生态系统多稳态研究进展
2021-10-09赵东升张雪梅
赵东升,张雪梅
1 中国科学院地理科学与资源研究所,中国科学院陆地表层格局与模拟重点实验室, 北京 100101 2 中国科学院大学, 北京 100149
由于人类对化石燃料的大规模应用,导致空气中CO2的浓度迅速升高,使得地球温度持续上升。IPCC 第五次评估报告指出,在1880年至2012年期间,地球平均温度上升了0.85℃[1],气温升高也导致了极端气候事件频率和强度显著增加。随着人类对资源的过度开发利用,导致环境破环,湖泊和河流中富营养化、有毒化学物质浓度升高,平原地区地下水位下降,海洋捕捞率上升,森林景观破碎化、草原过牧和生物多样性减少等。过去的研究大都认为气候变化和人类活动对生态系统的影响是渐进的、可预测的和可逆的[2]。但研究发现,生态系统也经常会出现系统性的突变,即在外力的驱动下发生稳态转换,如珊瑚礁的突然崩溃[3],绿洲突然转变为沙漠[4],湖泊中突然爆发的水华[5],以及害虫(蝗虫)的突然爆发等[6]。渐进影响与突然稳态转换的相互作用,增加了生态系统响应外界胁迫预测的难度。人类以高效率最佳状态的驱动力去响应其变化,往往会使得整个生态系统更容易遭受到意外事件的伤害[7]。
目前在微观水平上对生命的了解正在不断加深,但是对生态系统响应外界胁迫的稳定性和弹性机理知之甚少。对生态系统突变的研究,国内外学者进行了一些理论探索,但实践研究还相对较少。生态系统的灾难性突变对生态系统结构和功能的稳定性、生态系统服务的持续性和人类生存环境维持起着重要的作用。生态系统一旦发生突变将很难恢复,因此如何利用多稳态理论对生态系统突变进行早期预警,并对退化生态系统恢复提供科学的依据,是国际上生态学研究的热点[8]。其中弹性的研究受到各国学者们的较多的关注[7],但是对于复杂生态系统突变预警信号的研究仍显滞后。考虑到目前全球生物多样性的不断丧失和生态系统结构和功能退化的情况下,多稳态理论可以为退化的生态系统是否可能在人类干预下进行恢复提供一个关键的诊断工具,对生态系统的管理有着重要的意义[9]。
本文通过对多稳态概念的整理得出了一个更加综合的多稳态定义,并以放牧模型为例,解释了多稳态理论的相关概念,对生态系统稳态转换的各个预警信号的数学原理、优缺点和应用条件进行了分析,并对不同尺度下生态系统多稳态的研究方法进行了总结。最后展望了多稳态理论未来研究的问题和重点,为多稳态理论和实践的深入研究提供科学支撑。
1 生态系统多稳态理论
1.1 多稳态的概念
多稳态(alternative stable states)的概念是1969年由 Lewontin[10]研究“在一个给定的生境中是否会有二个或二个以上的稳定生物群落结构存在?”时提出的。Scheffer[2]和Dakos[3]对多稳态的理解是系统在相同条件下可能收敛到不同的状态。Clements 和Ozgul[11]则认为多稳态是生态系统的另一种形态,其特征是通过负反馈调节维持其功能和组成的变化。总而言之,多稳态是指在相同的外力驱动或干扰的情况下,其生态系统内生物群落的结构、物质和能量都会发生变化,并且可能表现为由负反馈调节维持的两种及以上不同的稳定状态。稳态在生态学领域是指生态系统现有的结构和功能在一定的时间和空间尺度上保持不变。
多稳态理论中的稳态是由负反馈调节所维持的,需要引入一定量的负熵流,才能保持稳定的状态,而系统突变是由正反馈所驱动的,需要外界条件朝一定的方向持续的变化才会发生稳态转换[12]。生态系统的演替也可以理解为稳态转换的过程,生态学家们需要了解生态系统可能存在的不同的状态和状态变化是否容易逆转,以及可能在哪发生转换,以预测突然变化可能发生的时间和原因。所以,多稳态理论在生态学上有着重要意义,有助于对生态系统状态变化的预测,并影响对生态系统的管理与决策[8]。
1.2 多稳态理论的阐释
1.2.1放牧模型
为了深入理解生态系统多稳态系统的动态,需要用数学模型建立一个理论框架来理解其动力学机制。Noy-Meir 在1975开发的牧场放牧模型可以很好的解释其动力学机制,在这个模型中植被的生长取决于资源的可获得性,而食草动物的密度是由人类管理者控制的。植被(V)随时间(t)的动态变化可写为[13]:
(1)
式中,G(V)为植被生长的函数;c(V)为食草动物(H)消耗植被的函数。当植被(V)随时间(t)的动态变化率为0时出现平衡,即dV/dt=0,此时G(V)=c(V)H。在图形上把植被生长函数和消耗植被函数绘制在一起时,交点即为平衡点(图1)[14]。

图1 放牧模型(图片引用于Beisner[14])Fig.1 Model of the Grazing Animals on Pasture Land
图1左用Logistic方程作为植被生物量函数,其中K代表环境承载力;图1中是食草动物对植被生物量的消耗作为植被生物量的函数,三条线分别代表高强度、中强度和低强度放牧情况下植被的总消耗量。图1右为植物生长和植被消耗的结合,其中黑色的实心点表示稳定的平衡点。在中度放牧的情况下,以P点为例,当V向右移动时(C线>G线),也就是植被的消耗量大于植被的生产量;当V向左移动时(G线>C线),也就是植被的生产量大于消耗量,因此系统总是会返回到原来的平衡点。而空心点N则表示不稳定的平衡点或临界点。V的轻微增加会使得植被的增长大于消耗(G线>C线),并随着植被的生长继续向右移动,直到它们达到平衡点P;V的轻微下降会导致植被的消耗量大于生产量(C线>G线),因此V会进一步向左移动,到达较低的平衡点Q[13-14]。
1.2.2相位空间图

图2 多稳态相位空间(图片改编于Scheffer和Carpenter[15])Fig.2 Phase Space of Alternative Stable States
为了更好的理解多稳态理论,可以使用相位空间在V与H的关系图中绘制所有的可能平衡点(图2)[15]。图2中的小球表示生态系统的状态,更倾向于落在稳定平衡的山谷中。山顶表示不稳定平衡的蓝色虚线,并标志着不同吸引盆地之间的边界。外部条件的变化影响着稳定的格局。当外部条件发生不利的变化时,吸引盆地可能会缩小,当它最终消失时(F2),就会突变到另一个吸引盆地。
图2中的状态A是放牧模型中的低强度模式,AB段的绿线为低强度放牧下的稳态,为草原生态系统。BD段的蓝色实线为中等放牧模式下的稳态,此时草原生态系统草的数量和质量大大减少。当放牧强度达到高强度时(图2中的状态D),轻微的扰动,便会使得草原生态系统转变为荒漠。DE段的红线为稳定的荒漠生态系统。其中图2的F1和F2点为临界点、阈值或分歧点(Tipping point / Threshold /Bifurcation Point),是指在系统非常不稳定的时候,即使是很小的扰动也会导致状态发生巨大的变化。从数学上讲,当占主导地位的特征值达到零时,系统从一个稳定状态的吸引盆地通过一个不稳定的区域快速转移到第二个吸引盆地[11]。指示临界点到来的一些特征或统计信号被称为预警信号(Early Warning Signal)。从草原生态系统到荒漠生态系统的转变称为稳态转换(Regime Shift/Critical Transition),是指生态系统由一种稳定状态向另一种稳定状态的转变,由系统的外部强迫引起[11]。
AB段的绿线为低强度放牧下的稳态;BD段的蓝色实线为中等放牧模式下的稳态;DE红线为高强度放牧下的稳态;F1和F2点为临界点在多稳态的生态系统当中,初始状态的扰动将决定系统可能达到的稳定状态。这也称为“路径依赖”,足够严重的扰动可以诱导向另一种稳定状态的转变。如果当前状态周围的吸引盆地较小,则系统更容易转化到另外一种状态[15]。其中吸引盆地的大小也称为弹性(Resilience),通常是指系统在保持功能的同时可以吸收的干扰量。当系统受到压力时,系统的弹性会发生变化,因为系统的外力会改变吸引盆地的形状,从而改变系统的弹性[10]。外部条件的变化可能会降低弹性,却不会对平衡状态产生较大影响。虽然此时的生态系统看起来没有改变,但是已经变得很脆弱,很小的扰动都会导致其向另一种状态转变[15]。
1.2.3生态系统恢复和演替
从荒漠生态系统恢复到最初的草原生态系统,仅仅恢复崩塌前的条件是远远不够的。如图2所示需要恢复的程度要远远超出分歧点F2,需要多恢复DB距离,直到分歧点F1,在那里系统才会自动突变到之前的状态。而多恢复的距离DB称之为滞后(Hysteresis),是指在灾难性突变之后缺乏可逆性,系统保持在一个状态,除非它达到另外一个分歧点才能恢复[11]。但是系统很难恢复到之前的状态,而是变成了另外一种自组织的状态,有可能重复上一循环,也有可能进入新的不同的循环(生长、积累、重组和更新),称为扰沌(Panarchy)[16]。如图3所示当系统由状态b恢复到F1点的时候,他可能突变到a,从而恢复到之前的状态,也有可能突变到e 从而达到另外一种状态,之后又开始一轮新循环。

图3 多稳态与生态系统演替、恢复和扰沌的关系Fig.3 The Relationship Between Alternative Stable States and Ecosystem Succession, Restoration and Panarchya, b, c, d, e, f, g, h, i, j, k, l为不同的生态系统状态,F1—F10 为不同生态系统稳态转换的临界点
用多稳态理论解释生态系统演替的过程:如图3所示,状态i为稳定的生态系统状态,随着外界环境条件的变化和内部的竞争到F8时,会突变到状态h系统; 若环境条件继续变换,则状态h会通过稳定的状态a系统,变化到F2时,之后突变到状态b系统;若环境条件依旧继续变化,状态b 将保持稳定通过状态c直到F3时,然后突变到d达到另外一种稳定的状态。从i—h(a)—b(c)—d的过程,是生态系统稳态转换的过程也是生态系统演替的过程。由内蒙古草原向西北荒漠的演替为例:状态i为稳定的半湿润草甸草原,随着降水量的减少,草甸草原的质量和数量都会大量减少,系统的弹性会降低,直到F8时,系统会由草甸草原突然转变到半干旱典型草原h(a);随着降水量继续减少,典型草原的弹性会下降,典型草原h会通过a渐渐到达F2,之后会突变到干旱荒漠草原b(c);若降水依旧减少,则荒漠草原将会由b通过c,直到F3时,荒漠草原会突变到干旱荒漠状态d。从半湿润草甸草原—半干旱典型草原—干旱荒漠草原—干旱荒漠的过程,是生物群落的稳态转换过程,也是群落演替过程。
2 稳态转换的预警信号
2.1 临界减速指标
临界减速是指在临界点附近,系统的稳定性下降,导致系统恢复到以前状态的速度变慢的现象。因为当靠近分歧点时,雅可比矩阵特征值的最大实部趋于零:动力系统从小扰动中恢复的速度越来越慢[17]。其中临界减速主要的指标有恢复率变慢、自相关、方差和条件异方差的增加[18]。Drake和Griffen为了验证临界减速(CSD)发出的预警信号,采取了大型蚤的复制种群进行了实验,表明由环境条件控制下降引起的稳态转换的种群,在环境恶化开始之后和临界转换之前,表现出CSD的统计特征[19]。
2.1.1恢复率变慢
在发生临界转换的分歧点处,占主导地位的实特征值变为零。随着系统越来越接近分歧点,之前稳定的吸引盆地的斜坡变得不那么陡峭,因此,当系统在向这样一个临界阈值移动时,从轻微扰动中的恢复将逐渐变得越来越慢,其速率逐渐变为零。当系统处于稳定的状态时,即系统的弹性很强,则从扰动中恢复所需要的时间很短;当系统处于临界转换的时候,即系统的弹性较弱时,则从扰动中恢复所需要的时间较长。这种现象最直接的含义是,原则上可以通过研究小实验扰动下的恢复率来探测系统与阈值的距离,这是一种在不破坏系统的情况下探测系统脆弱性的方法[2]。
Veraart等通过在恒化器中培养蓝藻,使其暴露在不断增加的光的胁迫下,并定期进行轻微扰动,测量其藻类的恢复速率,最后表明测量扰动的恢复率是监测临界减速的稳健方法[20]。Nes和Scheffer也通过生态模型的计算,发现局部扰动实验的恢复时间可能是过渡前临界减速的最通用和最可靠的指标[21]。
2.1.2自相关和方差增加(Increase in Autocorrelation and Variance)
在生态系统快要接近分歧点时,自相关和方差都会增加。一个简单自回归模型就可以了解到其数学原理[22]。假设每个周期Δt之后存在状态变量的重复扰动(即增加噪声)。在干扰之前的恢复速度大约是指数型,且具有一定的恢复速率λ:(公式改编于Scheffer等人[22])
(2)
yn+1=e-λΔtyn+σεn
(3)
这里的yn是状态变量x偏离平衡点的偏差,εn是来自标准正态分布的随机数,σ为标准差。若λ和Δt都与yn无关,该模型可以写成一阶自回归模型:
yn+1=αyn+σεn
(4)
自相关系数α≡e-λΔt,当α=0时为白噪声,当α=1时为红噪声。一阶自回归模型
yn+1=c+αyn+σεn的数学期望为:
(5)
当c=0时,均值为0,方差为:
(6)
当接近平衡时,回归平衡的速度降低,恢复速率λ趋近于0+,自相关系数α≡e-λΔt趋向于1-,此时方差趋向于无穷大[22]。这就是临界减速使得方差和自相关系数增加的原因。
方差是一个重要的指标,而且它是一个简单的特征,理论上可以很容易地衡量。其中Carpenter等[23]研究在湖泊转变为富营养化条件之前,湖泊水中磷的变化越来越明显。并且模拟显示,不断上升的标准差可能提前大约十年发出早期预警信号。即使并不知道湖泊生态系统动力学的具体情况,通过研究和计算简单时间序列模型的标准差,也可以预测临界转换的到来。然而在某些特定的条件下,在稳态转换之前,方差可能会下降而不是上升。当环境因素随机波动,生态系统在临界值附近对这些因素变得不那么敏感时,或者当严重的减速降低了生态系统跟随环境高频波动的能力时,就会发生这种情况。此外,当可用数据有限时,由于接近稳态转换的低频普遍存在,方差可能被系统性地低估[18]。Veraart等[20]在指标验证实验中指出方差的检验效果不是很好,甚至呈略微的下降趋势而非升高。Dakos等[24]也认为并不是所有独立的潜在的系统转变都会表现出方差的增长。
相比方差,自相关在稳态转换时总是增加的。Liu等[25]利用DLM和相关气候数据,对美国加利福尼亚州林区的归一化差异植被指数(NDVI)进行了时变自相关估计,表明异常高的滞后一个时间步长的自相关性可作为森林死亡的早期预警信号。Cailleret等[26]也发现方差的增加可能是裸子植物死亡风险的有力预测因子。诸多的实验和模型模拟也都证明自相关是一个强有力的工具,但是自相关需要长时间且等距的时间序列,并且需要足够的分辨率来捕获系统内部动态的特征时间尺度[27]。数据的去趋势化是至关重要的,因为未经过滤的趋势可能会导致自组织模式,而这些模式可能会影响扰动系统动态响应的检测[17]。Dakos等曾使用模型来描述沙漠化过程,在其中一个模型中,放缓未能转化为方差和相关性的增加。这是由该模型产生规律性的自组织植被模式造成的[28]。这一发现表明了作为临界转换指标的方差和自相关的局限性。虽然临界减速在稳态转换时可能是一种普遍现象,但在特定系统中通过间接指示器检测它可能有一定的条件限制。
2.1.3条件异方差
条件异方差是指一个时间步长的方差与一个或多个先前时间步长的方差呈正相关关系。这表明高变化期将倾向于紧随高变化期,而低变化期将倾向于紧随低变化期。因为临近转变的时间序列的部分将呈现为高变异性的集群,而远离该转变的时间序列的部分将呈现为低变异性的集群,所以条件异方差可以作为稳态转换的预警信号。条件异方差基于拉格朗日乘数检验,该检验首先将拟合模型的残差提取到时间序列来计算,然后对残差进行平方,最后将残差本身回归滞后一个时间步长,滞后残差的线性回归的正斜率表明条件异方差[29],其一般用于模型模拟研究。
2.2 空间性指标
2.2.1空间相关性
对于具有多稳态的生态系统模型的空间单元,如果假设每个单元的条件不同,那么单元之间的扩散交换将不断地趋向于减小单元之间的这种差异。准确地说,由反应部分(F)和扩散部分(D)控制的两个相邻单元(x1和x2)之间的运动,其方程[18]为:
(7)
(8)
式中,pi为两个单元之间的异质性的参数,而c是驱动系统到临界点的控制参数,计算其雅可比矩阵的特征值。当系统远离临界点时,单元由“反应”和“扩散”过程支配。当系统靠近临界点时,因为临界减速现象,“反应”单元会变得很小,而“扩散”单元与稳态转换的距离无关,仅取决于两个单元之间的梯度。此时“扩散”将占主导地位,从而使两个单元之间的差异相等。导致一个单元中的状态将强烈依赖于其邻居的状态,所以在接近过渡的时候,单元之间将会有更强的相关性[18]。
空间相关性的指标包括空间方差和空间自相关,Donangelo等[30]用空间湖泊模型来研究湖泊富营养化,并且表明空间方差的增加可以作为湖泊从贫营养状态转变为富营养状态的预警信号,其单变量或多变量可观测的空间方差在作为稳态转换的预警信号要优于时间方差。Kéfi等[31]利用元胞自动机模型研究了干旱生态系统的荒漠化过程,表明放牧压力下植被斑块大小分布的变化可能是荒漠化开始的预警信号。在由许多耦合单元组成的系统中,增强的空间相关性可以作为更强大的预警信号。如果系统中有足够的空间异质性和连通性,这种空间相关性的增加代表了比时间序列的指标更好的早期预警信号。空间相关性通常要优于时间序列的预警信号指标,这证实了空间指标可能比时间指标更可靠的说法[32]。空间方差和空间自相关的空间模拟在这些类型的空间扩展系统中表现更好[30, 32-33]。
但是这些空间预警信号有3个主要缺点。首先,它们在很大程度上依赖于状态变量与其空间均值的偏差。因此,当系统接近稳态转换时,这些指标受到不同位置的状态变量的时间均值的变化的影响,这很可能发生在不同的异质系统中。其次,这些方法一次只查看一幅影像,因此限制了它们的信息收集能力。例如,仅使用单幅影像很难识别与稳态转换相关联的时间模式[34]。而且空间相关性也可能触发假警报,如果环境中的异质性随着时间的变化或小规模干扰增加景观的不均匀性,这可能导致空间相关性的增加,从而产生对即将发生的变化的假警报[32]。
2.2.2空间异方差
空间异方差检验首先对数据进行过滤,然后再使用回归来检验过滤值平方之间的自相关,自相关采用Moran′s I指数计算。空间异方差检验可以促进使用空间显性数据,因为它对边缘的响应很强,特别是在干旱系统的荒漠化方面,因为在干旱环境中,空间相互作用在构建生态系统格局和过程中至关重要[35]。而且空间异方差与临界减速之间没有直接的关系,而是对空间变异的聚集性做出响应[36]。此外,空间异方差只需要少量的时间序列数据,并且这些时间的间隔不必有相等的间隔,所以非常适合时间尺度较长的陆地植被系统[25,33],尤其是有观测误差和环境特征梯度的实测数据,这一指标很有效。空间异方差也会受到限制,因为这种分析通常不能确定观测数据中观察到的模式是由于稳态转换的生态系统状态,而不是其他机制。除此之外,它还可能会降低具有高环境噪声(观测误差)的生态系统参数的有效性[35]。
2.3 非临界减速指标
2.3.1偏度
偏度是多稳态理论中的不对称指标,受外部大波动和景观非对称性对生态系统动力学的影响。偏度信号利用了生态系统的两个普遍特征:非线性和大的外部波动。无论是弹性降低(动力学参数发生变化)还是外部波动增加(外部噪声部分的宽度增加),都可能发生稳态转换。偏度(无量纲)可用τ来表示,其公式[37]如下:P(x)为概率密度函数,μ为均值,σ为标准差:
(9)
式中,P(x)为概率密度函数,μ为均值,σ为标准差。当系统远离稳态转换时,偏度为零,偏度越大则越靠近稳态转换。根据所考虑的生态系统的不同,变化可以由零偏斜到正值或负值,或者从一种偏斜情况到另一个偏斜情况。Guttal和Jayaprakash利用模拟模型和对撒哈拉气候突变的实测数据进行分析,发现偏度作为稳态转换的指标是可行的[37]。
2.3.2闪烁
闪烁是指当系统进入导致分歧的双稳态区域时,如果随机作用足够强,开始在两个可选择的吸引盆地之间来回摆动[38]。这也是一种早期预警信号,因为如果潜在的条件变化持续存在,则系统很可能永久地转移到另一种稳态当中。闪烁的一种特殊情况称为随机共振,如果随机扰动与缓慢的周期变化相结合,就会在最终转变到另一种稳定状态之前发生一系列周期性转变[2]。统计上,可以观察到状态的频率分布中的闪烁,表现为方差和偏度的增加以及双峰(反映了两种可选的状态)[39]。
Wang等[40]结合了来自湖泊流域系统的经验数据和数学模型,表明可以从稀疏的数据中检测到闪烁,并且如果系统受到足够大的外部影响,闪烁将成为更强的系统转变的早期预警信号。Dakos等通过湖水富营养化模型,比较了基于临界减速指标和闪烁的区别,发现闪烁系统在许多方面与稳态转换的经典图像不同,但其动态变化可能会携带即将发生稳态转换有价值的信息[41]。
2.4 生态功能指标
交互网络可以成为植被变迁的预警信号[42];生产力动态的变化可以成为生物群落尺度上的生态系统状态转换的预警信号[43];干旱化的加剧可以成为多种生态系统属性发生突变的预警信号[44]。植物特性的变化可以使得临界点更早的到来,也会改变恢复的路径[3]。基于生物量数据的统计矩阵和体型动态的预警信号可以预测复杂系统的恢复,将这些信号组合成一个单一的度量可以产生最可靠的恢复信号[9]。与协方差矩阵的最大特征值相关的特征向量有助于识别最易受临界转变影响的区域,因为它只考虑了系统平衡的波动,从而消除了平衡值的任何变化的影响[34]。
3 不同尺度下生态系统多稳态检测
在不同的尺度下可以发生不同稳态的转换。在种群尺度上,恒化器中培养蓝藻,蓝藻会随环境的变化迅速转移到不同的稳态,而且可以采用实验来测量各个指标[20]。而在群落尺度上,可以从物种和群落水平上测量生物量的时间序列,特有种可能是检测群落发生稳态转换的最佳指示物种,将临界点理论和网络结构模式相结合可以得出的临界减速指标[45]。在生态系统尺度上,不同类型的生态系统在不同时间尺度上的稳态转换也是不同的。在湖泊生态系统中,在日尺度上,浮游生物群落可能会迅速转移和循环;而在多年尺度上,整个生态系统可能由稳定状态突然转化为持续的浑浊状态。此时湖泊稳态的转化也可以通过检测食物网的重组过程来预测[46]。在海洋生态系统中则不能通过相关尺度的实验来探索。在地球系统中,像厄尔尼诺、PDO等气候事件几年就会发生一次;而在更长时间尺度上沙哈拉沙漠的诞生都是突然的稳态转换[47]。在对大尺度系统的研究时,要通过模型模拟来完成,也必须依赖对自然实验效果的解释,对其进行参数的调整。所以尺度越大、系统越复杂,我们对稳态转换背后的机理的了解就越匮乏,预测也就越困难如图4所示[2]。
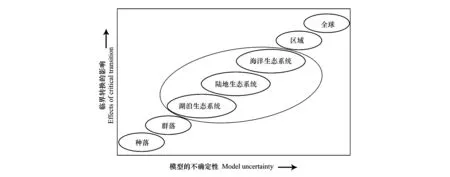
图4 尺度对模型和稳态转换影响(图片改编于Scheffer [2])Fig.4 Effect of Scale on Uncertainty of the Model and Critical Transition
3.1 种群和群落尺度
(1)小型模拟实验
在种群尺度上,可以通过设置小型的模拟实验,通过改变外部的环境状况使系统发生稳态转换,采用观测值进行各个预警指标的计算,从而预测或验证多稳态理论。Veraart 等通过在恒化器中培养蓝藻,使其暴露在不断增加的光胁迫下,并定期进行轻微扰动,通过测量光衰减率来衡量蓝藻生物量,并间接测量蓝藻的恢复速率。这个模型显示了多稳态和临界点,蓝藻为自己的生长提供了所需的荫蔽创造了正反馈,但太高的光强对初级生产力是有害的,这构成了双稳态背后的机制。最后的实验结果表明,即测量扰动的恢复率是监测临界减速的稳健方法[20]。
(2)网络结构模式
在群落尺度上,可以从物种和群落水平上测量生物量的时间序列,特有种可能是检测群落发生稳态转换的最佳指示物种,将临界点理论和网络结构模式相结合可以得出的临界减速指标。Dakos和Bascompte[45]使用79个互利共生网络的结构来模拟一个环境逐渐变化的场景,该场景第一次导致一个物种的突然灭绝事件,然后是一系列物种的灭绝,直到群落完全崩溃。其结果表明,首次灭绝的物种拥有最强的变异系数,互利共生网络的嵌套结构会增加稳态转换的概率。
3.2 生态系统尺度
(1)大型模拟实验
在湖泊生态系统尺度上,在日尺度上,浮游生物群落可能会迅速转移和循环;而在多年尺度上,整个生态系统可能由稳定状态突然转化为持续的浑浊状态。此时湖泊稳态的转化也可以通过检测食物网的重组过程来预测。Carpenter等在3年多的时间里通过逐渐增加了顶级捕食者来破坏湖泊的食物网,与另一个相邻的湖泊作为参考生态系统被同时监测。在食物网完成稳态转换前一年多的食物网重组期间,在实验组的湖泊中,稳态转换的预警信号是明显的[46]。
(2)脆弱性结构分析
在生态系统尺度上可以采用脆弱性的结构进行分析,虽然生态系统的复杂性阻碍了精确的机械建模,但某些特征是脆弱性的通用标志,这些标志通常出现在一些稳态转换之前。这类系统的总体响应有两个主要特征:组件的异质性和它们的连接性,这些特性如何影响稳定性取决于网络中交互作用的性质。单个节点的响应异质性和低连接度可能会导致整个网络响应环境变化是渐变的而不是突变的。这是因为相对阻力较小,所以不同的节点将会逐个转移到另一种状态。相比之下,同质性的节点和高连接度的网络可能会使得稳态转换出现阻力,直到临界点时,其所有的节点都同步移动到另一个状态[38]。
3.3 区域尺度
(1) 空间代替时间
在有一定梯度的区域可以采用空间替代时间的方法,而且要想用空间代替时间的方法找到稳态转化的点需要比较苛刻的条件:首先需要一个环境梯度,例如我国的内蒙古地区的草地生态系统的演变是由降水梯度所控制的,并且纬度基本一致,就可以建立一个天然的实验场。基于这个梯度就可以假设如果将来的降水增多,那么荒漠草原就可能会转化成典型草原,典型草原就可能转化成草甸草原;如果将来的降水减少,那么典型草原可能会转化成荒漠草原,草甸草原可能会转化成典型草原。Hu等人基于中国内蒙古地区由东到西降水梯度递减的事实,通过变异系数、不对称性和对降水敏感性这3个指标,量化了地上净初级生产力(ANPP)的动态空间格局,并把干旱荒漠草原、半干旱典型草原到半湿润草甸草原的过渡联系起来。其表明这3个ANPP动态指标均表现出明显的空间分布特征,并呈现出草地类型间的峰值信号转换。因此,监测ANPP动态变化有可能预测未来生态系统状态的变化。但是,使用空间代替时间的方法来预测生态系统稳态转换对气候变化的响应是具有局限性的。例如在土壤属性上,气候变化、植被动态和响应之间存在时空不一致性[43]。
(2) 卫星遥感监测
在区域尺度上可以使用卫星遥感监测的方法来验证或预测稳态的转换。具体有两种方法:第一类是遥感影像目视解译与计算机自动分类相结合的方法;第二类是基于遥感或其所衍生的指标,利用数理统计进行时间序列对比分析[8]。例如Hirota等利用 MODIS(Moderate Resolution Imaging Spectroradiometer)的植被覆盖度产品数据揭示了非洲、澳洲和南美洲的热带和亚热带地区存在着三种稳态,即森林态、稀树草原态和无树态[48];Miller分析了美国科罗拉多高原半干旱区草原的地面样方数据,发现该区存在生物结皮-多年生草地态、裸地-多年生草地态以及裸地-单年生草地态[49]。Berdugo等结合地面样点数据和来自 Google Earth的高分辨率卫星数据,发现旱区在全球尺度上存在着高功能区和低功能区两种状态[50]。
3.4 全球尺度
全球尺度的研究大部分采用模型模拟或者全球实地调研和遥感观测相结合的方法。在古气候变化和物种大灭绝的研究中,最好的方法就是利用模型模拟古气候事件发生的时间序列,再进行稳态转换的验证。古气候突变事件包括:大约5000年前,撒哈拉突然从一个有湖泊的植被繁茂的区域变成了沙漠[47];大约1.3万年前,由于流入大西洋淡水的减少,导致温盐环流的骤停,使得格陵兰岛的气温骤降超过10℃,将该地区推回到几个世纪以来的冰河时代[51];以及历史上五次著名的物种大灭绝。Dakos等从地质记录中重建了8个古代气候突变的时间序列,采用了简单的气候模型来模拟地球从温室到冰球的转变;温盐环流模型,模拟冰河时代;北非沙漠化气候模型,模拟了撒哈拉沙漠的诞生。通过模拟的数据分析了它们的自相关变化,表明在气候变化之前就已经有了波动开始减缓的特征,用数学方法证明了自相关增加是临界点的标志[17]。全球实地调研和遥感观测相结合做的很好的研究有Berdugo等调查了20个生态系统结构和功能属性对全球干旱的响应,表明干旱化导致多种生态系统属性发生系统性的突变。这些变化依次发生在干旱值为0.54、0.7和0.8时,表现为植物生产力、土壤肥力、植被盖度和丰富度的急剧下降[44]。
总而言之,定量预测仅限于较小尺度的系统,这种系统可以观察到多稳态并从实验上探索临界点。比如我们可以知道,在一个淡水湖中当磷负荷在什么时候可能会变成浑浊的状态。在较大尺度的预测则比较困难,比如我们很难知道亚马逊热带雨林被烧掉多少之后会不可逆转的转化为稀树草原状态[2],所以在较大尺度上多采用模型模拟的方法来找到临界点。而且生态系统每增加一个单位面积,该系统崩溃所需的时间单位就会越来越小,这意味着大系统往往比小系统移动得更慢,但速度却不成比例[52]。
4 问题与展望
4.1 主要问题
4.1.1多稳态的机理研究不足
目前仍无法探测一个系统究竟存在哪些稳态,在什么条件下会转变成另一种稳态。现有的多稳态模型是否可以起到通过树木看到森林的作用,还需要通过模型和实验相结合的方法来进行验证。多稳态的突变推动机制一般为正反馈机制,是否还存在其他的推动机制值得深入探讨。性状动态的变化表明稳定性的丧失,还是对新的当地条件的适应也是值得思考的,因为性状分布的变化可以表明群体处于压力之下[53]。表型转变之后可能会出现突然的种群或群落范围的崩溃[54],或可以使种群应对不断变化的环境条件,而不会出现丰度的显著下降。虽然群落水平的稳定性的丧失仍可能通过改变个体大小的种间相互作用的强度来传播。但是在某些条件下,例如当现有的遗传变异耗尽时,性状反应可能不会领先于即将到来的群落崩溃[11]。因此,量化基于特征的早期预警信号,从而减少预测的不确定性,是值得深入研究的。
4.1.2数据收集和处理问题
在实践中,要想检测到时间序列的预警信号,实时检测可能来得太迟而无法采取行动,因为高质量和高分辨率的时间序列的数据很难快速获得[22,55]。如何克服数据质量差的影响也是一大难点,例如,抽样中人为因素的作用。以及如何消除噪音,目前在多稳态模型计算中的一个重要假设是模型系统的随机动力学是生态数据的完整表示。而真实的生态数据容易产生多种误差源,包括外部误差和观测误差,以及识别和模拟动态过程所涉及的不确定性[38]。一些观察到的时间序列中的不对称性变化不一定意味着接近临界转换,因为它可能不是由于内在动力学而是由于外部噪声中的不对称性。现在普遍的做法是使用高斯白噪声或零模型来模拟自然的误差,但是这些方法过于简单并不能很好的消除相关趋势。所以,用适合的噪声来消除嘈杂的外部变量,将真实的系统变化与其他的噪声源分离是需要深入研究的。
4.1.3多稳态的尺度推绎问题
在多稳态的研究中,不同尺度系统有着不同的研究方法。例如,定量观测实验适用于小尺度的研究,而在较大尺度的研究则一般采用简化的模型来模拟研究。在空间上,选择正确的尺度有可能极大地改变空间信号的可靠性[56],而在时间序列中,时间尺度对于系统进程速率有重要的影响[53],了解尺度对这些信号可靠性的影响,从而选择正确的尺度来应用这些方法是至关重要的。一些预警信号往往涉及的特定机制而不能推广到其他系统[57]。如何将小尺度的方法和结论推演到更大的尺度上,如何在较大尺度上设计实验来校正模型的参数,以及不同尺度的预警信号是否可以通用,仍需进一步的研究。
4.1.4预警信号问题
在稳态转换的预警中,经常会出现两种错误。第一类错误是如果系统灾难性转换正在到来,但是预警信号却没有显示,所以我们没有做出预警。此时我们就要承担稳态转换之后的后果,例如湖泊变浑浊后的治理和草原变成荒漠的治理。这可能因为在系统突然转变之前,没有渐进地接近临界点。例如,信号可能保持在与临界点一定的距离,但被罕见的极端事件驱动到另一个稳定状态[22]。如果外部扰动机制随着时间的推移而改变,就会出现统计困难,这可能会扭曲或抵消预警信号。
第二类错误是如果系统的灾难性转换还没有到来,但是预警信号却显示临界点即将到来,于是我们做出了预警。此时我们就需要承担过度反应所承担的损失,例如过度抑制放牧来阻止荒漠化的过程,这虽然使得草原适当生长,但是减少了食草动物的数量,损害了放牧者的利益,使得整体的生态系统服务价值没有达到最大。第二类错误可能是由于预警信号的统计困难造成的。例如,增加的自相关的检测可能需要长时间序列[58]。如果假想的预警信号不是接近稳态转换的结果,就会出现误报。这可能是偶然发生的,也可能是由于系统内部的混乱趋势或外部扰动造成的[22]。
在到达临界点之前,扰动通常会触发一个转变。因此,尽管稳态转换指标的趋势可能起到警示作用,但转化的实际时刻仍然很难预测。当在实际应用时,一个关键问题是信号是否能够足够早地被检测到,以便采取行动为防止系统发生稳态转换。当涉及到即将到来的稳态转换中做出决策时,我们面临着巨大的不确定性。这种不确定性源于多种因素,包括预测不准确、数据不足和隐藏的非线性,以及对风险的感知和容忍度的特殊性,所以如何提高预测的准确性,量化正确的预警信号的概率,并对错误预警信号的比例进行加权是避免犯这两类错误的关键。
4.2 未来发展方向
4.2.1基于生态系统恢复实践不断完善多稳态理论
系统在发生转化之后,其结构和功能与之前的系统往往有很大的差别,内部的机制也发生了很大的变化。如果要想恢复到原来的状态,仅仅重建系统到崩溃之前是远远不够的,就像放牧模型一样,因为过度放牧使得草原生态系统转化为荒漠生态系统,通过减少放牧的数量并不能使荒漠生态系统恢复到草原生态系统,而需要做更多的努力,例如改善土壤的有机质含量、人工灌溉等等,才有可能恢复到之前的状态。加强生态系统修复实践的结果和数据的收集,将其应用在多稳态理论研究系统的临界点以及反馈控制机制的完善上,是重点的研究方向[12]。
基于生物量和特征的指标具有广泛的普适性,因为它们几乎没有对系统的基本结构和功能做出假设,因此可以为退化系统的恢复提供一个关键工具[9]。此外,现有的生态系统复原理论只考虑了超过临界点的外部条件变化引起的稳态转换,因此缺乏进化观。而外部条件的变化对生态影响很小,也不会将系统推到临界点之外。因此,这种变化不会立即引起稳态转换,而是会触发一个进化过程,使表型性状超过临界点,从而导致(在经历了相当长的延迟之后)由选择引起的稳态转换,所以将进化动力学纳入生态系统多稳态检测是未来研究的方向之一[59]。
4.2.2多尺度下的生态保护管理和政策实践
将多稳态的理论运用在生态保护管理实践中,是多稳态理论运用的重要部分。可以通过分析不同尺度临界点的计算和研究方法,并根据现有数据了解这些方法的局限性和优势,不断完善多尺度分析方法,更好地服务于设计目标。这其中的一个关键部分是将这些预警信号与决策框架[60],资源有限保护的现实[61],利益相关者和管理者之间的冲突[62]和适应性管理战略[63]适当地联系起来[11]。建立和保持预期生态系统状态的弹性可能是生态系统管理最实际和有效的途径。鉴于生态系统和环境变化的复杂性,如何将早期预警信号整合到最优决策理论中,是生态保护研究的重要内容[63]。此外,如何更好的将多稳态的理论和生态保护管理和政策相结合,是未来研究的重点[64]。
4.2.3时间和空间预警信号的综合运用
将来自统一数据的多个信号组合在一起,可能会提供更可靠的稳态转换的信号,例如把空间和时间预警信号以及性状动力学等相结合[19,65]。因为一般的预警信号,如时间序列增加的自相关性和方差,检测这些领先指标的滞后时间通常很长,而空间模式可能提供更强大的领先指标,因为它们比时间序列中的单个数据点包含更多的信息[30],但是空间指标很难识别与稳态转换相关联的时间模式,而且对于具有自组织图案形成的系统,存在特定的信号,所以将多个指标相结合可能成为更准确的稳态转换的指标。然而,目前在这一领域已经使用了非常有限的相对简单的方法,例如多个统计信号的归一化和求和[19,54]。更复杂的统计工具,如时间序列的多变量分析或机器学习算法,提供了简单模型选择框架的扩展,以及各种信号之间的互相关系[66]。所以将多元的预警信号整合到一起从而提供更准确的预测,是未来研究的方向。
致谢:张雪莲女士绘制本文图形,特此致谢。
