ZmbZIP76 通过减轻活性氧损伤和渗透胁迫增强植物对盐碱胁迫的耐受性
2021-10-05贺琳张苗苗吴紫璇孙立强杨克军
贺琳,张苗苗,吴紫璇,孙立强,杨克军
(黑龙江八一农垦大学,大庆 163319)
干旱和盐碱等非生物逆境是限制植物生长和减少作物产量的主要环境因素[1-2]。植物为了生存,在长期的进化过程中形成了复杂而高效的应答机制,从分子、细胞、生理和生化水平做出适应性调整,以抵御不利环境[2]。在植物应答非生物逆境过程中,转录因子起着关键的调控作用。碱性亮氨酸拉链(basic leucine zipper,bZIP)转录因子家族是植物中规模最大、种类最丰富的转录因子之一,其通常又被称为AREB(ABA-reactive element binding factor)或ABF(ABRE-binding factors)[3]。
bZIPs 家族基因在植物抵御非生物胁迫反应中发挥重要作用,大部分bZIP 基因如ABF2/AREB1,ABF4/AREB2 和ABF3 可以被ABA、干旱和盐诱导表达[4-5]。一些bZIP 基因的过表达可以提高植物对非生物胁迫的耐受性。例如,ABF3 和ABF4 蛋白通过上调ABA 和逆境相关基因表达进而提高转基因拟南芥对干旱的耐受性[6]。拟南芥bZIP 基因AtTGA4 通过改善硝酸盐的运输和同化,提高转基因植物的抗旱性[7]。AtABF3 在苜蓿(Medicago sativa)中过表达降低了叶片的蒸腾速率和活性氧的积累,增强了叶片对干旱、盐和氧化胁迫的耐受性[8]。OsbZIP23 在水稻中的过表达显著提高了水稻的抗旱性、耐盐性以及对ABA 的敏感性[9]。OsbZIP72 在水稻中的过表达可以提高ABA 响应基因的表达水平,转基因植物对ABA的敏感性提高,抗旱性增强[10]。干旱和ABA 处理诱导了棉花GhABF2 的表达,但高盐抑制了GhABF2 的表达,干旱和盐处理后过表达GhABF2 的棉花脯氨酸含量、超氧化物歧化酶活性和过氧化氢酶活性均显著高于野生型[11]。玉米ABP9 基因能够被ABA、H2O2、干旱和盐诱导表达,过表达ABP9 基因的拟南芥对干旱、盐、冷和氧化胁迫等多种非生物逆境表现出较强的耐受性[12]。虽然目前关于bZIP 转录因子在盐、旱和冷逆境下的功能研究较多,但是关于bZIP转录因子在植物抵御盐碱复合胁迫过程中的功能的报道相对较少。
Jakoby 等[13]根据碱性结构域及保守结构域,将拟南芥的75 个bZIP 类转录因子基因家族成员分为A、B、C、D、E、F、G、H、I 和S 类10 个亚家族。其中,A 亚族bZIPs 在种子和植物组织对ABA 和逆境胁迫的响应信号网络中发挥重要作用。例如,Ying 等[14]发现AbZIP 转录因子ZmbZIP72 在拟南芥中过表达可增强植物的抗旱性和耐盐性,提高种子对ABA 和渗透胁迫的敏感性。大豆bZIP 基因GmbZIP44,GmbZIP62和GmbZIP78 分别属于S、C 和G 亚家族,三者在拟南芥中过表达,均能提高转基因拟南芥对盐和低温胁迫的耐受性[15]。B 亚家族的两个成员AtbZIP17(At2g40950)和AtbZIP28(At3g10800)参与调控盐胁迫反应和内质网胁迫反应[16-17]。F 亚家族是个小亚家族,其成员(F-bZIPs)主要参与调控植物的缺锌胁迫反应[18-19]。拟南芥仅含有3 个F-bZIPs 基因(At4g35040,At2g16770,At3g51960),其中At3g51960(AtbZIP24)在盐胁迫网络中发挥负面调节作用[20]。有报道指出玉米中有4 条F-bZIPs 基因(GRMZM-2G000171,GRMZM2G055413,GRMZM2G175870,G -RMZM2G033230)[21],但是迄今功能均未见报道。鉴于此,研究首先采用实时定量PCR 研究F-ZmbZIPs 的表达模式,发现ZmbZIP76(GRMZM2G055413)可以同时被ABA 和NaHCO3胁迫诱导表达。进而对转基因材料进行深入分析,NaHCO3处理后,与野生型株系相比,ZmbZIP76 转基因拟南芥的根长显著增加,脯氨酸水平和活性氧清除能力显著提高,相反,ZmbZIP76 转基因拟南芥体内的过氧化氢含量和丙二醛(MDA)含量显著降低。研究结果表明ZmbZIP76 转录因子通过调控转基因植物的多种生理过程来增强其对盐碱胁迫的耐受性。
1 材料和方法
1.1 实验材料
野生型拟南芥(哥伦比亚型)为黑龙江八一农垦大学作物逆境功能组学实验室保存。为了获得过表达ZmbZIP76 的拟南芥株系,构建过表达载体35S:ZmbZIP76,转入农杆菌。挑取活化的EHA105(35S:ZmbZIP76)单菌落培养至OD600 为0.6~0.8,离心沉淀菌体。配制侵染液:5%Sugar+100 μM AS+100 μL·L-1Triton X-100+0.02% Silwet L-77+2 ng·L-16-BA+沉淀后的菌体,补充去离子水使农杆菌的OD600 为0.8。侵染拟南芥花序,收集T0 代种子在含有50 mg·L-1卡那霉素的1/2MS 培养基上筛选,获得T1 代苗,对其进行DNA 水平检测,检测成功的株系培养至T3代用于功能研究。
1.2 基因表达分析
为了分析ZmbZIP76 基因在盐碱胁迫下的表达模式,用100 mM NaHCO3的溶液处理二叶时期的合344 玉米自交系土壤苗0,12,24 和72 h。不同处理条件下的幼苗根和叶分开取样,液氮速冻,用TRIzol 法提取RNA,利用逆转录酶(FastQuant RT kit,TIANGEN,China)反转录获得cDNA 模板。以ZmActin1(GRMZM2G126010)为内参,所用引物序列见表1。反应体系为10 μL:SYBR Premix Ex TaqTmⅡ(TaKaRa公司)5 μL,上下游引物(10 μmol·L-1)各1 μL,cDNA模板1 μL,无菌双蒸馏水2 μL。反应程序为:95 ℃3 min;95 ℃30 s,58 ℃30 s,72 ℃30 s,40 个循环。每个处理3 次重复。采用2-ΔΔCt方法计算各基因表达量,利用SPSS18.0 进行差异显著性分析。

表1 荧光定量PCR 引物序列Table 1 Primers of fluorescent quantitative PCR
1.3 生长表型分析
利用10% NaClO对不同株系(WT、OE3 和OE16)的拟南芥种子进行消毒。将消毒后的种子均匀播种于1/2 MS 培养基上,在22±2 ℃,光照强度100 μmol·m-2·s-1,光照周期为16 h 光照/8 h 黑暗的人工气候室中培养,1 周后将长势相同的幼苗移栽到含有NaHCO3(0、6 mM)的1/2 MS 培养基表面进行竖直培养,胁迫1 周后对相同处理条件下OE 株系与WT 株系之间根长的差异进行统计分析(P<0.05,one-way ANOVA)。每个株系每个处理至少50 株苗,实验进行3 次独立的生物学重复。
1.4 生理指标的测定
选取1 周苗龄的野生型株系(WT)、过表达株系(OE3 和OE16)分别置于含有0、6 mM NaHCO3的1/2 MS 培养基上培养1 周,分别测定不同株系不同处理后的超氧化物歧化酶活性(SOD)、过氧化物酶活性(POD)、H2O2含量、脯氨酸含量和丙二醛(MDA)含量。SOD 和POD 活性分别采用氮蓝四唑(NBT)光化还原法和愈创木酚法测定[22];H2O2含量测定参考Ve likova 等[23];脯氨酸含量采用磺基水杨酸提取,茚三酮比色法[24];MDA 含量采用硫代巴比妥酸比色法(TBA)测定[25]。实验进行3 次独立的生物学重复,利用SPSS 软件对相同处理条件下OE 株系与WT 株系之间生理指标的差异进行统计分析(P<0.01,one-way ANOVA)。
2 结果与分析
2.1 玉米ZmbZIP76 基因在ABA 和NaHCO3 胁迫下的表达分析
ABA 和NaHCO3处理的各个时间点ZmbZIP76在根中均显著上调表达,NaHCO3处理12 h 后表达量最高,是对照的17.7 倍,随后表达量开始降低,ABA 处理24 h 后表达量最高,为对照的7.7 倍,随后表达量有所降低(图1)。ABA 和NaHCO3处理下ZmbZIP76 在叶中的表达模式极为相似,都是在胁迫处理的24 h 开始出现显著增加的表达量,在处理的72 h 表达量达到最高峰,ABA 和NaHCO3处理72 h时ZmbZIP76 的表达量分别是对照的5.9 倍和8.6 倍(图1)。
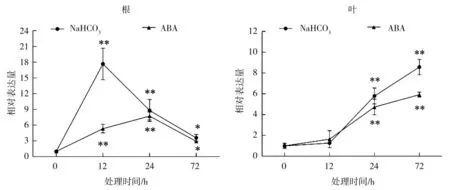
图1 NaHCO3 和ABA 处理下ZmbZIP76 基因在玉米叶和根中的表达Fig.1 The expression of ZmbZIP76 gene in leaves and roots of maize under NaHCO3 and ABA treatments
2.2 ZmbZIP76 基因过表达对拟南芥根长的影响
为了比较NaHCO3胁迫下ZmbZIP76 过表达株系与WT 株系根长的差异,将1 周大小长势一致的不同株系的幼苗分别转移到含有0、6 mM NaHCO3的1/2 MS 培养基上竖直培养1 周,测量根长并拍照。结果如图2 所示,正常生长条件下,野生型株系和过表达株系根长无显著差异,但是经过6 mM NaHCO3处理后,过表达株系OE3 和OE16 的根长分别是野生型株系根长的1.4 和1.45 倍(P<0.05,one-way ANOVA test),说明过ZmbZIP76 过表达提高了拟南芥幼苗对NaHCO3胁迫的耐受性。
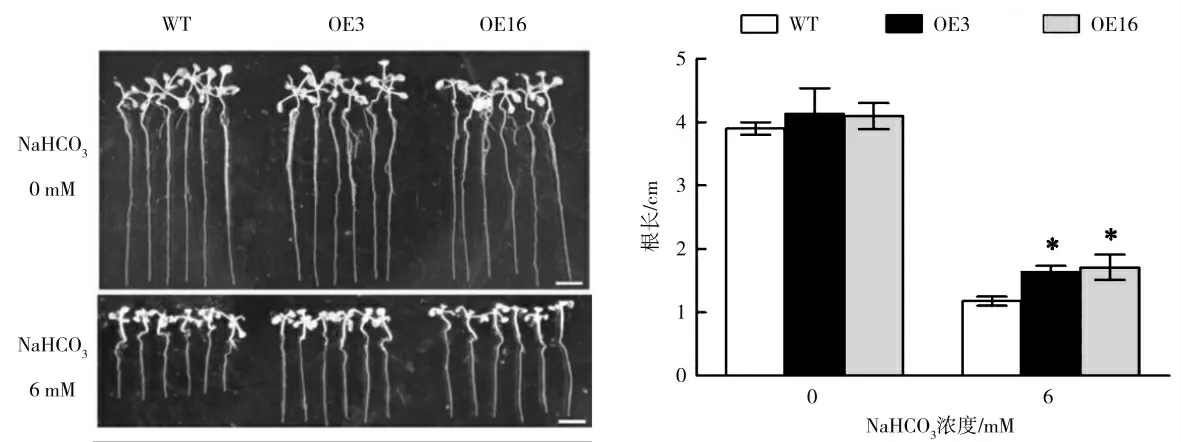
图2 NaHCO3 处理下ZmbZIP76 转基因拟南芥根长Fig.2 The root length of ZmbZIP76 transgenic Arabidopsis under NaHCO3 treatment
2.3 ZmbZIP76 基因过表达对植物抗氧化防御系统的影响
正常生长条件下,野生型株系和ZmbZIP76 过表达株系(OE3 和OE16)体内SOD 和POD 活性未见显著差异,然而,在NaHCO3胁迫下,OE3 和OE16 体内SOD 活性分别是野生型植物的1.94 倍和1.97 倍,OE3 和OE16 体内POD 活性分别是野生型植物的2.1 倍和2 倍(图3)。盐碱胁迫条件下ZmbZIP76 过表达株系中SOD 和POD 活性显著高于野生型拟南芥(P<0.01,one-way ANOVA test),表明ZmbZIP76 基因对SOD 和POD 酶活具有正向调控作用。H2O2含量测定进一步显示,NaHCO3胁迫条件下过表达株系OE3 和OE16 体内H2O2含量显著低于野生型株系(图3),进一步证明ZmbZIP76 基因通过促进植物体内活性氧的清除来提高植物对盐碱胁迫的耐受性。
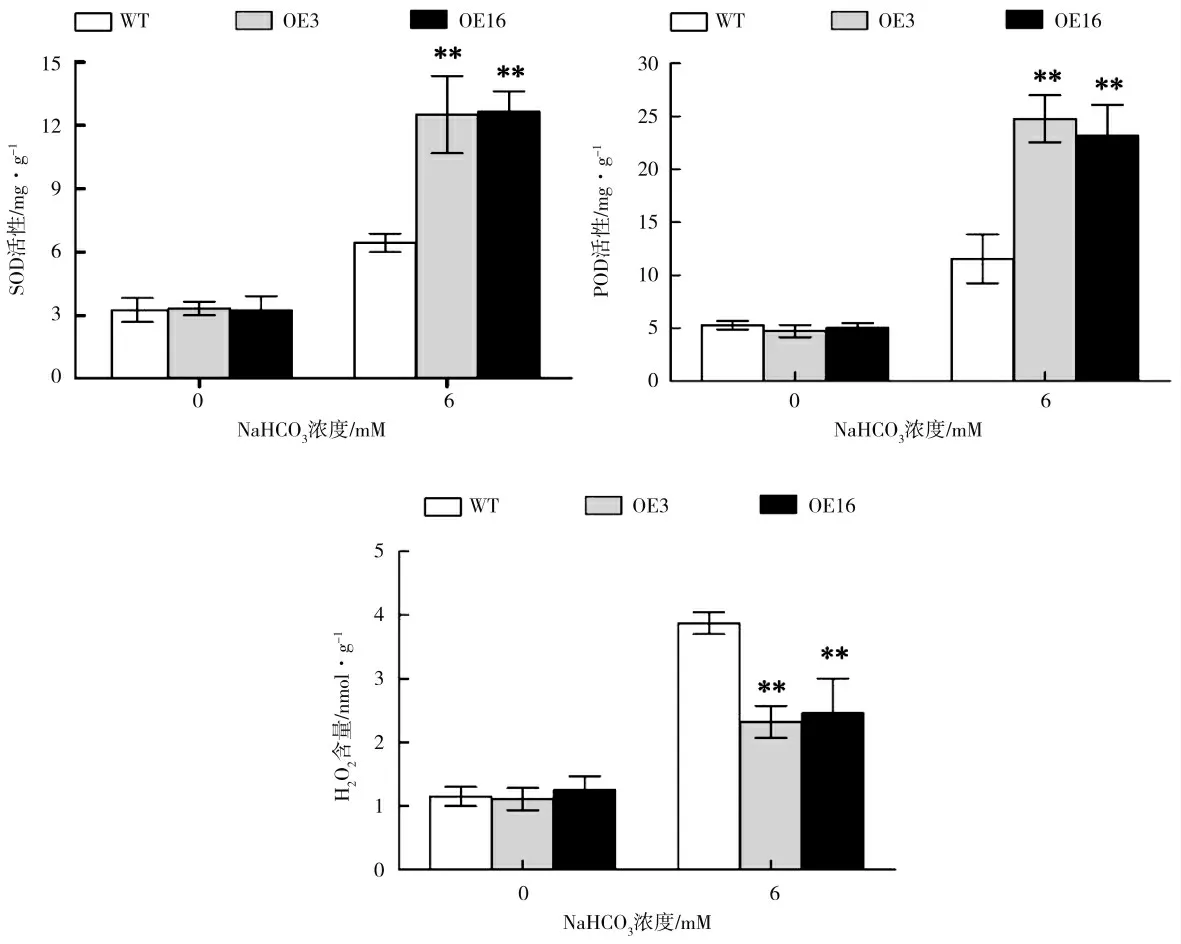
图3 NaHCO3 胁迫下ZmbZIP76 转基因拟南芥活性氧清除能力Fig.3 The reactive oxygen scavenging ability of ZmbZIP76 transgenic Arabidopsis under NaHCO3 stress
2.4 ZmbZIP76 基因过表达对植物脯氨酸含量的影响
正常生长条件下,野生型株系和ZmbZIP76 过表达株系(OE3 和OE16)体内脯氨酸含量未见显著差异,然而,在NaHCO3胁迫下,OE3 和OE16 中脯氨酸含量分别是野生型株系的1.54 倍和1.46 倍(图4)。ZmbZIP76 过表达株系中脯氨酸含量显著高于野生型(P<0.01,one-way ANOVA test),表明盐碱胁迫条件下ZmbZIP76 通过正向调控植物体内脯氨酸的合成进而提高植物耐盐碱性。

图4 NaHCO3 胁迫下ZmbZIP76 转基因拟南芥脯氨酸含量Fig.4 The proline content of ZmbZIP76 transgenic Arabidopsis under NaHCO3 stress
2.5 ZmbZIP76 基因过表达对植物MDA 含量的影响
正常生长条件下,野生型株系和ZmbZIP76 过表达株系(OE3 和OE16)体内MDA 含量未见显著差异,然而,在NaHCO3胁迫下,ZmbZIP76 过表达株系中MDA 含量显著低于野生型(P<0.01,one-way ANOVA test),OE3 和OE16 中MDA 含量分别较野生型株系下降了37.6%和39%(图5)。

图5 NaHCO3 胁迫下ZmbZIP76 转基因拟南芥MDA 含量Fig.5 The MDA content of ZmbZIP76 transgenic Arabidopsis under NaHCO3 stress
3 讨论
bZIP 转录因子在多种生物学过程以及非生物胁迫反应中发挥重要作用,然而,大部分关于bZIPs 的报道集中于拟南芥和水稻中,玉米中关于bZIP 的报道有但相对较少。从玉米中分离了一条能够同时响应 ABA 和 NaHCO3胁迫的bZIP基因GRMZM2G055413,命名为ZmbZIP76,证明了其在盐碱胁迫下的正向调控功能。
ABA 信号转导途径在非生物胁迫反应中发挥重要作用,包括转录因子在内的许多基因都可以响应ABA 信号,如DREB/CBF,MYB,bHLH,ERF,bZIP 和NAC[26]。转录因子通过与靶基因启动子中顺式元件相互作用,在响应ABA 信号中发挥重要作用[27]。其中,bZIP 是最大的转录因子基因家族之一,在调控依赖于ABA 的植物逆境反应中发挥着关键作用。目前,拟南芥中bZIP 转录因子参与的ABA 信号模式已经建立,PYR/PYL/RCARs→PP2C→SnRKs→AREBs→靶基因。其中,PYR/PYL/RCARs 是ABA 受体蛋白。PP2C(protein phosphatase 2C)是丝氨酸/苏氨酸蛋白磷酸酶,可以负调控ABA 信号转导途径及多种逆境胁迫转导途径[28]。SnRK(sucrose non-fermenting 1-related protein kinase)是广泛存在于植物中的Ser/Thr类蛋白激酶,可以与PP2Cs 发生相互作用从而导致自身失活和去磷酸化。非胁迫条件下,PP2Cs 可与SnRKs 结合,使其发生去磷酸化作用而失活[29]。逆境胁迫下,ABA 可以和受体PYR/PYL/RCAR 形成复合物,该复合物会阻断PP2C 介导的SnRKs 的失活,未失活的SnRKs 激酶可以通过磷酸化bZIP 转录因子将其激活,bZIP 转录因子进一步通过识别ABRE 元件最终激活ABA 信号相关靶基因的表达[29]。研究发现ZmbZIP76 可以同时被ABA 和NaHCO3诱导表达,值得注意的是,ZmbZIP76 在两种处理下的表达模式极为相似,其表达量在根中均是先升高再降低,在叶中都是于胁迫处理的24 h 开始显著增加,在处理的72 h 达到表达的最高峰(图1),表明ZmbZIP76参与调控的植物盐碱胁迫反应很可能依赖于ABA途径,即ZmbZIP76 转录因子可能通过调节ABA 信号来提高转基因拟南芥对盐碱胁迫的耐受性。
非生物胁迫条件下,有效的抗氧化系统是植物抵抗逆境的一个主要途径[30]。盐胁迫条件下ThbZIP1基因在烟草中过量表达,可以提高过氧化物酶(POD)和超氧化物歧化酶(SOD)的活性,增加可溶性糖和可溶性蛋白的含量,还可以促进活性氧的消除[31]。玉米中发现的bZIP 转录因子ABP9 通过调节细胞内的ROS 水平提高转基因拟南芥对多种逆境的耐受性[12]。研究发现NaHCO3胁迫下ZmbZIP76 过表达株系(OE3 和OE16)中SOD 和POD 活性显著高于野生型,H2O2含量显著低于野生型株系(图3),表明NaHCO3胁迫条件下ZmbZIP76 基因可以通过提高抗氧化防御系统中关键酶SOD 和POD 的活性,加速对活性氧的清除。类似的是,Qu 等发现微藻ChbZIP1转录因子通过对活性氧清除系统的调节来提高转基因拟南芥对盐碱胁迫的耐受性[32]。
逆境下植物体内高水平的ROS 积累会引起膜脂过氧化,从而产生大量MDA,因此逆境下植物体内ROS 和MDA 含量通常具有正向关性,MDA 含量既可以直接用于衡量膜系统受损程度,又可以间接用于反应植物体内ROS 的含量。研究发现NaHCO3胁迫条件下ZmbZIP76 过表达株系中MDA 含量和ROS含量均显著低于野生型(图3 和图5),表明ZmbZIP76 的过表达可以通过加快ROS 的清除,减轻膜系统的损伤来提高转基因拟南芥对NaHCO3胁迫的耐受性。与此一致的是,Bi 等报道TabZIP15 的过表达使小麦体内H2O2含量和MDA 含量显著降低,转基因植物表现出提高的耐盐性[33]。Wu 等发现野生大豆GsbZIP67 基因通过降低MDA 含量和提高POD活性从而来增强苜蓿对盐碱胁迫的耐受性[34]。
脯氨酸在植物抵御非生物胁迫反应中发挥重要作用,在遭受逆境时,植物体内的脯氨酸浓度会增加,以此调节细胞内外渗透压防止细胞受损。逆境条件下一些bZIP 转录因子可以同时调控抗氧化防御系统和脯氨酸的合成。例如,甘薯bZIP 转录因子IbbZIP1 的过表达在干旱和盐胁迫下不但提高了植物体内SOD 活性,减少了H2O2含量,还增加了脯氨酸含量,最终增强了IbbZIP1 转基因拟南芥在干旱和盐胁迫下的耐受性[35]。相类似的是,研究发现盐碱胁迫条件下ZmbZIP76 过表达株系中脯氨酸含量显著高于野生型,SOD 和POD 活性也显著高于野生型,H2O2含量则显著低于野生型株系(图3 和图4),表明玉米bZIP 转录因子同样可以通过正向调控抗氧化防御系统和脯氨酸合成来提高植物对盐碱胁迫的耐受性。由此可见,逆境相关bZIP 转录因子参与调控的植物抗逆生理反应在物种间具有相似性。
4 结论
ZmbZIP76 既可以受ABA 诱导表达,又可以受NaHCO3诱导表达,且在两种处理下表达模式相似。盐碱胁迫下,与野生型株系相比,ZmbZIP76 转基因拟南芥的根长显著增加,脯氨酸含量、SOD 和POD活性显著提高,过氧化氢含量和MDA 含量显著降低。研究证实了ZmbZIP76 转录因子是盐碱胁迫下的正向调节因子,研究结果将为进一步利用转基因手段培育玉米抗/耐品种提供理论依据和基因准备。
