环境因素一直是淡水浮游动物生态学研究的重点,在阐释河流浮游动物群落的变化时,通常会探讨水温、营养盐、酸碱度(pH)等水体理化因素的影响。根据本研究浮游动物与环境因子的Pearson相关性和RDA分析结果显示,对镇江保护区浮游动物群落结构的影响较为显著的环境因子是水温、TN、pH和溶解氧。温度通常被认为是影响浮游动物种类组成、数量和分布的最主要因素[2,14]之一,在本次调查中,浮游动物密度和生物量的变化与水温的变化趋势基本一致,随着水温的升高,浮游动物繁殖速度加快,种群数量会迅速增长,因此,密度和生物量在夏、秋季节达到顶峰。有研究表明[26- 27]水体中的氮等营养盐浓度变化,能够影响浮游动物群落的演替。聂雪等[28]对鄱阳湖子湖浮游动物群落结构的研究结果显示,营养盐通过调控浮游植物来间接影响浮游动物群落结构,氮水平升高使得水体的营养化水平提高,造成浮游植物大量繁殖,浮游植物及其分解后的碎屑可以为浮游动物提供更为丰富的食物来源,从而促进浮游动物的生长繁殖。在本研究中TN与浮游动物密度的著正相关关系,体现出TN对浮游动物密度增长的向作用。溶氧和pH也会影响浮游动物群落的变化,郭坤等在对湖北长湖[29]的研究中发现,春、夏季节大部分优势种与酸碱度、溶氧呈负相关,较高的溶氧和pH抑制了一些轮虫和枝角类的生长。对比本研究中物种数量和密度与pH、溶氧呈现的显著负相关性,可以推测保护区江段的大多数物种对偏碱性、高溶氧的水体具有良好的适应性。
相比与湖泊、水库等静态水体,河流是具有单一流向的线型生境,流动状态、水文状况、岸线类型等诸多条件,都会对河流浮游动物群落的形成产生重要影响[30]。Pace等[31]对哈德逊河浮游动物群落的调查中发现河流的流动状态与浮游动物丰度存在密切的相关性,不同河流、湖泊之间水体滞留时间的差异,会导致浮游动物丰度的差异,即使是在同一条河流的不同河段,其浮游动物群落的丰度也不完全相同。与沿江一些湖泊相比[32- 33],本研究中浮游动物的低密度和生物量印证了这一观点。Matsumura等[34]在研究和对比了巴西兴谷河旱季和雨季前后的浮游动物的组成和丰度后认为,降水量、以及浊度和径流量也是浮游动物群落的显著影响因子。考虑到长江的汛期,保护区浮游动物的现存量在丰水期远高于枯水期,也可能与降水和径流量变化有关。近年来浮游动物与气候变化的关系成为热点[35],但相关研究在国内涉及较少,在今后的研究中应予以探究。

3.3 渔产潜力及江豚容纳量分析
