信鸽外纹体神经元对习得性目标认知的响应特性
2021-10-08牛晓可孔德铭王治忠乔长城
牛晓可, 孔德铭, 王治忠*, 乔长城
(1.郑州大学电气工程学院/河南省脑科学与脑机接口技术重点实验室, 郑州 450001; 2.中国电子科技集团公司第二十七研究所, 郑州 450001)
脑认知作为21世纪新兴研究门类,是探究大脑工作机制的重要一环,已成为全世界科技竞争的重要关注点之一。近十几年脑科学的实验研究表明认知训练可提升大脑功能,改善阿尔茨海默症等众多脑疾病症状,对于医学史的发展具有重大的意义[1]。认知是指生物通过感知器获取环境信息、知识的智能加工过程,具体包括知觉、注意、学习记忆、思维、决策、兴趣、执行能力等。习得性认知主要指生物在后天生活中由学习和经验得到,通过某种反应得到强化形成一种条件反射的认知过程。
Hossein等[2]通过设计人与非人的真实世界物体图像对鸽子进行习得目标认知行为训练,结果表明,对于不同目标,鸽子外纹体神经元响应特征可以表征这些视觉信息。万红等[3]对信鸽进行十字迷宫训练,提取了有效的神经元信号局部场电位(local field potential,LFP)能量特征,准确地解码出信鸽习得性认知后的行为信息。杨莉芳[4]对鸽子进行认知学习训练,结果表明,鸽子在习得性认知前,其脑间LFP能量特征同步性在视觉刺激期间无明显变化,经过长时间的认知学习,对于一个习得性认知视觉刺激,其脑间能量特征的同步性显著提高。李晓媛等[5]对小鼠进行了视觉认知训练,结果表明,大鼠对恐惧图像产生认知后其相关认知脑区神经元集群的锋电位(Spike)活动相较认知前发生改变。刘殿玮[6]报道了经过记忆训练后,脑源性神经营养因子在与认知功能密切相关的纹状体区等表达较高。以上实验研究表明认知相关脑区的神经元响应在习得性认知训练后都发生了显著的改变,然而这些研究都以哺乳类动物为研究对象,虽然鸟类属于与人类亲缘关系较远的类群,但是随着大量的研究已经证明其认知能力也十分出色[7]。目前却少见关于鸟类在习得性目标认知前后相关脑区神经元响应会发生哪些变化的文献报道。
鸟类卓越的目标认知能力与其视觉通路的独特结构与功能息息相关,主要以离顶盖通路较为发达,由视网膜-顶盖-圆核-外纹体等组成,其中外纹体与哺乳类外纹状体皮层具有同源性[8-9],具有较大的神经元感受野,通常可以延伸到对侧眼睛的整个视野。已有研究者[10]证实了鸽子外纹体神经元与奖励刺激之间存在一定的关系,证明了信鸽外纹体神经元会根据认知学习中的任务奖励刺激改变其活动模式。Scarf等[11]证明了外纹体神经元集群对多种刺激都有强烈的反应,但在处理不同的刺激时,它们的Spike序列发生了改变。此外,Johnston等[12-13]证明了信鸽外纹体神经元能根据奖励相关的任务偶然性来改变它们的活动模式;并证明外纹体神经元参与视觉特征信息处理过程的同时对奖励信息也有表征,且发现鸽子在执行视觉认知训练过程中左脑具有优势。根据已有研究结论可推测,鸟类外纹体神经元会根据相关任务和任务奖励改变响应活动并参与视觉记忆工作,但并未给出鸟类对目标认知过程中神经元信号响应规律研究的定量分析。
现以信鸽为动物模型,对其进行特定目标认知训练,采用多通道金属电极阵列同步采集不同认知阶段外纹体神经元电活动,分别提取Spike发放率特征、LFP伽马子频带能量特征,并进一步计算不同试次神经元响应间的同步性以及神经元间的互信息量,将认知前后的这些特征进行对比,结果表明以上特征均在目标认知前后发生了显著变化,该研究是对现有研究的有效补充,对于深入理解鸟类高级视觉皮层对目标认知过程的信息处理机制具有重要意义。
1 方法与材料
1.1 实验材料
选用4只体重为 300~400 g的健康成年信鸽,雌雄不苟。采用单笼配置,每天给予动物光照、黑暗各 12 h,动物房温度保持为(23±2) ℃,饲喂过程中,无限制供应饮水,为了保持鸽子在训练过程中能够持续执行任务,限制其饮食(体重维持在正常体重的80%左右),并使其保持饥饿状态,饲养和照料标准均遵循《郑州大学实验动物管理规定》。
1.2 训练方案
选择“Y”字形训练箱对信鸽进行习得性目标认知训练,当信鸽行至分岔口处(有玻璃门挡住前进路线,可透过玻璃门看到两条臂端的显示屏),位于臂端的显示器上随机显示目标图片或背景干扰图片,持续3 s。3 s后玻璃门自动打开,鸽子选择一条臂行走,如果选择了特定目标所在的臂,则弹出食盒给予奖励[图1(a)],反之则不给[图1(b)],图1给出训练过程中两个不同试次,其中,0点表示目标呈现时刻。
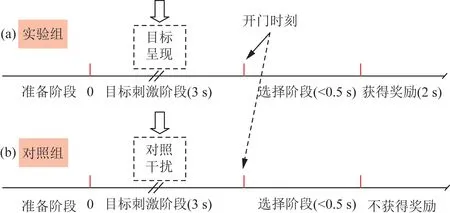
图1 信鸽训练期间不同试次的事件序列Fig.1 Event sequences of different trials during pigeons training
训练分为预训练和正式训练,其中预训练为信鸽熟悉训练箱的过程,有时需要研究人员手动投食引导信鸽获得奖励,正式训练前期阶段,信鸽不能正确建立认知目标与奖励的联系,表现在行为抉择正确率低于80%,经过一段时间的认知学习,信鸽行为抉择正确率有较显著提升,稳定3 d达90%以上,即判断信鸽对目标建立习得性认知并处于认知后阶段。选取灰色背景下移动的特定动物图片为信鸽认知的目标,以灰色背景图片(或其他干扰目标)作为对照。
1.3 电极的植入与信号的采集
电极植入预训练阶段(熟悉训练箱的阶段)之后正式训练之前进行。手术时,在信鸽腹腔注射戊巴比妥钠(1.5%, 2.5 mL/kg),将处于麻醉状态信鸽俯卧固定在脑立体定位仪中。由于信鸽在执行视觉认知训练过程中左脑具有优势,因此在信鸽左侧脑区确定植入中心点位置(前/后±9.25 mm, 左/右 ±5.00 mm),脑区定位坐标根据鸽脑立体定位图集选择,植入16通道(4×4)不锈钢微丝电极阵列后利用牙托水泥固定,完成手术后的前3天在伤口周围涂抹抗生素(红霉素软膏)以防感染,信鸽需一周恢复时间,期间提供充足的饮水和食物。
训练过程中使用Cerebus 采集系统(美国Blackrock公司)以30 kHz采样率进行电生理信号的同步采集。在完成所有的记录和行为实验后,对电极记录位点进行电损伤(2 mA, 30 s)以标记电极在鸽脑中实验记录位置[14]。进行灌流取脑,将脑从颅骨中取出并在30%浓度蔗糖溶液中进行脱水处理,在多聚甲醛溶液中固定48 h。从记录部位收集冰冻的冠状脑切片(30 μm),并用甲苯胺蓝染色(Sigma-Aldrich, Saint Louis, USA),对记录电极在大脑中的位置进行了组织学检查,脑立体定向图(A/P 8.75 mm)中所绘与脑切片均为信鸽左脑,如图2所示。
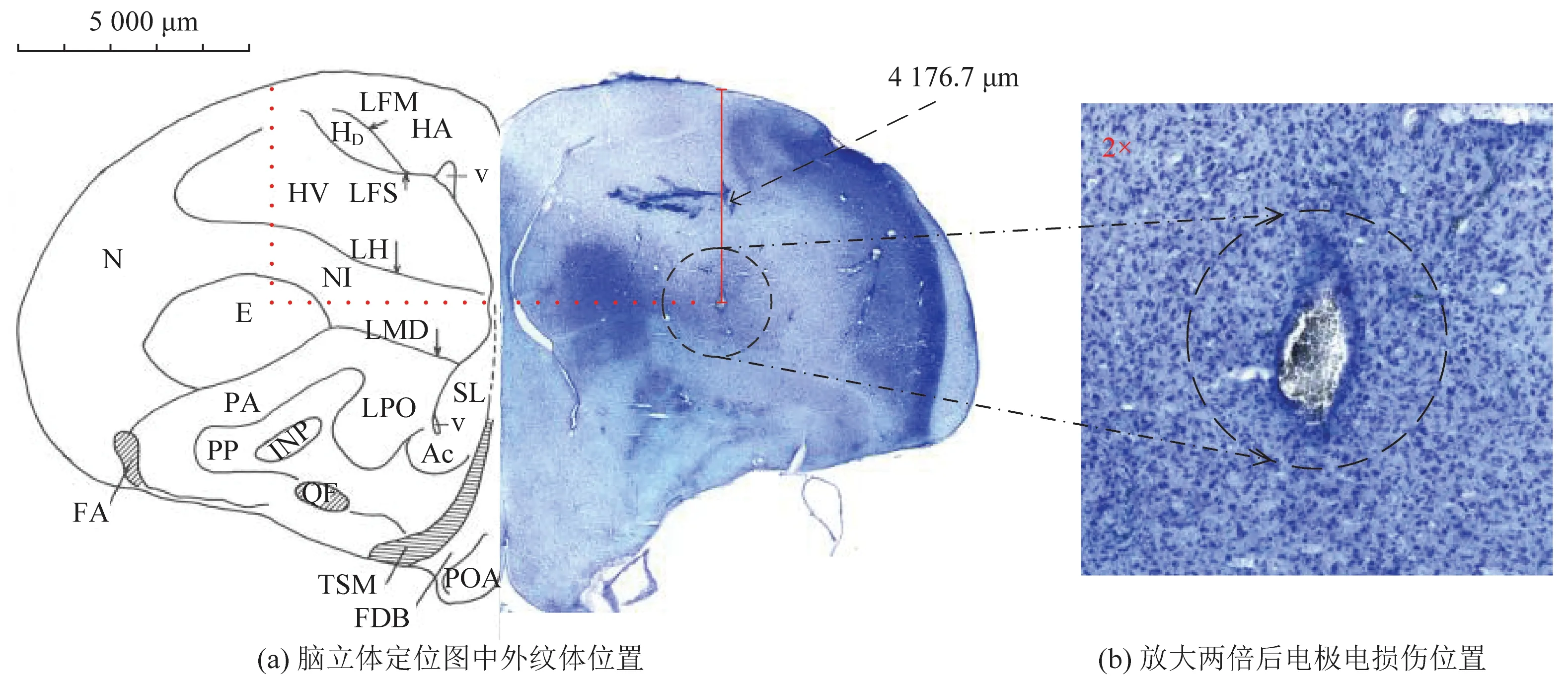
E为外纹体(ectostriatum);Ac为伏核(nucleus accumbens);CO为视神经交叉(chiasma opticum);FA为额叶束(tractus fronto-archistriatalis);HA为纹状体副束核(hyperstriatum accessorium);HD为背侧纹状体(hyperstriatum dorsale);HV为上纹体腹核(hyperstriatum ventrale);INP为脚内核(nucleus intrapeduncularis)图2 鸽脑立体定位图中外纹体所处位置与记录电极电损伤标记位置Fig.2 The location of ectostriatum in stereotaxic atlas of the pigeon brain and the marked location of the recording electrode with electrical damage
1.4 神经信号处理和响应特征提取
在实验过程中,由于数据采集线或者电极可能会因为外界因素而造成损坏,产生坏道,因此,需要对坏道进行剔除。采集的信号包括高频的Spike(250~5 000 Hz,Butterworth带通滤波器) 和低频的LFP(0~250 Hz,Butterworth低通滤波器)。
1.4.1 针对Spike信号响应特征提取
首先,对Spike信号进行预处理操作[15],由于电极尖端检测到的信号通常是一个或多个神经元的放电信号,因此,采用小波变换和聚类分析法来实现对神经元Spike信号的分类,在以阈值为基础的Spike检测中,还会引入一些大幅值干扰,因此,采用共同平均参考方法去除大幅值噪声干扰。
然后,提取神经元的Spike信号发放率特征,根据训练过程可知,目标呈现的3 s是鸽子观察两个屏幕的时间段,同时为了对比信鸽观察目标前后神经元响应的差别,因此这里重点分析目标呈现前1 s到开门后1 s(鸽子做成决策后)共5 s内的响应。计算方法如下:设定时间窗口大小为Δt=50 ms,计算窗口中每个神经元Spike发放的个数,设发放个数为fij,表示第i个神经元在第j个时间窗口内的发放数。发放率矩阵为Rt=[Rt1,Rt2,…,Rtj],对其进行归一化处理,并使用大小为15的Gaussian窗降噪平滑曲线。

(1)
进一步地,计算了不同试次发放率曲线间的Spearman秩相关系数,用于衡量不同试次间神经元响应间的可靠性,计算公式为

(2)
式(2)中:E为变量X和Y间的期望;μX为变量X的均值;μY为变量Y的均值;cov(X,Y)为变量X和变量Y之间的协方差;σX为变量X的标准差;σY为变量Y的标准差。Spearman秩相关系数能够反映两组信号之间关系的密切程度,越接近1表明越相关,说明神经元响应越趋于一致。
1.4.2 针对 LFP信号的响应特征提取
首先,对LFP信号进行预处理操作,采用基于最小均方(least mean squares,LMS)算法的自适应滤波器[16]来去除信号中存在的50 Hz工频干扰,采用共同平均参考法有效地去除由于鸽子自身运动所产生的电极间相关噪声。
然后,分析提取的5 s内神经元LFP信号的能量密度,采用多窗口方法(multitaper method,MTM)估计LFP信号的频谱[17],并进一步分析训练前后LFP信号主要频率段的能量密度变化。其基本思想是利用离散扁球序列(discrete prolate spheroidal sequence,DPSS)的多个正交的基函数(tapers)与原始数据相乘,并进行傅里叶变换后得到数据集,再估计其谱分量,计算方法如下。
假设一个离散时间序列xt,其中t=1,2,…,N,对应的频谱为
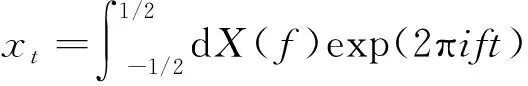
(3)
式(3)中:X(f)为傅里叶变换后的序列;f为频率。其频谱可通过式(4)进行估计:
(4)
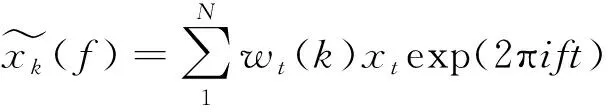
(5)
式中:xt为离散时间序列;wt(k)表示K个长度为N、带宽为W的第k阶DPSS基,其中k=1,2,…,K。选择7个正交的DPSS基函数估计LFP信号的能量,设时间带宽乘积NW=4。 将该5 s内LFP信号能量记为Pt,则能量矩阵为Pt=[Pt1,Pt1,…,Ptn],其中,Ptn为第n组实验响应信号的能量。
1.4.3 两两神经元间的互信息量分析
进一步分析了信鸽信鸽外纹体两两神经元响应信号的标准化互信息量,用于探究两两神经元间携带的信息量是否具有相似性。计算方法如下。
对于离散随机变量X、Y,有
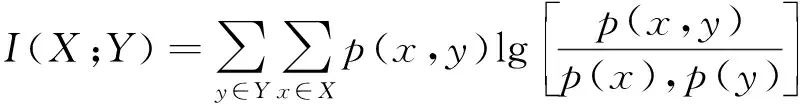
(6)
式(6)中:p(x,y)为变量X、Y的联合概率分布函数;p(x)和p(y)分别为变量X和Y的边缘概率分布函数。
两两神经元间的互信息量反映了一个神经元包含的另一个神经元信息的多少,标准化互信息量越接近1,两两神经元之间携带相似的信息就越多,说明集群携带的信息越冗余。
2 结果与分析
共记录了4只信鸽(A1、A3、B66和B93)连续15 d的行为训练和神经响应数据(分别选取了4、5、5和8个神经元),每只信鸽每天采集10~16个试次。下面主要以其中一只鸽子(A1)为例,给出一个经典神经元响应特征的结果,以及所有神经元的统计结果。
2.1 行为认知结果
根据训练正确率(一天中训练成功的试次占全部训练试次)将正式训练分为三个阶段,分别为习得性认知前阶段、认知中阶段和认知后阶段。图3给出了4只信鸽为期15 d的训练结果(其中认知前阶段持续3~4 d,认知中阶段持续2~3 d,认知后阶段持续8~10 d)。
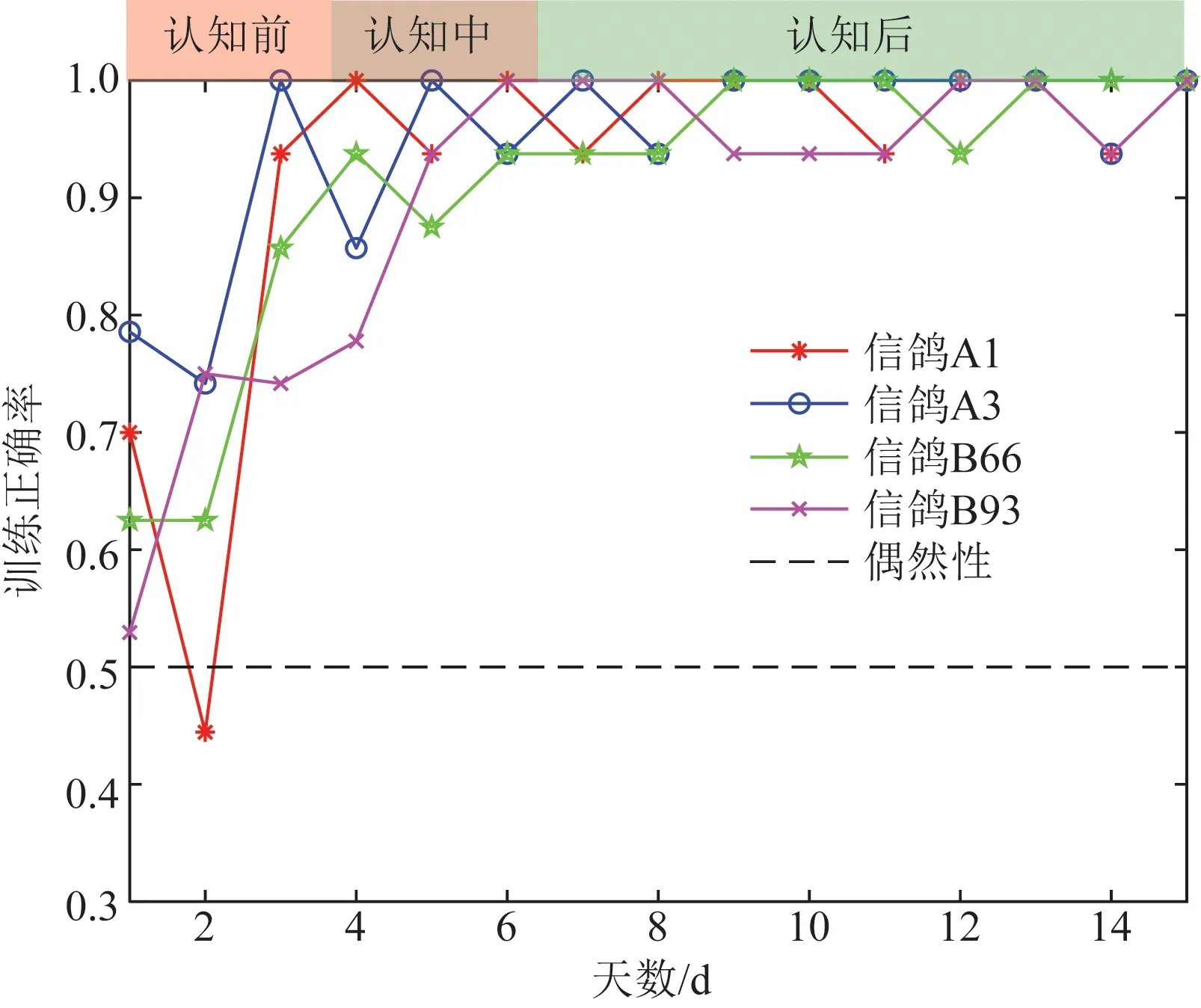
图3 不同阶段训练正确率Fig.3 Training accuracy at different stages
由图3可以看出,随着信鸽训练程度的提高,训练正确率也逐渐提高,达到较高水平并维持稳定,认知后阶段信鸽表现出色,可通过快速扫视屏幕识别目标,并选择正确的奖励区域获得奖励,但从图3中可以看出,在训练前阶段正确率已经普遍高于基线水平,出现这种情况的原因正式训练前的预训练阶段采用人工诱导的方式建立奖励和图形之间的认知关联,这个阶段鸽子可能已经初步建立了认知,但是由于加入了人工因素未纳入统计,因此,进入正式训练后信鸽对于这个训练模式并不是完全陌生的,只是正确率不是很稳定,需要进一步强化。
2.2 不同训练阶段的神经元响应对比
对图1所示一个完整训练试次过程中目标观察阶段神经响应数据的变化过程进行重点分析。为了作为参照,分别往前和往后多选取1 s的数据组成共5 s,后续的分析均是针对该5 s的数据进行的(后续统一称为特定时间窗内)。
2.2.1 Spike信号响应特性分析
分析了特定时间主要提取特定时间窗内神经元的Spike发放率特征,分析鸽子外纹体神经元在视觉认知过程中响应的变化规律,以一个经典神经元(鸽A1的2通道神经元)为例,绘制了其Spike平均发放率随时间的变化曲线,如图4所示。
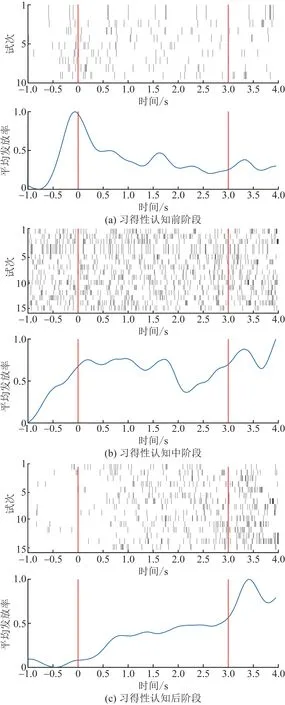
图4 不同训练阶段单神经元活动和归一化Spike发放率Fig.4 Single neuron activity and normalized Spike firing rate at different training stages
由图4可以看出,在习得性认知前阶段,信鸽没有做过或者做过很短时间的训练任务,并未对训练任务中呈现的目标产生认知,即信鸽不能认识到目标与奖励的关联,任务成功率具有很大偶然性,由图4(a)不难看出,在屏幕呈现认知目标期间,神经元对目标刺激产生较少的动作电位。
认知中阶段,信鸽可快速学习认知目标和适应训练模式,由图4(b)可以看出,神经元目标呈现期间发放率高于呈现前,并且在此期间持续产生动作电位,此阶段神经元响应信息主要集中在目标呈现期间。
认知后阶段,信鸽已完全适应训练模式,并能快速识别目标,开门后可快速选择奖励区获得奖励,由图4(c)可以看出,在呈现认知目标期间神经元Spike信号发放率逐渐增加,并在开门后1 s内达到峰值,此阶段神经元响应主要集中开门前后的时段,即执行行为前决策发生的时段。
2.2.2 不同试次响应间的同步性分析
进一步计算了不同试次神经元Spike信号发放率之间的相关系数,用于衡量神经元响应的可靠性。以其中8 d数据为例,每一天不同试次的同步性统计结果(均采用均值±标准差的形式)如图5所示。

红线为对平均相关系数作线性拟合后的曲线(p<0.05,95%置信区间)图5 不同训练阶段神经元发放率特征不同试次之间的相关系数变化曲线Fig.5 Correlation coefficient change curve of neuron Spike firing rate between different trials at different training stages
由图5可以看出,随着训练程度的提高,不同试次神经元发放率间的相关系数逐渐增大,结果表明信鸽习得性目标认知建立后,不同试次得到的响应特征趋于稳定,而在习得性目标认知前阶段不同试次间相关系数较小,说明训练前的响应包含了更多干扰信息,具有一定的随机性。
2.2.3 LFP信号响应特性分析
分析了待定时间窗内神经元LFP信号伽马频带能量特征。首先使用MTM方法估计了目标呈现前1 s和呈现期间3 s内LFP的信号频谱,得到一个具有明显差异的子频带。以其中一个神经元(A1鸽的2通道神经元)为例,对比结果如图6所示。

实线为目标呈现前信号的能量谱;虚线为目标呈现期间的能量谱图6 伽马频带的LFP信号频谱图Fig.6 Spectrum diagram of LFP signal in gamma band
由图6可以看出,能量在伽马频带中53~67 Hz频率段有明显增强。因此,主要针对LFP信号伽马子频带(55~70 Hz)进行时频分析。同样采用MTM方法估计每个试次特定时间窗内(宽度1 s,步进长度0.5 s)LFP信号伽马子频带的时频谱图,如图7(每个子图的上框图)所示。进一步地将所有频带的能量进行叠加,绘制能量随时间变化的趋向,如图7每个子图的下框图所示。

图7 不同训练阶段单神经元的LFP信号伽马子频带时频谱图Fig.7 Time-frequency spectrum diagram of LFP signals in the gamma subband of a classical neurons at different training stages
通过对比习得性认知前、认知中和认知后三个阶段伽马子频带能量的变化,发现其与Spike信号发放率特征具有相似的响应变化规律,在认知前阶段, 1~3 s内信号能量有明显增强,即响应信息主要集中在目标呈现期间;在认知后阶段,2.5~3.5 s内能量增强,即响应信息主要集中在执行行为前的决策期间。
2.2.4 两两神经元间的互信息量分析
进一步对比了习得性目标认知前后神经元间携带的信息量的变化。4只鸽子54对神经元互信息量的统计结果散点图,如图8所示。

每一个点表示一对神经元;红色为基于局部场电位的结果;黑色为基于Spike发放率的结果图8 神经元与神经元之间互信息量的对比关系Fig.8 Comparison of mutual information between two neurons
统计结果表明,认知后阶段信鸽外纹体两两神经元之间基于Spike信号发放率的互信息量非常显著地低于认知前阶段(p< 0.01,Wilcoxon秩和检验),基于LFP信号的互信息量稍显著地低于认知前阶段(p< 0.05,Wilcoxon秩和检验)。说明对于习得性目标信鸽神经元集群间响应信息中冗余信息减少,其编码信息能力得到加强。此外,基于Spike发放率的互信息量普遍低于LFP,可能是由于LFP中包含更多反映背景整合成分的冗余信息而造成的。
3 讨论
采用信鸽作为动物训练模型,对信鸽进行目标认知训练,同步采集信鸽外纹体区神经元集群响应信号,提取了Spike发放率特征,采用MTM方法估计并计算了LFP信号特定频带能量密度,进一步从不同试次响应的同步性和神经元间的互信息量角度,对比信鸽在不同的目标认知阶段其外纹体神经元响应特征的变化,定量分析了信鸽神经元对习得性目标的响应规律。本文研究中得到了与Johnston等[12-13]相似的研究结果,Spike发放序列在习得目标前后发生了改变,信鸽在习得性认知前阶段,其响应信息主要集中在目标呈现期间;在认知后阶段,响应信息主要集中决策期间,并且不同试次之间神经元响应趋于一致。随着信鸽认知程度的提高,对于习得性认知目标,其神经元可以高效地执行视觉信息加工处理,编码效率显著提高。
4 结论
已有研究揭示了鸽子视觉神经元偏侧化的三个属性:鸽子用右眼-大脑左半球学习视觉辨别更快;左半球的神经元具有更高的辨别能力;双眼条件下的视觉识别多为左半球控制[11],因此,本文研究中只考虑了信鸽左脑外纹体神经元的响应特性,并未涉及信鸽视觉神经元偏侧化这一方面,所以在认知前阶段,信鸽对于偏左眼方位呈现认知目标时,其获得奖励的决策受到偏侧化影响。此外,对于习得性目标,信鸽在决策期间,其外纹体神经元有着比目标呈现期间更为明显的活动,表现在神经元在此期间Spike信号发放率达到峰值,特定频率段内LFP信号能量明显增强,对于不同训练阶段奖励信息对信鸽外纹体神经元活动的影响也并未过多涉及。存在的不足之处将在以后的研究中进行改进和完善。
对信鸽目标认知过程中其外纹体神经元的响应特性做出了定量分析,为进一步研究鸟类高级视觉皮层对目标认知过程的信息处理机制提供了重要参考。
