树种混交度
——一个基于树种谱系的林分混交度新概念
2021-10-08冯思思李建军
张 科,冯思思,黄 浪,李建军
(1.浙江省林业信息宣传服务中心,杭州 310020;2.中南林业科技大学 计算机与信息工程学院,长沙 410000)
对林分空间结构分析和模拟优化是森林可持续经营一个重要研究内容[1-2]。量化林分空间结构的指标有很多,如混交度、角尺度、大小比数、密集度等,其中混交度是用来描述树种混交程度(即树种空间隔离程度)的一个常用参数。
传统混交度的概念定义了空间结构单元内对象树与其最近邻木为不同种时的个体所占比例[3-7],其后,有多位学者在此基础上展开了进一步的分析、研究,对混交度的定义进行了修正和完善。2001年,惠刚盈等[8-9]通过比对分析,认为混交度最适合的空间结构单元为“1+4”,即由对象树与其最近的4株邻木所组成的结构单元,并提出了林分平均混交度的概念,得到了广泛的认可;2004年,汤孟平等[1]针对传统混交度仅考虑对象树与其最近邻木的种类异同,而忽略了最近邻木之间的种类异同的缺陷,提出树种多样性混交度的概念;但是惠刚盈等[10]认为树种多样性混交度混淆了4株相邻木中有3个相同种和4株相邻木有2个相同种的混交度,并提出了修正的树种混交度概念;汤孟平等[11]研究发现修正的树种混交度与树种多样性混交度存在相同的缺陷,并结合Simpson多样性指数提出了全混交度的概念,能较好地描述树种多样性和树种空间隔离关系;2014年,张结存等[12]在全混交度的基础上提出了改进型混交度,解决了全混交度不能够体现出完全混交的缺陷以及传统混交度和树种多样性混交度等忽略的树种株数对混交度的影响。
通过分析上述各种混交度的定义,不难发现,在描述对象木与最近邻木为其他种的概率时,只有两种结果:相同种或不同种,并没有考虑物种亲缘关系的影响,这显然不能完全说明树种的混交程度。基于此,本文开展了融合树种谱系关系的混交度研究。
1 研究区概况
本文实测数据涉及湖南省大围山自然保护区、龙虎山林场、青石冈林场、芦头林场和乌云界国家级自然保护区5个研究区域,在每个研究区域各设置了5块20m×20m的区域典型森林类型固定样地,对胸径大于5cm的活立木进行定位、编号及每木检尺,检测内容包括树种、胸径、树高、冠幅等,同时以样地的左下角(西南角)为原点构建坐标系,生成每株林木的水平位置。
研究区森林类型主要为亚热带常绿阔叶林和中亚热带常绿阔叶林,主要树种有枫香(LiquidambarformosanaHance)、木荷(SchimasuperbaGardn.et Champ)、小叶青冈(Cyclobalanopsismyrsinifolia(Blume) Oersted)、青冈(QuercusglaucaThunb)、甜槠(Castanopsiseyrei(Champ.) Tutch)、青冈栎(Cyclobalanopsisglauca(Thunb.) Oerst)、杉木(Cunninghamialanceolata(Lamb.) Hook)、苦槠(Castanopsissclerophylla(Lindl.) Schott)、栲树(CastanopsisfargesiiFranch)、多脉青冈(CyclobalanopsismultinervisW.C.Cheng et T.Hong)、鹅掌楸(Liriodendronchinense(Hemsl.) Sargent)、冬青(IlexpurpureaHassk)、山茶(CamelliajaponicaL.)、长叶石栎(Lithocarpusharlandii(Hance ex Walp.) Rehd)、马尾松(PinusmassonianaLamb)、石栎(Lithocarpusglaber(Thunb.) Nakai)、白栎(QuercusfabriHance)、香樟(Cinnamomumcamphora(Linn.) Presl)等;25块样地中胸径大于 5 cm的林木共计1 257株,分属于22目31科48属62个树种(表1)。
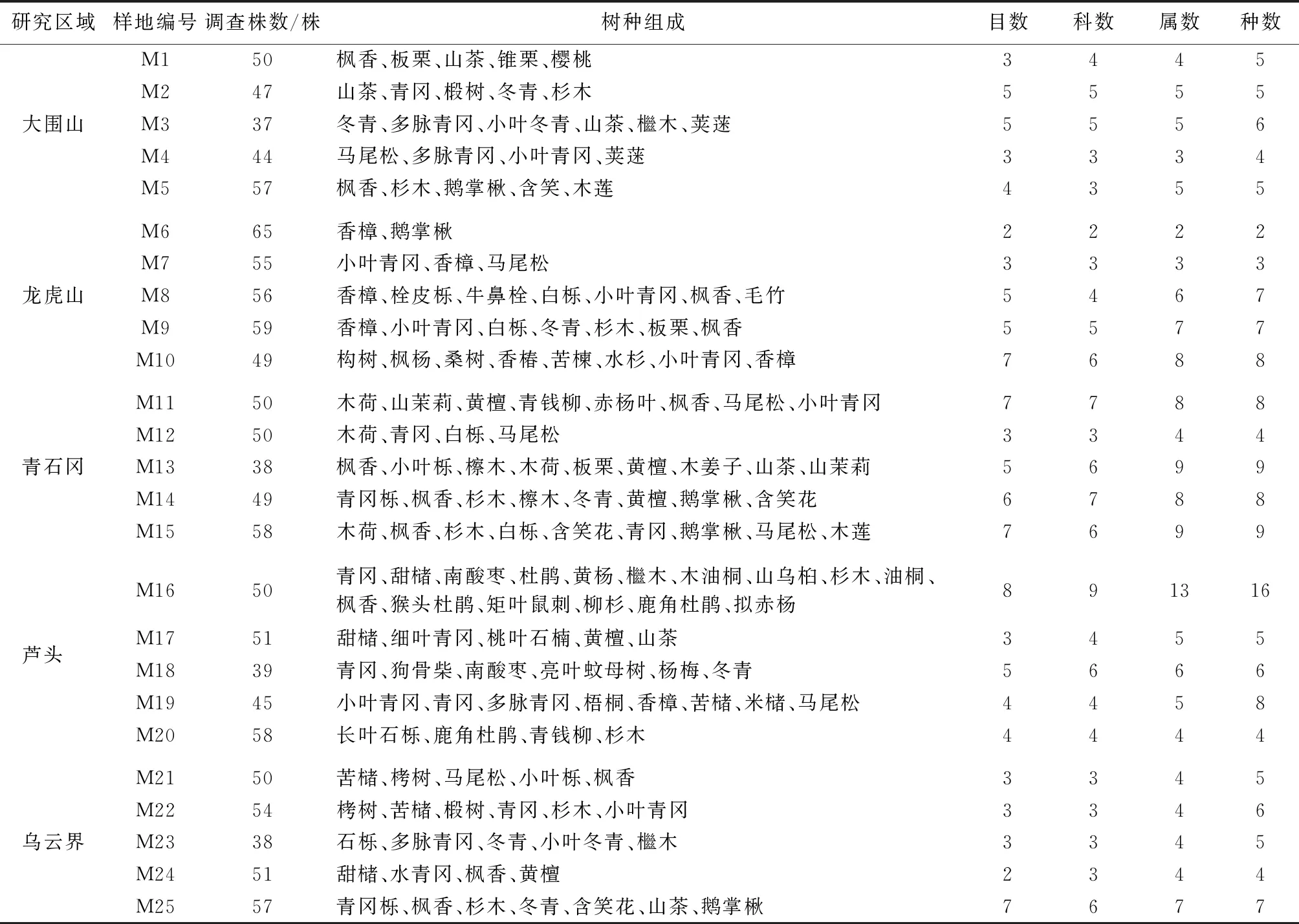
表1 样地概况
2 研究方法
2.1 传统混交度计算
混交度的计算通常是以林分内每一株林木为对象树,构建包含4株最近林木的空间结构单元,计算出每一株林木的混交度后,再取均值得到林分的平均混交度。
单木混交度计算表达式:
(1)
林分平均混交度计算表达式:
(2)
式(1)中:n为相邻木株数,一般取值为4;当第i株林木与第j株相邻木为同种时,vij=0;否则,vij=1。
式(2)中:N为林分内林木总株数。
2.2 基于树种谱系的混交度计算
生物学上将生物种群按“域、界、门、纲、目、科、属、种”予以分类,从上层的“域”到最低层的“种”,愈往下层物种之间的亲缘关系越近。根据负密度理论作用机制可知,近缘物种之间由于存在类似的生长环境,相互之间存在竞争会产生负效应[13-15]。因此,引入树种谱系关系能进一步细化林木之间的混交程度。本文考虑到按照完整的生物谱系分类结构过于繁琐,暂时仅从目、科、属、种分类进行研究,并以树种的亲缘谱系关系为依据进行逻辑赋值,用以下系数来表示,其取值结构如图1所示。基于树种谱系的混交度(简称树种混交度)计算表达式如下:

图1 vp取值结构
(3)
(4)
式(3)中:n为相邻木株数,一般取值为4;vpij取值如表2所示。式(4)中:N为林分内林木总株数。

表2 vp取值及说明
2.3 林分空间结构均质性模型
林分空间结构均质性评价模型是由李建军等[16-17]根据乘除法基本思想所提出的,该指标包含了林分空间结构在混交、竞争和空间分布格局3方面的内容,选取了混交度、大小比数、林层指数、竞争指数、空间密度指数、角尺度及开阔比数7个指标组成,其模型结构如图2所示。

图2 林分空间结构均质性模型结构
3 结果分析
3.1 传统混交度与树种混交度理论情况对比
依据公式(1)和(3)中vij和vpij的取值定义,当n=4时,传统混交度Mi的计算值有5种可能情况;而树种混交度TSMi的计算值有70种可能情况,远大于传统混交度的理论情况,这说明树种混交度在传统混交度的基础上可以更精确地反映结构单元中树种隔离程度。理论分析情况如表3和表4所示。

表3 Mi值理论情况

表4 TSMi值理论情况
3.2 传统混交度与树种混交度实测数据计算

表与计算结果及相关性分析

3.3 树种混交度在林分结构优化中的应用
3.3.1优化目标选择
遵循近自然经营理念,顺应自然规律,使林地的经济效益与生态效益协调平衡,林地的择伐数通常不超过该林地总林木数的15%。本研究中采用林分空间结构均质性模型计算25块样地中各单木的均质性指数Li,并按10%的择伐比例,选取各样地内均质性指数较低的林木作为择伐优化的目标树,以M1样地为例(表6)。林分空间结构均质性模型计算如式(5)[16]所示:
Li=
(5)
式中:TSMi为树种混交度;σTSMi为混交度标准差;OPi为开阔比数;σOPi为开阔比数标准差;Si为林层指数,σSi为林层指数标准差;Ui为大小比数,σUi为大小比数标准差;CIi为竞争指数,σCIi为竞争指数标准差;Di为空间密度指数,σDi为空间密度指数标准差;Wi为角尺度,σWi为角尺度标准差。
如表6所示,计算样地当前的均质性指数后,选取部分指数值较低的林木作为模拟择伐的对象,并计算模拟择伐后的林分整体均质性指数,以验证是否有利于林分状态的优化提升。

表6 样地M1择伐优化目标
3.3.2优化结果评价
(6)
式中:N表示林分中的林木株数。

表7 均质性指数评价标准
观察表8可知,模拟择伐优化后,25个样地的林分空间结构均质性指数均得到了提升,其中更有12个样地的评价等级获得了提升,说明优化的效果比较好,树种混交度在实际应用中是可行的。

表8 优化前后林分空间结构均质性指数对比

(续表)
4 结论与讨论
本研究考虑负密度效应,在传统混交度的基础上,结合树种谱系关系,构建了一个新的混交度指数——树种混交度指数;以湖南省大围山自然保护区、龙虎山林场、青石岗林场、芦头林场和乌云界自然保护区5个研究区域内的25块样地的实测林分数据,对传统混交度指数与树种混交度指数进行了对比分析,并利用林分空间结构均质性模型对各样地进行了模拟择伐优化,得出了以下结论:
1) 在对象树与其n株最近相邻木组成的林分空间结构单元中,当林木之间存在亲缘关系时,对象树的树种隔离情况受林木间亲缘谱系关系远近而影响不同,且TSMi 2) 借鉴林分空间结构均质性模型将树种混交度指数替换模型中原有混交度指数,对25块样地进行了模拟择伐优化,优化后有18块样地的林分空间结构均质性指数都得到了不同程度的提升,表明树种混交度指数用于林分空间结构评价与优化是可行的。 本文分析林木生物特性中亲缘谱系关系是从遗传学和分类学思想出发,按逻辑关系判断,群落生态学另有研究计算物种间谱系距离的方法,可作为研究树种隔离程度的新思路;同时,还可以考虑将生物亲缘谱系关系与其他的林分空间结构指数相结合,进一步完善相关指标的内涵。
