番茄OFP基因家族生物信息学分析
2021-09-28程欣
程欣

摘要:为了解番茄OFP基因家族的功能特征,通过生物信息学手段对番茄OFP基因家族成员进行全面的生物信息学分析,包括染色体定位、基因结构、蛋白质理化性质、二级结构、三维构象、启动子分析、亲缘关系及组织特异性分析。结果表明,番茄中的31个OFP基因在12条染色体上呈不均匀分布;除SlOFP6、SlOFP7、SlOFP12、SlOFP13基因以外,其余27个OFP基因均不含有内含子结构。蛋白质理化性质分析结果表明,大部分OFP蛋白为不稳定性蛋白,并且54.8%的OFP蛋白分布于细胞核中。蛋白质二级结构和三维构象显示,番茄OFP蛋白以α-螺旋和无规则卷曲为主要组成成分,表明番茄OFP蛋白之间存在保守性。启动子顺式作用元件分析结果表明,番茄OFP基因启动子上主要存在3类元件:光响应、激素响应应答以及逆境胁迫响应元件。而组织特异性分析结果表明,番茄OFP基因在各组织中的表达量差异较大,预示着番茄OFP基因生物学功能的多样性。分析结果将为后续研究番茄OFP基因家族成员的生物学功能提供参考。
关键词:番茄(Solanum lycopersicum);OFP基因家族;生物信息学分析;蛋白质理化性质;蛋白结构
中图分类号:S641.201;Q78 文献标志码: A
文章编号:1002-1302(2021)18-0039-09
收稿日期:2021-01-29
基金项目:重庆市科委自然科学基金(编号:cstc2018jcyjAX0458)。
作者简介:程 欣(1996—),女,四川广安人,硕士研究生,主要从事番茄功能基因的克隆与应用研究。E-mail:chengxin599@163.com。
番茄(Solanum lycopersicum)原产于南美洲,是管状花目茄科番茄属的一种一年生或多年生草本植物,是兼具营养价值和商业价值的一种蔬菜作物,是植物分子生物研究的一种模式植物。番茄富含大量维生素C、番茄红素和矿物质等多种营养成分,具有抗癌防衰、提高人体免疫力的功效,因此受到越来越多人的青睐。番茄果实为多汁浆果,有圆球形、高圆形、扁圆形、卵圆形、方圆形、梨形等多种果形。研究表明,OVATE是第一个被鉴定为控制番茄果实形状的主要定量性状基因座(QTL)[1]。
OFP基因家族编码OFP蛋白(OVATE family proteins)。OFP蛋白是仅存在植物中的一类转录抑制因子,其C末端包含一个约70个氨基酸残基的保守结构域,被称为OVATE結构域[2-5]。虽然OFP基因广泛地分布于植物中,但对于它们的功能研究却相对比较滞后,仅在拟南芥、水稻、辣椒、香蕉和番茄中开展有限的几个方面的功能研究,包括果实形状[1]、果实成熟与品质[6]、胚珠发育[7]、维管束的发育[8]、次生细胞壁的形成[9]、非生物胁迫[2]以及植物表面质积累[3]等。到目前为止,对OFP蛋白功能的研究更多的聚焦在拟南芥和水稻中,而番茄OFP蛋白的功能研究还比较少,仅对OVATE[1,10-12] 和SlOFP20[13-15] 基因的功能进行过报道,其他29个OFP蛋白的功能还未见报道。因此,本研究旨在利用生物信息学方法对番茄OFP基因家族的染色体定位、基因结构、保守基序、蛋白序列比对、系统进化关系、蛋白理化性质、蛋白质结构特征及功能、顺式作用元件的组成以及在不同组织中的差异表达等进行分析及预测,以期为进一步开展番茄OFP基因家族的功能研究提供参考。
1 材料与方法
1.1 番茄OFP基因家族成员的鉴定与染色体定位
从茄科基因组数据库(https://solgenomics.net/search/locus)下载其全部蛋白序列,再通过NCBI(https://www.ncbi.nlm.nih.gov/)在线数据库BLAST下载番茄OFP基因家族全部序列信息。从茄科基因组数据库(https://solgenomics.net/search/locus)获得OFP基因家族成员在染色体上的位置信息。
利用在线工具MG2C(http://mg2c.iask.in/mg2c_v2.1/)制作染色体定位图。
1.2 番茄OFP基因家族基因结构分析
从NCBI(https://www.ncbi.nlm.nih.gov/)在线数据库下载番茄OFP基因家族成员的蛋白质编码区(CDS)和DNA序列的FASTA格式。利用GSDS在线网站(http://gsds.cbi.pku.edu.cn/)绘制基因内含子和外显子及非编码区的分布图。
1.3 番茄OFP基因家族蛋白保守基序分析
通过MEME网站(http://meme-suite.org/)在线分析蛋白的保守基序,motif设置为10,参数设置为默认值,利用TBtools软件绘制保守基序图。
1.4 番茄OFP基因家族蛋白保守结构域序列比对和系统进化树分析
通过DNAMAN 6.0软件完成蛋白序列的氨基酸保守性分析,通过MEGA 6.0软件完成系统进化分析,随后通过iToL(https://itol.embl.de/upload.cgi)在线网站对进化树进行美化。
1.5 番茄OFP基因家族蛋白理化性质和亚细胞定位进行分析
通过ExPASy(https://web.expasy.org/protparam/)在线网站进行蛋白理化性质分析,通过GenScript(https://www.genscript.com/psort.html?src=leftbar)进行亚细胞定位分析。
1.6 番茄OFP基因家族蛋白二级结构分析
通过SOPMA在线网站(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)对蛋白序列的二级结构进行预测分析。
1.7 番茄OFP基因家族蛋白三维结构模拟
通过Swiss-Modle在线网站(https://swissmodel.expasy.org/)完成蛋白的同源建模,以获取三维结构模型。
1.8 番茄OFP基因家族基因启动子区域顺式作用元件分析
通过茄科基因组数据库(https://solgenomics.net/search/locus)下载OFP基因家族基因起始密码子上游3 000 bp区域序列作为启动子序列,然后通过PlantCare网站(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对基因启动子区域顺式作用元件进行预测分析,通过TBtools软件绘制启动子顺式作用元件图。
1.9 番茄OFP基因家族成员转录表达模式分析
通过NCBI PubMed 下载番茄的RNA-seq数据,查找搜索31个番茄OFP基因在各组织中的表达量,利用Helml软件绘制基因表达热图。
2 结果与分析
2.1 番茄OFP基因家族成员的鉴定与染色体定位
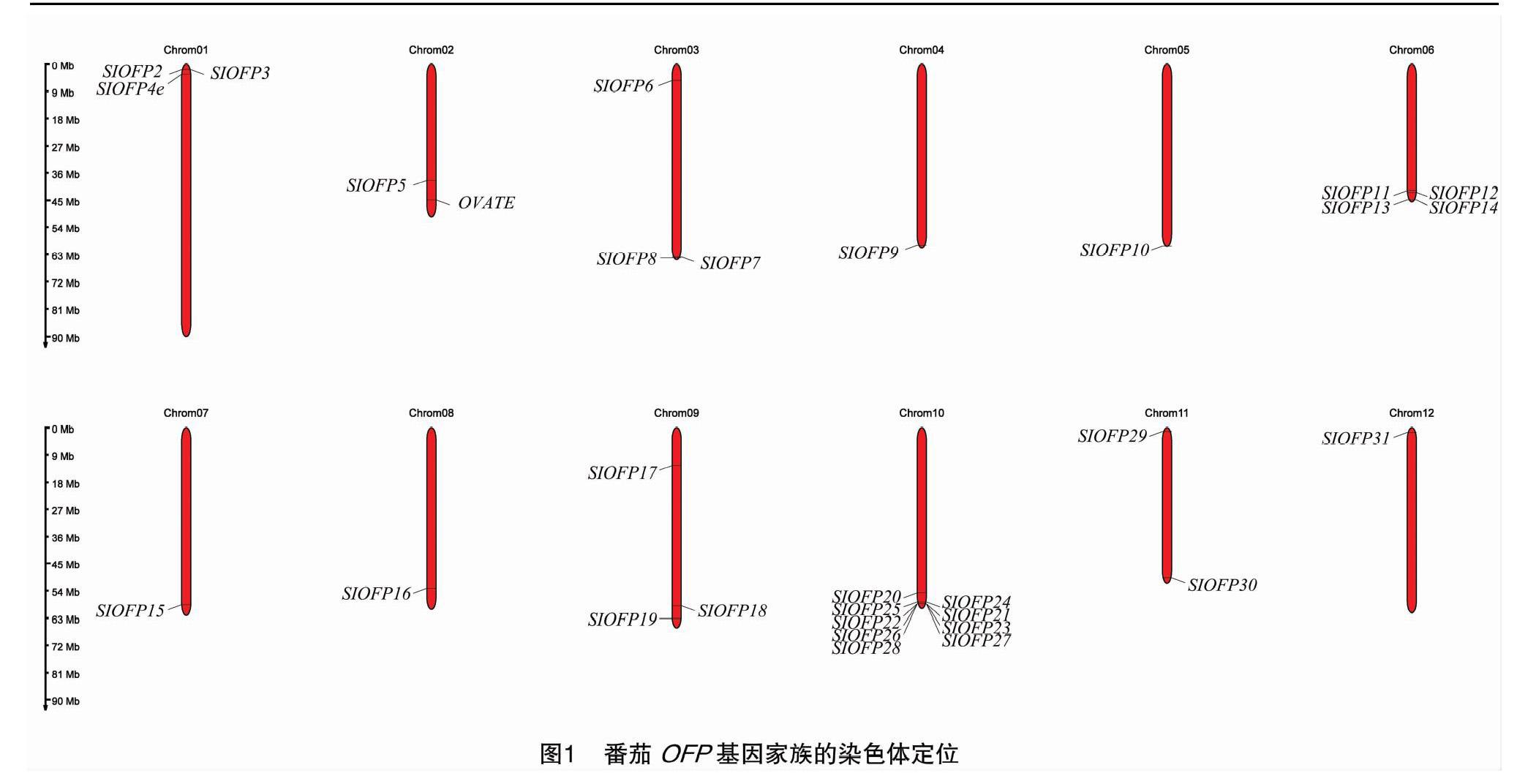
在茄科基因组数据库中检索OFP蛋白序列信息,再通过NCBI数据库和FGENESH搜索分析,最终共得到31个番茄OFP基因家族成员,并依次将它们命名为OVATE-SlOFP1~OVATE-SlOFP31。为了将这31个成员进行染色体定位分析,利用 SGN数据库确定了番茄OFP基因位置信息,结果表明OFP基因家族在番茄12条染色体上的分布并不均匀,其中10号染色体上分布的成员最多,有9个OFP基因;其次是6号染色体上分布有4个OFP基因;1、3、9号染色体上各有3个OFP基因;其余染色体上分布着 1~2个OFP基因(图1)。
2.2 番茄OFP基因家族基因结构分析
对番茄OFP基因家族的编码序列和基因序列信息进行分析,结果表明番茄OFP基因家族的编码序列外显子数量在1~3个之间,此外,除SlOFP6、SlOFP7、SlOFP12、SlOFP13以外,其他基因均不含有内含子结构(图2)。这体现出番茄OFP基因家族基因结构简单的特征。同时也说明番茄OFP基因家族呈现一定的保守性。
2.3 番茄OFP基因家族蛋白保守基序分析
对31个番茄OFP基因家族成员的蛋白序列进行保守基序预测分析,结果(图3)表明番茄OFP基因家族蛋白成员具有1~10个保守基序(Motif),只有SlOFP8蛋白不具有Motif1保守基序。另外,除SlOFP11蛋白以外其余所有OFP蛋白都含有Motif2和Motif3保守基序,且先后顺序都相同,这说明OFP基因家族在番茄中比较保守。此外,有研究表明,保守基序图中字母越高,表明该字母对应的碱基出现的频率越大,相对保守程度也越高。由图3可知,保守基序Motif2保守程度最高,Motif4、Motif6和Motif9基序保守程度也较高,对保守基序进行分析将有利于探索蛋白质的结构组成。
2.4 番茄OFP基因家族蛋白保守结构域序列比对和系统进化树分析
对番茄OFP基因家族蛋白序列进行氨基酸组成对比分析,结果表明OFP基因家族蛋白存在高度保守的氨基酸序列组成的保守结构域(OVATE结构域),这与其在进化过程中的功能高度保守密切相关(图4)。
为了进一步明确番茄OFP基因家族不同蛋白之間的亲缘进化关系,基于番茄基因组中31条OFP蛋白序列构建系统进化树,结果显示,番茄中已经被鉴定功能的OVATE和SlOFP20分别与SlOFP6、SlOFP17最同源(图4)。
2.5 番茄OFP基因家族蛋白理化性质和亚细胞定位分析
利用ExPASy在线软件对番茄OFP家族编码的蛋白质进行理化性质分析。结果表明,蛋白质的氨基酸长度在81~504 之间,其中SlOFP16蛋白氨基酸数量最多,SlOFP11蛋白氨基酸数量最少;蛋白质的相对分子质量在10.925 74 ~57.230 52 ku之间,最大的是SlOFP16蛋白,最小的是SlOFP11蛋白;等电点在4.28~10.37之间,最大的是SlOFP17蛋白,最小的是SlOFP24蛋白;脂溶指数在52.3~88.11之间,最大的是SlOFP11蛋白,最小的是SlOFP31蛋白;不稳定系数在15.72~74.16之间,最稳定的是SlOFP11蛋白,最不稳定的是SlOFP15蛋白;蛋白质的总平均疏水性数值在-1.016~-0.140之间,所有蛋白质的疏水性数值均为负值,说明番茄OFP基因家族蛋白均属于亲水性蛋白。亚细胞定位预测结果表明,SlOFP2、SlOFP10蛋白定位于内质网上,SlOFP3、SlOFP6、SlOFP18和SlOFP21蛋白定位于线粒体内,SlOFP8、SlOFP19、SlOFP22、SlOFP27、SlOFP29和SlOFP30蛋白定位于细胞质中,仅有SlOFP26蛋白定位于过氧化物酶体内,其余OFP蛋白皆定位在细胞核上(表1)。
2.6 番茄OFP基因家族蛋白二级结构分析
利用SOPMA在线软件对番茄OFP基因家族成员进行蛋白质二级结构预测,结果表明,番茄OFP蛋白质都是由α-螺旋(alpha helix)、β-转角(extended helix)、无规卷曲(random coil)和延伸链(extended strand)等4种二级结构构成(表2)。其中,SlOFP5蛋白几乎不含β-转角;在SlOFP4的蛋白二级结构中,延伸链占比>α-螺旋占比,其余OFP蛋白二级结构中都是α-螺旋占比>延伸链占比,并且所有蛋白的二级结构中,α-螺旋和无规则卷曲占据主体部分。因此可以判断,α-螺旋和无规则卷曲是番茄OFP蛋白的主要组成成分,β-转角和延伸链散布于整个蛋白序列中,起辅助修饰作用。
2.7 番茄OFP基因家族蛋白三维结构模拟
为了解析番茄OFP基因家族的三维构象,采用同源建模法,利用Swiss-Modle在线软件进行同源建模。结果表明,SlOFP4、SlOFP6、SlOFP10、SlOFP14、SlOFP16、SlOFP17、SlOFP20、SlOFP22和SlOFP24蛋白的三级结构相似;SlOFP2和SlOFP21蛋白的三级结构相似;SlOFP5和SlOFP19蛋白的三级结构也相似(图5)。说明番茄OFP基因家族蛋白之间既存在保守性又存在差异性。
2.8 番茄OFP基因家族基因启动子区域顺式作用元件分析
通过SGN数据库获取番茄OFP基因家族成员起始密码子ATG上游3 000 bp的序列,接着利用 PlantCARE 网站对其进行启动子顺式作用元件分析,结果(图6)显示,番茄OFP基因家族成员起始密码子上游存在着光响应顺式元件(G-Box、ACE、GT1-motif)、激素响应应答元件(CGTCA-motif、P-box、ABRE、TCA-element、AuxRR-core)以及逆境胁迫响应顺式元件(ARE、TC-rich repeats、LTR、MBS、DRE)。此外,胚乳表达元件(GCN4_motif)、种子特异性调控(RY-element)以及分生组织表达元件(CAT-box)在番茄OFP基因家族成员启动子上也很常见。仅有SlOFP27基因啟动子上存在干旱、低温及盐胁迫响应元件(DRE),推测该基因可能参与多个逆境胁迫响应过程;SlOFP23、SlOFP26、SlOFP27等 3个基因启动子上存在黄酮类生物合成基因调控元件(MBSI),预测这3个基因可能参与黄酮类生物合成过程。
2.9 番茄OFP基因家族成员转录表达模式分析
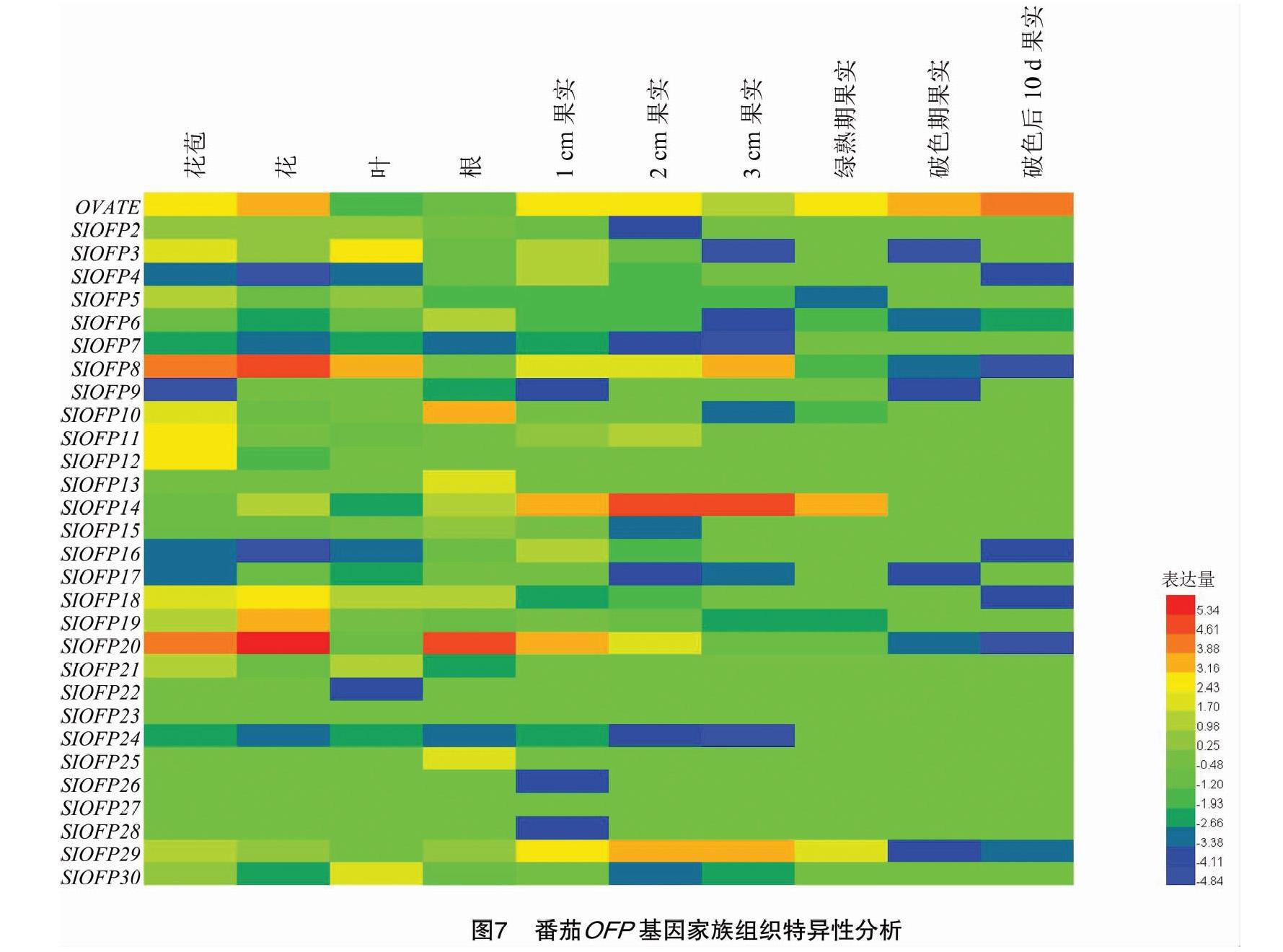
为了更好地探究番茄OFP基因家族在番茄生长发育过程中的生物学功能,本研究利用3万多差异基因表达的番茄RNA-seq数据分析了番茄OFP基因在10个番茄组织中的表达水平。结果表明,OVATE和SlOFP8 2个基因在番茄花和果实中大量表达,SlOFP14和SlOFP29 2个基因主要在青果时期大量表达,SlOFP20 基因则主要在番茄花和根中大量表达(图7)。
3 讨论与结论
番茄是全球消费量最大的蔬菜作物之一,兼具丰富的营养价值和巨大的商业价值[1]。OFP基因家族是植物特有的转录因子家族之一,通过赤霉素[8,17]、油菜素内酯[18-19]、乙烯[20]以及生长素[2]等多种激素途径,参与调控植物生长发育和非生物胁迫响应的多个方面。近年来,随着高等植物基因组测序的不断发展,研究人员在马铃薯、葡萄、拟南芥、水稻、玉米等13种陆地植物中检测出OFP基因[5]。但仅有少部分OFP基因的功能已被鉴定。因此,本研究利用生物信息学方法,对番茄中31个OFP基因进行比较全面的生物信息学分析,为探究番茄OFP基因的生物学功能提供参考价值。
本研究通过对番茄OFP蛋白的基本理化性质进行分析发现,番茄OFP蛋白均为亲水性蛋白,且大部分蛋白不稳定,这表明番茄OFP蛋白成员之间存在保守性。此外,通过对番茄OFP蛋白的二级结构进行分析发现,α-螺旋和无规则卷曲结构是主要的二级结构成分,少量延伸链和β-转角散布于蛋白序列中,起一定辅助修饰的作用。番茄OFP蛋白的三维构象则更为直观的体现出α-螺旋和无规则卷曲是番茄OFP蛋白的主要组成部分。
在组织特异性表达方面,OVATE、SlOFP8、SlOFP14、SlOFP20以及SlOFP29基因在果实中大量表达。另外,OVATE、SlOFP3、SlOFP8、SlOFP10、SlOFP11、SlOFP12、SlOFP18、SlOFP19及SlOFP20基因在花中表达量比较高,这表明大部分番茄OFP基因可能参与调控番茄花和果实的生长发育。然而这些番茄OFP基因具体的功能如何还需后续研究鉴定。
综上,本研究利用各种生物信息学分析网站和软件对番茄OFP基因家族进行了全面的生物信息学分析,包括基因结构、染色体定位、蛋白质基本理化性质、亲缘关系以及组织特异性分析等。预测分析结果为探索OFP基因在番茄各组织生长发育及非生物胁迫响应方面的生物学功能提供了理论依据和数据支持。
参考文献:
[1]Liu J,van Eck J,Cong B,et al. A new class of regulatory genes underlying the cause of pear-shaped tomato fruit[J]. Proceedings of the National Academy of Sciences of the United States of America,2002,99(20):13302-13306.
[2]Ma Y,Yang C,He Y,et al. Rice OVATE family protein 6 regulates plant development and confers resistance to drought and cold stresses[J]. Journal of Experimental Botany,2017,68(17):4885-4898.
[3]Tang Y,Zhang W,Yin Y L,et al. Expression of ovate family protein 8 affects epicuticular waxes accumulation in Arabidopsis thaliana[J]. Botanical Studies,2018,59(1):12.
[4]Wang S,Chang Y,Guo J,et al. Arabidopsis ovate family proteins,a novel transcriptional repressor family,control multiple aspects of plant growth and development[J]. PLoS One,2011,6(8):e23896.
[5]Liu D,Sun W,Yuan Y W,et al. Phylogenetic analyses provide the first insights into the evolution of OVATE family proteins in land plants[J]. Annals of Botany,2014,113(7):1219-1233.
[6]Liu J H,Zhang J,Wang J Y,et al. MuMADS1P and MaOFP1 regulate fruit quality in a tomato ovate mutant[J]. Plant Biotechnology Journal,2017,16(5):989-1001.
[7]Pagnussat G C,Yu H J,Sundaresan V. Cell-fate Switch of synergid to egg cell in Arabidopsis eostre mutant embryo sacs arises from misexpression of the BEL1-like homeodomain gene BLH1[J]. The Plant Cell,2007,19(11):3578-3592.
[8]Schmitz A J,Begcy K,Sarath G,et al. Rice ovate family protein 2 (OFP2) alters hormonal homeostasis and vasculature development[J]. Plant Science,2015,241:177-188.
[9]Li E Y,Wang S C,Liu Y Y,et al. OVATE FAMILY PROTEIN4 (OFP4) interaction with KNAT7 regulates secondary cell wall formation in Arabidopsis thaliana[J]. The Plant Journal,2011,67(2):328-341.
[10]Liu J H,Zhang J,Miao H X,et al. Elucidating the mechanisms of the tomato ovate mutation in regulating fruit quality using proteomics analysis[J]. Journal of Agricultural and Food Chemistry,2017,65(46):10048-10057.
[11]Rodríguez G R,Kim H J,Van Der Knaap E. Mapping of two suppressors of OVATE (sov) loci in tomato[J]. Heredity,2013,111(3):256-264.
[12]Wang Y,Clevenger J P,Illa-Berenguer E,et al. A comparison of sun,ovate,fs8.1 and auxin application on tomato fruit shape and gene expression[J]. Plant & Cell Physiology,2019,60(5):1067-1081.
[13]Zhou S E,Hu Z L,Li F F,et al. Overexpression of SlOFP20 affects floral organ and pollen development[J]. Horticulture Research,2019,6(1):125.
[14]Zhou S E,Cheng X,Li F F,et al. Overexpression of SlOFP20 in tomato affects plant growth,chlorophyll accumulation,and leaf senescence[J]. Frontiers in Plant Science,2019,10:1510.
[15]Wu S,Zhang B,Keyhaninejad N,et al. A common genetic mechanism underlies morphological diversity in fruits and other plant organs[J]. Nature Communications,2018,9(1):4734.
[16]霍建勇.中國番茄产业现状及安全防范[J]. 蔬菜,2016(6):1-4.
[17]Wang S C,Chang Y,Guo J J,et al. Arabidopsis Ovate Family Protein 1 is a transcriptional repressor that suppresses cell elongation[J]. The Plant Journal,2007,50(5):858-872.
[18]Yang C,Ma Y M,He Y,et al. OsOFP19 modulates plant architecture by integrating the cell division pattern and brassinosteroid signaling[J]. The Plant Journal,2018,93(3):489-501.
[19]Yang C,Shen W J,He Y,et al. OVATE family protein 8 positively mediates brassinosteroid signaling through interacting with the GSK3-like kinase in rice[J]. PLoS Genetics,2016,12(6):e1006118.
[20]Liu J H,Zhang J,Hu W,et al. Banana ovate family protein MaOFP1 and MADS-box protein MuMADS1 antagonistically regulated banana fruit ripening[J]. PLoS One,2015,10(4):e0123870.
